
Abstract
Thyroid hormones (THs) are recognized as key metabolic hormones, and the metabolic rate increases in hyperthyroidism, while it decreases in hypothyroidism. The aim of this work was to investigate how changes in metabolism induced by THs could affect the activities of acetylcholinesterase (AChE), (Na+, K+)- and Mg2+-ATPase in the hypothalamus and the cerebellum of adult rats. Hyperthyroidism was induced by subcutaneous administration of thyroxine (25μg/100 g body weight) once daily for 14 days, while hypothyroidism was induced by oral administration of propylthiouracil (0.05%) for 21 days. All enzyme activities were evaluated spectrophotometrically in the homogenated brain regions of 10 three-animal pools. Neither hyper-, nor hypothyroidism had any effect on the examined hypothalamic enzyme activities. In the cerebellum, hyperthyroidism provoked a significant decrease in both the AChE (−23%, p < 0.001) and the Na+, K+-ATPase activities (−26%, p < 0.001). Moreover, hypothyroidism had a similar effect on the examined enzyme activities: AChE (−17%, p < 0.001) and Na+, K+-ATPase (−27%, p < 0.001). Mg2+-ATPase activity was found unaltered in both the hyper- and the hypothyroid brain regions. In conclusion: neither hyper-, nor hypothyroidism had any effect on the examined hypothalamic enzyme activities. In the cerebellum, hyperthyroidism provoked a significant decrease in both the AChE and the Na+, K+-ATPase activities. The decreased (by the THs) Na+, K+-ATPase activities may increase the synaptic acetylcholine release, and thus, could result in a decrease in the cerebellar AChE activity. Moreover, the above TH-induced changes may affect the monoamine neurotransmitter systems.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Thyroid hormones (THs) are recognized as key metabolic hormones that play a critical role in brain development (and not only), mediating important effects within the central nervous system (CNS) throughout life (Bernal and Nunez, 1995; Bradley et al., 1989; Cook et al., 1992; Gullo et al., 1987; Mellstrom et al., 1991; Oppenheimer and Schwartz, 1997; Rastogi et al., 1977). Moreover, adult-onset thyroid dysfunction is associated with both neurological and behavioural abnormalities (DeGroot et al., 1984), emphasizing the importance of THs for normal brain function. Neurologically, hypothyroidism (Hypo) has been associated with cerebellar ataxia, confusion, delusions, memory impairment, hallucinations and psychotic behaviour. On the contrary, in the case of hyperthyroidism (Hyper), symptoms include irritability, nervousness, anxiety, sleep disturbances, tremulousness, delirium, stupor and even coma (Adams and Rosman, 1971; Hall et al., 1986).
Acetylcholinesterase (AChE, EC 3.1.1.7) is a cholinergic enzyme, the role of which is very important in the acetylcholine (ACh) cycle, including the release of ACh (Kouniniotou-Krontiri and Tsakiris, 1989). In addition, it was found that AChE is co-released from the dopaminergic neurons, implying an interaction between these two molecules which is important for the dopaminergic function (Klegeris et al., 1995). (Na+, K+)-ATPase (EC 3.6.1.3) is an enzyme implicated in neuronal excitability (Sastry and Phillis, 1977), metabolic energy production (Mata et al., 1980), as well as in the uptake and release of catecholamines (Bogdanski et al., 1968; Swann, 1984), serotonin (Hernandez, 1989) and glutamate (Lees et al., 1990). Moreover, the role of Mg2+-ATPase is to maintain high brain intracellular Mg2+, changes of which can control rates of protein synthesis and cell growth (Sanui and Rubin, 1982).
In a previous study (Carageorgiou et al., 2005), Mg2+-ATPase activity was found unaffected in the hyperthyroid rat whole brain, while AChE and Na+, K+-ATPase activities were found reduced. In contrast, Mg2+- and (Na+, K+)-ATPase activities were found to be increased in the hypothyroid rat brain, while AChE activity was found decreased.
The aim of this work was to assess the activities of AChE, (Na+, K+)-ATPase and Mg2+-ATPase in the hypothalamus and the cerebellum of adult rats with experimental hyper- and hypothyroidism.
Materials and methods
Animals
Albino Wistar adult male rats (six months old) were used in all experiments. The rats were housed four in a cage, at a constant room temperature (22±1°C) under a 12-h light: 12-h dark (light 08:00–20:00 h) cycle. Food and water were provided ad libitum. Animals were cared for in accordance with the principles of the “Guide to the Care and Use of Experimental Animals” (Committee on Care and Use of Laboratory Animals, 1985).
Experimental hyper- and hypothyroidism
Hyperthyroidism was induced in rats by thyroxine administration. l-Thyroxine (T4) (Sigma, St. Louis, MO, USA) was dissolved in 99% ethanol by adding a small volume (20 μl) of 25% NaOH and diluted 33 times by adding 0.9% NaCl to obtain a stock solution of 1 mg/ml. Before each injection, a fresh solution was made in 0.9% NaCl to obtain a concentration of T4 at 50 μg/ml. Thyroxine, 25 μg/100 g body weight was given subcutaneously once daily for 14 days. On the other hand, hypothyroidism was induced in rats by administration of 6-n-propyl-2-thiouracil in drinking water to a final concentration of 0.05% for 21 days. Each treatment results in a long-term moderate hyperthyroidism (Pantos et al., 1999) or hypothyroidism (Pantos et al., 2003). Two controls were used: a) saline controls (SC) that were treated with subcutaneous injections of normal saline given once daily for 14 days (control of hyperthyroid rats), and b) controls without any treatment (NTC) for 21 days (control for hypothyroid rats).
Tissue preparation
The animals were sacrificed by decapitation and the brain regions were rapidly removed. The tissue was homogenized in 10 vol. ice-cold (0–4°) medium containing 50 mM Tris (hydroxymethyl) aminomethane-HCl (Tris-HCl), pH 7.4 and 300 mM sucrose, using an ice-chilled glass homogenizing vessel at 900 rpm (4–5 strokes). Then, the homogenate was centrifuged at 1,000×g for 10 min. to remove nuclei and debris (Tsakiris et al., 2000; Tsakiris, 2001). In the resulting supernatant, the protein content was determined according to the method of Lowry et al. (1951) and then the enzyme activities were measured.
Determination of brain acetylcholinesterase activity
AChE activity was determined by following the hydrolysis of acetylthiocholine according to the method of Ellman et al. (1961), as described by Tsakiris (2001). The incubation mixture (1 ml) contained 50 mM Tris-HCl, pH 8, 240 mM sucrose and 120 mM NaCl. The protein concentration of the incubation mixture was 80–100 μg/ml. The reaction was initiated after addition of 0.03 ml of 5,5′-dithionitrobenzoic acid (DTNB) and 0.05 ml of acetylthiocholine iodide, which was used as substrate. The final concentration of DTNB and substrate were 0.125 and 0.5 mM, respectively. The reaction followed spectrophotometrically by the increase of absorbance (Δ\(\overline {OD}\)) at 412 nm.
Determination of Na+, K+-ATPase and Mg2+-ATPace activities
(Na+, K+)-ATPase activity was calculated from the difference between total ATPase activity (Na+, K+, Mg2+-dependent ATPase) and Mg2+-dependent ATPase activity. Total ATPase activity was assayed in an incubation medium consisting of 50 mM Tris-HCl, pH 7.4, 120 mM NaCl, 20 mM KCl, 4 mM MgCl2, 240 mM sucrose, 1 mM ethylenediamine tetraacetic acid K2-salt (K+-ΕDΤΑ), 3 mM disodium ATP and 80–100 μg protein of the homogenate in a final volume of 1 ml. Ouabain (1 mM) was added in order to determine the activity of Mg2+-ATPase. The reaction was started by adding ATP and stopped after an incubation period of 20 min by addition of 2 ml mixture of 1% lubrol and 1% ammonium molybdate in 0.9 M H2SO4 (Bowler and Tirri, 1974; Tsakiris, 2001). The yellow colour which developed was read at 390 mM.
Statistical analysis
The data were analyzed by a two-tailed Student’s t-test. The values of p < 0.05 were considered statistically significant.
Results
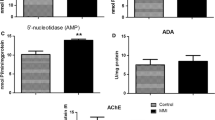
Table 1 presents the effects of hyper- and hypothyroidism on the hypothalamic AChE, (Na+, K+)- and Mg2+-ATPase activities in adult rats. The examined rat hypothalamic enzymes do not seem to be affected by the experimental procedures followed. The effects of hyper- and hypothyroidism on the cerebellar AChE, (Na+, K+)- and Mg2+-ATPase activities in adult rats, are shown in Table 2. In the cerebellum, hyperthyroidism provoked a significant decrease in both the AChE (−23%, p < 0.001) and the Na+, K+-ATPase activities (−26%, p < 0.001). Moreover, hypothyroidism had a similar effect on the examined enzyme activities: AChE (−17%, p < 0.001) and Na+, K+-ATPase (−27%, p < 0.001). Mg2+-ATPase activity was found unaltered in both the hyper- and the hypothyroid brain regions.
Discussion
The overall analysis of our data revealed that THs affected the examined parameters only in the cerebellum. None of the hypothalamic examined parameters was affected by any of the experimental procedures followed. This is in accordance with the fact that no findings have been reported so far, concerning the function of the hypothalamic AChE, (Na+, K+)- and Mg2+-ATPase in hyper- or hypothyroidism. It may be suggested that the hypothalamus, because of its intimate relationship with both the endocrine and the autonomic system, appears to possess significant regulatory mechanisms, towards homeostatic functioning of its neuronal systems (Iversen et al., 2000).
The data concerning the effects induced by alterations of the thyroid state on adult rat brain AChE activity are limited. It appears that the present findings in the cerebellum (concerning AChE activity) are in accordance with previous findings (Carageorgiou et al., 2005) refereeing to the whole rat brain AChE activity (exhibiting a similar reduction in both hyper- and hypothyroid states).
It is known that choline acetyltransferase (ChAT) activity (a—considered to be—specific marker for cholinergic neurons) decreases in the cerebellum of hyper- and increases in the cerebellum of hypothyroid suckling rats (Virgili et al., 1991). The level of ChAT activity correlates with Purkinje cell size, supporting the concept of a neurotrophic role of ACh (Clos et al., 1989). Moreover, in comparison with euthyroid suckling rats, the density of muscarinic receptors in the cerebellum was found to be 40% higher in hyperthyroidism and 30% lower in thyroid deficiency (Patel et al., 1980). ChAT and AChE are transiently expressed together in functionally non-cholinergic Purkinje cells. In contrast with most regions of the CNS, the high ratio of ChAT to AChE activity suggests an enhanced synthesis of ACh (Clos et al., 1989). It may be suggested that hyperthyroid cerebellum exhibits a diminished ACh synthesis, compared to the euthyroid one, followed by an increased muscarinic receptor density. Providing that AChE is the degradative enzyme of ACh, one could speculate that thyroxine plays a neurotrophic role and enhances the increase of ACh concentration in the synaptic cleft by reducing AChE activity and increasing the muscarinic receptor synthesis. The observed reduced AChE activity in the hyperthyroid cerebellum (Table 2) might be the result of a thyroxine-induced increase of intracellular calcium (Ernest, 1989), which could result in synaptosomal membrane changes and modulation of lipid(s)-protein(s) interaction(s).
In hypothyroidism, the observed reduced AChE activity in the cerebellum (Table 2) may be the result of a long-term diminution of the number of the active enzyme molecules in the membrane, mainly due to metabolic reasons. This is in accordance with the findings of Ahmed et al. (1993), where AChE activity was found reduced (by 45%) in the cerebellum of hypothyroid adult rats. Moreover, Li et al. (1996) suggested that iodine and THs deficiency can affect the maturation of ChAT and disturb the transformation and maturation of various molecular types of AChE. In addition to the reported low muscarinic receptor density, the reduced AChE activity might contribute to the clinically known hypothyroid cerebellar ataxia (Aminoff et al., 2005). The subject needs further investigation.
The observed inhibition of cerebellar Na+, K+-ATPase in hyperthyroidism (Table 2) is in accordance with previous findings in whole rat brain (Carageorgiou et al., 2005), and may be due to the increase of intracellular calcium by L-T4 (Ernest, 1989). However, the hypothyroid cerebellar Na+, K+-ATPase activity was also found significantly decreased (Table 2), in contrast to the whole brain Na+, K+-ATPase activity that was previously reported to be enhanced (Carageorgiou et al., 2005). This region-specific finding seems to be in accordance with those of Atterwill et al. (1985), where hypothyroidism significantly reduced the Na+, K+-ATPase activity in the suckling rat cerebellum, but not in the forebrain. Moreover, Atterwill et al. (1985) observed a 20–45% reduction of the alpha (+) form of Na+, K+-ATPase (high ouabain affinity) against control cerebellum, compared to a 60–70% reduction in the activity of the alpha form of Na+, K+-ATPase (low ouabain affinity). The latter might be indicative of a retarded development of a selective cerebellar cell population, containing predominantly the alpha form of the enzyme.
It should be also noted that the decrease in Na+, K+-ATPase activity can enhance (at least in part) the ACh release (Meyer and Cooper, 1981), and thus, affect in a similar way the cerebellar AChE activity. Moreover, the observed alterations of Na+, K+-ATPase activity may modulate cerebellar catecholaminergic, serotoninergic and glutamatergic systems (Bogdanski et al., 1968; Hernandez, 1987; Lees et al., 1990, Swann, 1984), as well as neural excitability (Sastry and Phillis, 1977) and metabolic energy production (Mata et al., 1980). It must be also noted that in mature rat cerebellum, hypothyroidism produced a 61% increase of monoaminoxidase (MAO) activity (Ahmed et al., 1993).
The unaltered Mg2+-ATPase activity in the hyperthyroid cerebellar tissue is in accordance with our earlier whole brain findings (Carageorgiou et al., 2005), while the hypothyroid cerebellar enzyme activity was not found increased as expected. The latter could be indicative of a different (state- and region-dependent) Mg2+-ATPase behavior.
In conclusion: neither hyper-, nor hypothyroidism had any effect on the examined hypothalamic enzyme activities. In the cerebellum, hyperthyroidism provoked a significant decrease in both the AChE and the Na+, K+-ATPase activities. Moreover, hypothyroidism had a similar effect on the above enzyme activities. Mg2+-ATPase activity was found unaltered in both the hyper- and the hypothyroid brain regions. These effects could be due to region- and state-specific differences in metabolism (provoking changes in the synthesis, the maturation and the function of at least some of the above enzymes), as well as to the THs-induced alterations of the intracellular calcium concentration and the monoaminergic systems’ actions.
References
Adams RD, Rosman NP (1971) Hypothyroidism: neuromuscular system. In: Werner SC, Ingbar SH (eds). The thyroid. Academic Press, New York, pp. 901–910
Ahmed MT, Sinha AK, Pickard MR, Kim KD, Ekins RP (1993) Hypothyroidism in the adult rat causes brain region-specific biochemical dysfunction. J Endocrinol 138:299–305
Aminoff MJ, Greenberg DA, Simon RP (2005) Clinical neurology. Lange Medical Books/McGraw-Hill, New York, pp. 117–118
Atterwill CK, Reid J, Athayde CM (1985) Effect of thyroid status on the development of the different molecular forms of Na+,K+-ATPase in rat brain. Mol Cell Endocrinol 40:149–158
Bernal J, Nunez J (1995) Thyroid hormones and brain development. Eur J Endocrinol 133:390–398
Bogdanski DF, Tissari A, Brodie BB (1968) Role of sodium, potassium, ouabain and reserpine in uptake, storage and metabolism of biogenic amines in synaptosomes. Life Sci 7:419–428
Bowler K, Tirri R (1974) The temperature characteristics of synaptic membrane ATPases from immature and adult rat brain. J Neurochem 23:611–613
Bradley DJ, Young 3rd WS, Weinberger C (1989) Differential expression of alpha and beta thyroid hormone receptor genes in rat brain and pituitary. Proc Natl Acad Sci USA 86:7250–7254
Carageorgiou H, Pantos C, Zarros A, Mourouzis I, Varonos D, Cokkinos D, Tsakiris S (2005) Changes in antioxidant status, protein concentration, acetylcholinesterase, (Na+, K+)- and Mg2+-ATPase activities in the brain of hyper- and hypothyroid adult rats. Metabol Brain Dis 20:129–139
Clos J, Ghandour S, Eberhart R, Vincendon G, Gombos G (1989) The cholinergic system in developing cerebellum: Comparative study of normal, hypothyroid and underfed rats. Dev Neurosci 11:188–204
Committee on Care and Use of Laboratory Animals (1985) Guide for the care and use of laboratory animals. Institute of Laboratory Animal Resources, National Research Council, Washington, DC, p. 83
Cook CB, Kakucska I, Lechan RM, Koenig RJ (1992) Expression of thyroid hormone receptor beta 2 in rat hypothalamus. Endocrinology 130:1077–1079
DeGroot LJ, Larsen PR, Refetoff S, Stanbury JB (1984) The thyroid and its diseases. John Wiley, Brisbane
Ellman GL, Courtney KD, Andres Jr V, Featherstone RM (1961) A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem Pharmacol 7:88–95
Ernest S (1989) Model of gentamicin-induced nephrotoxicity and its amelioration by calcium and thyroxine. Med Hypotheses 30:195–202
Gullo D, Sinha AK, Woods R, Pervin K, Ekins RP (1987) Triiodothyronine binding in adult rat brain: compartmentation of receptor populations in purified neuronal and glial nuclei. Endocrinology 120:325–331
Hall RCW, Stickney S, Beresford TP (1986) Endocrine disease and behaviour. Integr Psychiat 4:122–135
Hernandez J (1987) Brain Na+, K+-ATPase activity possibly regulated by a specific serotonin receptor. Brain Res 408:399–402
Iversen S, Iversen L, Saper CB (2000) The autonomic nervous system and the hypothalamus. In: Kandel ER, Schwartz JH, Jessel TM (eds). Principles of neural science. McGraw-Hill, New York, pp. 960–981
Klegeris A, Korkina LG, Greenfield SA (1995) A possible interaction between acetylcholinesterase and dopamine molecules during autoxidation of the amine. Free Radic Biol Med 18:223–230
Kouniniotou-Krontiri P, Tsakiris S (1989) Time dependence of Li+ action on acetylcholinesterase activity in correlation with spontaneous quantal release of acetylcholine in rat diaphragm. Jnp J Physiol 39:429–440
Lees GJ, Lehmann A, Sandberg M, Hamberger A (1990) The neurotoxicity of ouabain, a sodium-potassium ATPase inhibitor, in the rat hippocampus. Neurosci Lett 120:159–162
Li Z, Yang R, Chen Z (1996) Effects of iodine and thyroid hormone deficiency during brain development on activities of cholinergic neuron-related enzymes in central nervous system of rats. Zhonghua Yu Fang Yi Xue Za Zhi 30:337–339
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Protein measurement with the folin phenol reagent. J Biol Chem 193:265–275
Mata M, Fink DJ, Gainer H, Smith CB, Davidsen L, Savakis H, Schwartz WJ, Sokoloff L (1980) Activity-dependent energy metabolism in rat posterior pituitary primarily reflects sodium pump activity. J Neurochem 34:213–215
Mellstrom B, Naranjo JR, Santos A, Gonzales AM, Bernal J (1991) Independent expression of the alpha and beta c-erbA genes in developing rat brain. Mol Endocrinol 5:1339–1350
Meyer ME, Cooper RJ (1981) Correlations between Na+, K+-ATPase activity and acetylcholine release in rat cortical synaptosomes. J Neurochem 36:467–475
Oppenheimer JH, Schwartz HL (1997) Molecular basis of thyroid hormone-dependent brain development. Endocr Rev 18:462–475
Pantos CI, Cokkinos DD, Tzeis SM, Malliopoulou V, Mourouzis IS, Carageorgiou HC, Limas C, Varonos D, Cokkinos DV (1999) Hyperthyroidism is associated with preserved preconditioning capacity but intensified and accelerated ischemic contracture in rat heart. Basic Res Cardiol 94:254–260
Pantos C, Malliopoulou V, Mourouzis I, Sfakianoudis K, Tzeis S, Doumba P, Xinaris C, Cokkinos AD, Carageorgiou H, Varonos DD, Cokkinos DV (2003) Propylthiouracil-induced hypothyroidism is associated with increased tolerance of the isolated rat heart to ischaemia-reperfusion. J Endocrinol 178:427–435
Patel AJ, Smith RM, Kingsbury AE, Hunt A, Balazs R (1980) Effects of thyroid state on brain development: muscarinic acetylcholine and GABA receptors. Brain Res 198:389–402
Rastogi RB, Hrdina PD, Dubas T, Singhal RL (1977) Alterations of brain acetylcholine metabolism during neonatal hyperthyroidism. Brain Res 123:188–192
Sanui H, Rubin H (1982) The role of magnesium in cell proliferation and transformation. In: Boynton AL, McKochan WL, Whitfield JP (eds). Ions cell proliferation and cancer. Academic Press, New York, pp. 517–537
Sastry BS, Phillis JW (1977) Antagonism of biogenic amine-induced depression of cerebral cortical neurones by Na+, K+-ATPase inhibitors. Can J Physiol Pharmacol 55:170–180
Swann AC (1984) (Na+, K+)-adenosine triphosphatase regulation by the sympathetic nervous system: Effects of noradrenergic stimulation and lesion in vivo. J Pharmacol Exp Ther 228:304–311
Tsakiris S (2001) Effects of L-phenylalanine on acetylcholinesterase and Na+, K+-ATPase activities in adult and aged rat brain. Mech Ageing Dev 122:491–501
Tsakiris S, Angelogianni P, Schulpis KH, Behrakis P (2000) Protective effect of L-cysteine and glutathione on rat brain Na+, K+-ATPase inhibition induced by free radicals. Z Naturforsch [C] 55:271–277
Virgili M, Saverino O, Vaccari M, Barnabei O, Contestabile A (1991) Temporal, regional and cellular selectivity of neonatal alteration of the thyroid state on neurochemical maturation in the rat. Exp Brain Res 83:555–561
Acknowledgements
This work was funded by the University of Athens and the University of Ioannina. Many thanks are expressed to the medical students Elena Gkrouzman, Zois Mellios and Christina Katsioni for their assistance.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Carageorgiou, H., Pantos, C., Zarros, A. et al. Effects of hyper- and hypothyroidism on acetylcholinesterase, (Na+, K+)- and Mg 2+-ATPase activities of adult rat hypothalamus and cerebellum. Metab Brain Dis 22, 31–38 (2007). https://doi.org/10.1007/s11011-006-9034-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11011-006-9034-1