
Abstract
Parkinson’s disease (PD) is a neurodegenerative movement disorder due to selective loss of dopaminergic neurons of mesencephalic substantia nigra pars compacta (SNC) with debilitating motor symptoms. Current treatments for PD afford symptomatic relief with no prevention of disease progression. Due to the antioxidant and neuroprotective potential of sinapic acid, this study was conducted to evaluate whether this agent could be of benefit in an experimental model of early PD in rat. Unilateral intrastriatal 6-hydroxydopamine (6-OHDA)-lesioned rats were pretreated p.o. with sinapic acid at doses of 10 or 20 mg/kg. One week after surgery, apomorphine caused significant contralateral rotations, a significant reduction in the number of Nissl-stained and tyrosine hydroxylase (TH)-positive neurons and a significant increase of iron reactivity on the left side of SNC. Meanwhile, malondialdehyde (MDA) and nitrite levels in midbrain homogenate significantly increased and activity of superoxide dismutase (SOD) significantly reduced in the 6-OHDA-lesioned group. In addition, sinapic acid at a dose of 20 mg/kg significantly improved turning behavior, prevented loss of SNC dopaminergic neurons, lowered iron reactivity, and attenuated level of MDA and nitrite. These results indicate the neuroprotective potential of sinapic acid against 6-OHDA neurotoxicity that is partially due to the attenuation of oxidative stress and possibly lowering nigral iron level.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Parkinson’s disease (PD) is a rather common movement disorder with neurodegenerative nature caused by the selective loss of dopaminergic neurons within mesencephalic substantia nigra pars compacta (SNC). Progressive loss of these neurons leads to reduced production of dopamine (DA) and impairment of motor skills in afflicted patients. This finally leads to development of tremor, bradykinesia, rigidity and postural imbalance (Fahn 2003; Halliday and McCann 2010). The current treatment for PD is mainly limited to relief of symptoms and to delay the progression of neurodegenerative process. L-DOPA is currently the most effective drug against the motor symptoms of PD. Although initial treatment with L-DOPA could markedly improve PD symptoms, however, with progression of disease, L-DOPA loses its efficiency and effectiveness. In addition, chronic use of L-DOPA is associated with the development of some side-effects such as fluctuations in motor response and dyskinesia (Bargiotas and Konitsiotis 2013; Smith et al. 2012). Therefore, neuroprotective strategies are proposed to slow down the pathogenic process of PD (Baluchnejadmojarad et al. 2010). There is increasing evidence that oxidative stress due to increased generation of reactive oxygen species is strongly involved in the pathogenesis of PD, suggesting that pharmacological targeting of the antioxidant machinery may be of benefit in PD therapy (De Araujo et al. 2011; Roghani et al. 2010). Iron accumulation has a pivotal role in some neurological disorders like PD. The elevated iron level in SNC region of PD patients leads to increased oxidative stress burden and enhanced aggregation of intracellular α-synuclein as a pathological mechanism, finally leading to oxidative damage of dopaminergic neurons (Weinreb et al. 2013).
Natural antioxidant agents derived from plants could reduce oxidative stress, to chelate deleterious iron and to attenuate inflammation and in this way attenuate dopaminergic neurodegeneration in PD (Datla et al. 2007). Sinapic acid is a phenylpropanoid compound that is found in various herbal materials including cereals, nuts, oil seeds, and berries (Shin et al. 2013). Sinapic acid has shown antioxidant activity (Roy and Mainzen Prince 2013), metal chelating potential (Pari and Mohamed Jalaludeen 2011), anti-inflammatory (Yun et al. 2008) and neuroprotective properties (Lee et al. 2012). This compound also exhibits protective potential against lysosomal dysfunction (Roy and Stanely Mainzen Prince 2012) and protects rat liver from carbon tetrachloride-induced inflammation, most likely by acting as a free radical scavenger and modulator of NF-κB activation and proinflammatory cytokine expression (Shin et al. 2013). However, the diverse pharmacological properties of sinapic acid remain to be elucidated. The present study was undertaken to investigate the neuroprotective effect of sinapic acid pretreatment in 6-hydroxydopamine (6-OHDA)-induced model of PD in rat.
Materials and methods
Animals
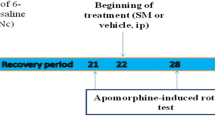
Adult male Wistar rats, weighing 200–260 g (n = 72) (Pasteur’s Institute, Iran) were housed in a temperature-controlled colony room under light/dark cycle with free access to food and water. Procedures involving animals and their care were conducted in conformity with NIH guidelines for the care and use of laboratory animals. The animals were held in the colony room for at least one week before being tested. Only rats not showing any biased rotational behavior (net rotations less than 30/h) following intraperitoneal injection of apomorphine hydrochloride (2 mg/kg) (Sigma-Aldrich, USA) were selected for the present study. The animals were randomly divided into six groups: sham-operated group, sham groups receiving sinapic acid at doses of 10 and 20 mg/kg, lesion group (6-OHDA), and lesion groups receiving sinapic acid at doses of 10 and 20 mg/kg. Unilateral intrastriatal 6-OHDA (Sigma Chemical, USA) injection (left side) was performed through a 5 μl Hamilton syringe on anesthetized rats (ketamine 80 mg/kg and xylazine 10 mg/kg, i.p.) using stereotaxic apparatus (Stoelting, USA) at the coordinates: L −3 mm, AP +9.2 mm, V + 4.5 mm from the center of the interaural line, according to the atlas of Paxinos and Watson (Paxinos and Watson 1986). At the end of injection, the needle was left in place for an additional 5 min and then withdrawn at a rate of 1 mm/min. The lesion group received a single injection of 5 μl of 0.9 % saline containing 2.5 μg/μl of 6-hydroxydopamine-HCL (6-OHDA, Sigma-Aldrich, USA) and 0.2 % ascorbic acid (W/V) at a rate of 1 μl/min. The sham group received an identical volume of ascorbate-saline solution. Sinapic acid (Sigma-Aldrich, USA) was administered p.o. using rodent gavage dissolved in 10 % Cremophore (Sigma-Aldrich, USA) at doses of 10 or 20 mg/kg. Dose of sinapic acid was chosen according to its protective effect against tetrachloride-induced hepatic injury in rats (Shin et al. 2013). Daily administration of sinapic acid was started 2 days before surgery at an interval of 24 h till one h pre-surgery.
Behavioral testing
All the animals (n = 72) were tested for rotational behavior by apomorphine hydrochloride (2 mg/kg, i.p.) 4–5 days before surgery (baseline) and 1 week after surgery. The rotations were measured according to a method as described previously (Roghani et al. 2010). Briefly, the animals were allowed to habituate for 10 min and then 1 min after the injection, full rotations were counted in a cylindrical container (a diameter of 33 cm and a height of 35 cm) at 10-min uninterrupted intervals for 60 min in a dimly-lighted and quiet room. Net number of rotations was defined as the positive scores minus the negative scores.
Determination of midbrain MDA concentration
Half of the rats (n = 6 for each group) were anesthetized with ketamine (150 mg/kg), decapitated, brains were removed, anterior third block of left midbrain was blotted dry, weighed, then made into 5 % tissue homogenate in ice-cold 0.9 % saline solution, centrifuged at 4 °C, obtained supernatant was aliquotted, then stored at −70 °C until assayed. The MDA concentration (thiobarbituric acid reactive substances, TBARS) in the supernatant was measured as described before (Roghani et al. 2010). Briefly, trichloroacetic acid and TBARS reagent were added to supernatant, then mixed and incubated at boiling water for 90 min. After cooling on ice, samples were centrifuged at 1,000 × g for 10 min and the absorbance of the supernatant was read at 532 nm. TBARS results were expressed as MDA equivalents using tetraethoxypropane as standard.
Measurement of midbrain SOD activity
The supernatant of midbrain homogenate was obtained as described earlier. SOD activity measurement was according to previous works (Roghani et al. 2010). Briefly, supernatant was incubated with xanthine and xanthine oxidase in potassium phosphate buffer (pH 7.8, 37 °C) for 40 min and Nitroblue tetrazolium (NBT) was added. Blue formazan was then monitored spectrophotometrically at 550 nm. The amount of protein that inhibited NBT reduction to 50 % maximum was defined as 1 nitrite unit (NU) of SOD activity.
Assay of midbrain nitrite concentration
Supernatant nitrite content was assayed by the Griess method according to previous studies (Baluchnejadmojarad and Roghani 2011). Because NO is a compound with a short half-life and is rapidly converted to the stable end products nitrate (NO3−) and nitrite (NO2−), the principle of the assay is the conversion of nitrate into nitrite by cadmium and followed by color development with Griess reagent (containing sulfanilamide and N-naphthyl ethylenediamine) in acidic medium. The total nitrite was measured by Griess reaction. The absorbance was determined at 540 nm with a spectrophotometer.
Protein assay
The protein content of the supernatant was measured with Bradford method using bovine serum albumin (Sigma-Aldrich, USA) as the standard (Bradford 1976).
Histological study
The remaining animals (n = 6 for each group) in each group were randomly used for histological assessment. At the end of behavioral experiments, the rats were deeply anesthetized with a high dose of ketamine (150 mg/kg) and perfused through the ascending aorta with 50–100 ml of 0.9 % saline followed by 100–200 ml of fixative solution containing 4 % paraformaldehyde in 0.1 M phosphate buffer (PB, pH 7.4) followed by 100 ml of 0.1 M PB containing 10 % sucrose. Following perfusion, the brains were removed from the skull, blocks of forebrain and brainstem were prepared, and after final steps of preparation (immersion in 30 % sucrose solution in 0.1 M PB for 2–3 days), sections were cut at a thickness of 30 μm on a freezing microtome (Leica, Germany) and collected in PB (0.05 M). Every third section was Nissl-stained with 0.1 % cresyl violet (Sigma-Aldrich, USA). The other sections were alternately used for the following histochemical experiments.
Tyrosine hydroxylase (TH) immunohistochemistry
Tyrosine hydroxylase (TH) is the rate limiting enzyme for dopamine synthesis and is usually used as a selective marker of dopaminergic neurons (Zhou et al. 2013). The sections on gelatin-coated slides were incubated in 0.1 % sodium borohydride in phosphate-buffered saline (PBS, pH 7.4), methanol containing 0.03 % H2O2, then in Triton X100 (0.03 %) and 0.1 % bovine serum albumin in PBS, with rinsing 2–3 times in the PBS between between the steps in a humidified chamber in darkness at room temperature. Then, sections were incubated with primary polyclonal rabbit anti-TH antibody (Abcam, Germany; 1/200) in PBS at darkness overnight and then with secondary anti-rabbit IgG-peroxidase antibody (HRP bound) raised in goat (Abcam, Germany; 1/200) in PBS for two hours. To reveal the bound peroxidase, the sections were incubated in diaminobenzidine tetrahydrochloride (DAB; 10 mg/20 ml of PBS and 0.03 % H2O2) for 3–5 min, rinsed, dehydrated, cleared and coverslipped.
Iron staining
Iron staining was conducted as described previously by Wang et al. (Wang et al. 2012). For this purpose, sections were soaked for 30 s in research grade water before staining, then incubated for 30 min in a freshly prepared solution of equal parts of 2 % HCL and 2 % potassium ferrocyanide. After washing with 0.01 M PBS, sections were immersed in a solution containing 99 % methanol and 1 % hydrogen peroxide for 20 min to quench endogenous peroxidase activity and then incubated in a solution of DAB containing H2O2.
Neuronal counting
For each animal, mesencephalic sections (Interaural 2.9–4.2 mm) were examined by a method as described previously (Baluchnejadmojarad et al. 2010). Briefly, Nissl-stained and/or TH-immunoreactive neurons of the SNC were counted manually (Light microscopy; X400) using a superimposed grid to facilitate the procedure. At least 2 sections representative of each of 4 Paxinos-Watson planes (4.2, 3.7, 3.2, 2.97; Interaural) were examined. Since there is much variations between the rats regarding neuronal population if certain area of the SNC is selected for counting, we preferred to count the Nissl-stained neurons within the entire extent of the left side of SNC. Meanwhile, neurons with stained nuclei and visible nucleoli were included in the counting. In addition, intensity of reactivity for iron within anterior one-third of midbrain was also evaluated as follows: no reactivity 0, very mild reactivity 1, mild reactivity 2, moderate reactivity 3, strong reactivity 4, and very strong reactivity 5. All counting and assessments were done blind to the treatments received.
Statistical analysis
All data were expressed as mean ± S.E.M. For the drug-induced rotational behavior, the non-parametric Kruskall-Wallis test was used. Inter-group differences for values of neuronal numbers for the injected side and biochemical assays were found out using one-way ANOVA followed by Tukey’s post-hoc test. In all analyses, the null hypothesis was rejected at a level of 0.05.
Results
In this study, the effect of oral administration of sinapic acid pretreatment at doses 10 and 20 mg/kg was assessed on apomorphine-induced rotational behavior for 1 h (Fig. 1). There were no significant differences among the groups before the surgery. In contrast, apomorphine caused a very significant contralateral turning in the rats of 6-OHDA group (p < 0.0001) and induced less significant rotations in 6-OHDA + Sinapic acid10 and 6-OHDA + Sinapic acid20 groups (p < 0.001 and p < 0.01, respectively) versus Sham group. In addition, the groups 6-OHDA + Sinapic acid10 and 6-OHDA + Sinapic acid20 significantly showed lower rotations (p < 0.05 and p < 0.01, respectively) versus 6-OHDA group.

Total net number of apomorphine-induced rotations/1 h at 1st week post-surgery in sham and 6-OHDA-lesioned groups. Note that the positive values indicate contralateral rotations. 6-OHDA stands for the neurotoxin 6-hydroxydopamine. (n = 12 for each group) * p < 0.01, ** p < 0.001, *** p < 0.0001 (versus Sham), ¥ p < 0.05, ¥¥ p < 0.01 (versus 6-OHDA)
The results of Nissl staining (Fig. 2) showed that there was no significant difference amongst sham, sham + Sinapic acid10 and Sinapic acid20 groups regarding the number of neurons on the left side of SNC. In this respect, a significant reduction was observed in 6-OHDA group (p < 0.01) and no significant reduction was noted for sinapic acid-pretreated 6-OHDA groups. Meanwhile, number of Nissl-stained neurons on the left side of SNC was significantly higher in 6-OHDA + Sinapic acid20 versus 6-OHDA group (p < 0.05). In this respect, number of Nissl-stained neurons were significantly higher in 6-OHDA + Sinapic acid20 versus 6-OHDA group at planes 3.2 (p < 0.01) and 3.7 (p < 0.05) with no such effect at planes 2.96 and 4.2.

Total number of Nissl-stained neurons on the left side of substantia nigra pars compacta (SNC) at different interaural stereotaxic planes (top panel) and its averaged number at all planes (bottom panel) and coronal section photomicrographs in different groups 1 week post-surgery. 6-OHDA and SNR stand for the neurotoxin 6-hydroxydopamine and substantia nigra pars reticulate, respectively. (n = 6 for each group) * p < 0.05, ** p < 0.01, *** p < 0.005 (versus Sham), ¥ p < 0.05, ¥¥ p < 0.01 (versus 6-OHDA)
TH immunohistochemical results (Fig. 3) showed that there was no significant difference amongst the groups sham, sham + Sinapic acid10 and Sinapic acid20 regarding TH-positive neurons on the left side of SNC. In addition, a significant reduction of TH-positive neurons was observed in 6-OHDA group (p < 0.005) as compared to Sham group. Meanwhile, a less significant reduction of such neurons was observed in 6-OHDA + Sinapic acid10 (p < 0.01) and 6-OHDA + Sinapic acid20 (p < 0.05) groups when compared to 6-OHDA group. Meanwhile, number of TH-positive neurons on the left side of SNC was significantly higher in 6-OHDA + Sinapic acid10 (p < 0.05) and 6-OHDA + Sinapic acid20 versus 6-OHDA groups (p < 0.01).

Averaged total number of TH-positive neurons on the left side of substantia nigra pars compacta (SNC) (top panel) and coronal section photomicrographs showing its immunoreactivity (bottom panel) in different groups 1 week post-surgery. 6-OHDA and SNR stand for the neurotoxin 6-hydroxydopamine and substantia nigra pars reticulate, respectively. (n = 6 for each group) * p < 0.05, ** p < 0.01, *** p < 0.005 (versus Sham), ¥ p < 0.05, ¥¥ p < 0.01 (versus 6-OHDA)
Iron-staining histochemical data (Fig. 4) showed that there was a significant increase in iron reactivity on the left side of midbrain of 6-OHDA (p < 0.01) and 6-OHDA + Sinapic acid10 (p < 0.05) groups versus sham and such reactivity was significantly lower in 6-OHDA + Sinapic acid20 group as compared to 6-OHDA (p < 0.05).

Averaged reactivity intensity for iron on the anterior one-third of the left midbrain (top panel) and coronal section photomicrographs showing its reactivity (bottom panel) in different groups 1 week post-surgery. 6-OHDA and SNR stand for the neurotoxin 6-hydroxydopamine and substantia nigra pars reticulate, respectively. (n = 5 for each group) * p < 0.05, ** p < 0.01 (versus Sham), ¥ p < 0.05 (versus 6-OHDA)
Measurement of some oxidative stress markers (Fig. 5) indicated that 6-OHDA group showed a significant elevation of MDA (as a marker of lipid peroxidation) (p < 0.05) and nitrite content (p < 0.05) and a significant reduction of SOD activity (p < 0.05) and pretreatment of 6-OHDA-lesioned rats with sinapic acid at a dose of 10 mg/kg significantly improved MDA level (p < 0.05). Meanwhile, sinapic acid administration at a dose of 20 mg/kg significantly attenuated the increased level of MDA and nitrite content (p < 0.05) with no significant improvement of SOD activity.

MDA content (top panel), nitrite content (middle panel) and superoxide dismutase (SOD) activity (bottom panel) in midbrain homogenate from different groups. 6-OHDA stands for 6-hydroxydopamine. (n = 5–6 for each group) * p < 0.05 (versus Sham), ¥ p < 0.05 (versus 6-OHDA)
Discussion
In this study, we demonstrated that sinapic acid could dose-dependently mitigate apomorphine-induced rotational behavior, prevent loss of SNC dopaminergic neurons, lower number of iron-positive cells, and mitigate level of MDA and nitrite in 6-OHDA-lesioned rats.
The preferential and progressive degeneration of dopaminergic neurons within SNC in PD is the main causative factor for the forthcoming outcome (Hattori 2004; Schapira and Jenner 2011). The neurotoxin 6-OHDA is usually used for induction of PD in rats, which also causes selective degeneration of dopaminergic neurons (Schober 2004). Intrastriatal injection of 6-OHDA leads to unilateral damage of the nigrostriatal dopaminergic system that is followed by a reduction in the striatal dopamine level and an upregulation of dopaminergic post-synaptic receptors at the same side. These changes produce a prominent functional and motor asymmetry that can be evaluated by dopaminergic agonists like apomorphine (Jalali-Nadoushan and Roghani 2013). These rotations are considered as reliable indicators of nigrostriatal dopamine depletion (Shapiro et al. 1987). In this study, a significant attenuation of the apomorphine-induced rotations was observed in sinapic acid-pretreated 6-OHDA-lesioned group 1 week after surgery. The observed attenuation of rotational behavior in sinapic acid-pretreated lesioned group could be attributed to possible neuroprotective effect of sinapic acid against degeneration of dopaminergic neurons and maintenance of striatal dopamine at a level that is not accompanied with a marked rotational behavior.
Compelling evidence implicates oxidative stress, mitochondrial deficiency, and inflammation as important pathogenic factors in PD (Tapias et al. 2013). Oxidative stress is thought to play a significant role in the development and progression of neurodegenerative diseases like PD (Chakraborty et al. 2013). Oxidative stress enhancement also plays a pivotal role in 6-OHDA-neurotoxicity in experimental models of PD (Baluchnejadmojarad et al. 2010). Oxidative stress could affect the survival of dopaminergic neurons in PD. Neurons mostly depend on energy produced by mitochondria and are simultaneously faced with oxidative stress burden as well as increased levels of free iron, which can promote OH generation (Foley and Riederer 2000). Overload of the free radical formation may lead to cell death. In addition, auto-oxidation of dopamine may produce dopamine quinine (Lotharius and Brundin 2002). Formation of species such as semiquinones and other free radicals could especially damage nucleic acids, proteins, and membrane lipid components (Von Bohlen und Halbach et al. 2004). Therefore, some therapeutic approaches are aimed to attenuate oxidative stress. In addition, free radical scavengers may be helpful in prolonging survival time of dopaminergic neurons (Chen and Le 2006). In this respect, sinapic acid could attenuate neuronal damage and loss through counteracting oxidative stress, possibly via regulating antioxidant defense system as well as inhibition of free radical generation (Pari and Mohamed Jalaludeen 2011). In our study, sinapic acid was able to counteract the enhanced oxidative stress burden following 6-OHDA, as was observed by a lower midbrain level of MDA and nitrite.
Furthermore, part of toxicity of the neurotoxin 6-OHDA could be due to its potential to release ferritin-bound iron and increase its concentration at the site of neurodegeneration (Andringa et al. 2000). Iron chelation might be a promising therapeutic target for PD (Le 2014). Part of beneficial effect of sinapic acid in this study could be attributed to its potential as a possible iron-chelating agent, which itself warrants further investigation.
Neuroinflammation is another important feature in PD (Bisaglia et al. 2014). Pro-inflammatory cytokines released from glial cells could stimulate nitric oxide production and exert a deleterious effect on dopaminergic neurons by activating receptors that contain intra-cytoplasmic death domains involved in apoptotic pathway (Sriram and O’Callaghan 2007). Since sinapic acid has shown anti-inflammatory property (Shin et al. 2013) via NF-kappaB inactivation (Yun et al. 2008), sinapic acid may have decreased the level of inflammatory mediators within the brain, which itself contributes to neuroprotection in 6-OHDA-induced PD in rats, as observed in our study. This needs further research in future works.
In this study, we used sinapic acid as a neuroprotective agent in an early model of PD in rats. This compound is a major free phenolic acid in rapeseed meal, with its majority found in the esterified form of sinapine. Experiments conducted to delineate the effect of its dietary consumption on broiler chickens have shown that this compound is a safe agent with easy digestion and rapid absorption and with no toxic effect on some tissues like skeletal muscle, heart, liver, kidneys, or brain (Qiao et al. 2008).
Taken together, obtained results indicate the neuroprotective potential of sinapic acid against 6-OHDA neurotoxicity that is partially due to the attenuation of oxidative stress and nigral iron level. However, further studies are compulsory to explore its exact mode of action.
References
Andringa G, van Oosten RV, Unger W, Hafmans TG, Veening J, Stoof JC, Cools AR (2000) Systemic administration of the propargylamine CGP 3466B prevents behavioural and morphological deficits in rats with 6-hydroxydopamine-induced lesions in the substantia nigra. Eur J Neurosci 12:3033–3043
Baluchnejadmojarad T, Roghani M (2011) Chronic epigallocatechin-3-gallate ameliorates learning and memory deficits in diabetic rats via modulation of nitric oxide and oxidative stress. Behav Brain Res 224:305–310
Baluchnejadmojarad T, Roghani M, Mafakheri M (2010) Neuroprotective effect of silymarin in 6-hydroxydopamine hemi-parkinsonian rat: involvement of estrogen receptors and oxidative stress. Neurosci Lett 480:206–210
Bargiotas P, Konitsiotis S (2013) Levodopa-induced dyskinesias in Parkinson’s disease: emerging treatments. Neuropsychiatr Dis Treat 9:1605–1617
Bisaglia M, Filograna R, Beltramini M, Bubacco L (2014) Are dopamine derivatives implicated in the pathogenesis of Parkinson’s disease? Ageing Res Rev 13C:107–114
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Chakraborty S, Bornhorst J, Nguyen TT, Aschner M (2013) Oxidative stress mechanisms underlying Parkinson’s disease-associated neurodegeneration in C. elegans. Int J Mol Sci 14:23103–23128
Chen S, Le W (2006) Neuroprotective therapy in Parkinson disease. Am J Ther 13:445–457
Datla KP, Zbarsky V, Rai D, Parkar S, Osakabe N, Aruoma OI, Dexter DT (2007) Short-term supplementation with plant extracts rich in flavonoids protect nigrostriatal dopaminergic neurons in a rat model of Parkinson’s disease. J Am Coll Nutr 26:341–349
De Araujo DP, Lobato Rde F, Cavalcanti JR, Sampaio LR, Araujo PV, Silva MC, Neves KR, Fonteles MM, Sousa FC, Vasconcelos SM (2011) The contributions of antioxidant activity of lipoic acid in reducing neurogenerative progression of Parkinson’s disease: a review. Int J Neurosci 121:51–57
Fahn S (2003) Description of Parkinson’s disease as a clinical syndrome. Ann N Y Acad Sci 991:1–14
Foley P, Riederer P (2000) Influence of neurotoxins and oxidative stress on the onset and progression of Parkinson’s disease. J Neurol 247(Suppl 2):II82–94
Halliday GM, McCann H (2010) The progression of pathology in Parkinson’s disease. Ann N Y Acad Sci 1184:188–195
Hattori N (2004) Etiology and pathogenesis of Parkinson’s disease: from mitochondrial dysfunctions to familial Parkinson’s disease. Rinsho Shinkeigaku 44:241–262
Jalali-Nadoushan M, Roghani M (2013) Alpha-lipoic acid protects against 6-hydroxydopamine-induced neurotoxicity in a rat model of hemi-parkinsonism. Brain Res 1505:68–74
Le W (2014) Role of iron in UPS impairment model of Parkinson’s disease. Parkinsonism Relat Disord 20(Suppl 1):S158–161
Lee HE, Kim DH, Park SJ, Kim JM, Lee YW, Jung JM, Lee CH, Hong JG, Liu X, Cai M, Park KJ, Jang DS, Ryu JH (2012) Neuroprotective effect of sinapic acid in a mouse model of amyloid beta (1–42) protein-induced Alzheimer’s disease. Pharmacol Biochem Behav 103:260–266
Lotharius J, Brundin P (2002) Pathogenesis of Parkinson’s disease: dopamine, vesicles and alpha-synuclein. Nat Rev Neurosci 3:932–942
Pari L, Mohamed Jalaludeen A (2011) Protective role of sinapic acid against arsenic: induced toxicity in rats. Chem Biol Interact 194:40–47
Paxinos G, Watson C (1986) The rat brain in stereotaxic coordinates, 2nd edn. Academic, San Diego
Qiao HY, Dahiya JP, Classen HL (2008) Nutritional and physiological effects of dietary sinapic acid (4-hydroxy-3,5-dimethoxy-cinnamic acid) in broiler chickens and its metabolism in the digestive tract. Poult Sci 87:719–726
Roghani M, Niknam A, Jalali-Nadoushan MR, Kiasalari Z, Khalili M, Baluchnejadmojarad T (2010) Oral pelargonidin exerts dose-dependent neuroprotection in 6-hydroxydopamine rat model of hemi-parkinsonism. Brain Res Bull 82:279–283
Roy SJ, Mainzen Prince PS (2013) Protective effects of sinapic acid on cardiac hypertrophy, dyslipidaemia and altered electrocardiogram in isoproterenol-induced myocardial infarcted rats. Eur J Pharmacol 699:213–218
Roy SJ, Stanely Mainzen Prince P (2012) Protective effects of sinapic acid on lysosomal dysfunction in isoproterenol induced myocardial infarcted rats. Food Chem Toxicol 50:3984–3989
Schapira AH, Jenner P (2011) Etiology and pathogenesis of Parkinson’s disease. Mov Disord 26:1049–1055
Schober A (2004) Classic toxin-induced animal models of Parkinson’s disease: 6-OHDA and MPTP. Cell Tissue Res 318:215–224
Shapiro RM, Glick SD, Camarota NA (1987) A two-population model of rat rotational behavior: effects of unilateral nigrostriatal 6-hydroxydopamine on striatal neurochemistry and amphetamine-induced rotation. Brain Res 426:323–331
Shin DS, Kim KW, Chung HY, Yoon S, Moon JO (2013) Effect of sinapic acid against carbon tetrachloride-induced acute hepatic injury in rats. Arch Pharm Res 36:626–633
Smith Y, Wichmann T, Factor SA, DeLong MR (2012) Parkinson’s disease therapeutics: new developments and challenges since the introduction of levodopa. Neuropsychopharmacology 37:213–246
Sriram K, O’Callaghan JP (2007) Divergent roles for tumor necrosis factor-alpha in the brain. J Neuroimmune Pharmacol 2:140–153
Tapias V, Cannon JR, Greenamyre JT (2013) Pomegranate juice exacerbates oxidative stress and nigrostriatal degeneration in Parkinson’s disease. Neurobiol Aging 35:1162–1176
Von Bohlen und Halbach O, Schober A, Krieglstein K (2004) Genes, proteins, and neurotoxins involved in Parkinson’s disease. Prog Neurobiol 73:151–177
Wang J, Xu H, Jiang H, Du X, Sun P, Xie J (2012) Neurorescue effect of rosmarinic acid on 6-hydroxydopamine-lesioned nigral dopamine neurons in rat model of Parkinson’s disease. J Mol Neurosci 47:113–119
Weinreb O, Mandel S, Youdim MB, Amit T (2013) Targeting dysregulation of brain iron homeostasis in Parkinson’s disease by iron chelators. Free Radic Biol Med 62:52–64
Yun KJ, Koh DJ, Kim SH, Park SJ, Ryu JH, Kim DG, Lee JY, Lee KT (2008) Anti-inflammatory effects of sinapic acid through the suppression of inducible nitric oxide synthase, cyclooxygase-2, and proinflammatory cytokines expressions via nuclear factor-kappaB inactivation. J Agric Food Chem 56:10265–10272
Zhou ZD, Refai FS, Xie SP, Ng SH, Chan CH, Ho PG, Zhang XD, Lim TM, Tan EK (2013) Mutant PINK1 upregulates tyrosine hydroxylase and dopamine levels, leading to vulnerability of dopaminergic neurons. Free Radic Biol Med 68C:220–233
Acknowledgments
This work was the results of a Ph. D. thesis project financially funded and supported by Science and Research Branch of the Islamic Azad University (Tehran, Iran) in 2012. The authors would also like to appreciate Fariba Ansari and Maryam Sharayely for their great technical assistance.
Conflict of interest
There is no conflict of interest.
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article

Cite this article
Zare, K., Eidi, A., Roghani, M. et al. The neuroprotective potential of sinapic acid in the 6-hydroxydopamine-induced hemi-parkinsonian rat. Metab Brain Dis 30, 205–213 (2015). https://doi.org/10.1007/s11011-014-9604-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11011-014-9604-6