
Abstract
The present study was undertaken to evaluate the protective effects of genistein against cardiac inflammation and oxidative stress in streptozotocin (STZ) (45 mg/kg body weight)-induced diabetic rats. genistein (300 mg/kg/day) was administered orally for 24 weeks to STZ-induced diabetic rats. The effects of genistein on blood glucose, % glycosylated hemoglobin (HbA1c), C-reactive protein, tumor necrosis factor (TNF- α), transforming growth factor (TGF-β1), and total antioxidant were studied. Ultrastructural and histopathological assessment of injury were also undertaken using transmission electron microscope. STZ-induced diabetes resulted in significant increase in the levels of blood glucose, HbA1c, C-reactive protein, TNF- α and TGF-β1, and a decline in total antioxidant reserve of the myocardium. Administration of genistein to diabetic rats resulted in a decrease in blood glucose (p < 0.001), % HbA1c (p < 0.0001), C-reactive protein (p < 0.001), and expression of TNF- α (p < 0.001) and TGF-β1 (p < 0.0001) proteins. In addition, genistein treatment results in augmentation of total antioxidant (p < 0.01) reserve of the hearts. The above findings were supported by histological as well as immunohistochemical localization of NF-κB (p65) in the heart. Genistein treatment ameliorated the ultrastructural degenerative changes in the cardiac tissues as compared to the diabetic control. The result demonstrates that genistein restored the integrity of the diabetic myocardium by virtue of its anti-inflammatory and antioxidant effects.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Diabetic cardiomyopathy (DCM) is a leading cause of cardiovascular complications, characterized by pathological diastolic as well as systolic dysfunction, ventricular dilation, cardiomyocyte hypertrophy, and prominent interstitial fibrosis [1–3]. A number of clinical studies and animal models have established that hyperglycemia is the leading cause for the progression of diabetic cardiovascular complications. An elevated glucose levels induces structural and functional changes in the cardiomyocytes through the activation of several signal transduction pathways, including oxidative stress, cardiac inflammation, and fibrosis [4–8].
The increased levels of oxidative stress caused by imbalance between the generation of reactive oxygen species (ROS) and endogenous reserve of antioxidants, result in initiation and progression of cardiac dysfunction, remodeling of the extracellular matrix in the heart, and culminating in oxidative myocardial injury [9, 10].
Chronic inflammation directly and indirectly results in myocardial fibrosis, necrosis, and apoptosis [8]. This is supported by increased expression of TNF-α, a pro-inflammatory cytokine in the myocardium. Excessive TNF-α level activates NF-κB and induces the RAGE gene resulting in left ventricular (LV) dysfunction and cardiomyopathy [11–14].
C-reactive protein (CRP), a non-specific inflammatory marker, is elevated in the serum of patients with type 1and type 2 diabetes and in a streptozotocin (STZ)-induced type 1 experimental diabetes model [15, 16]. Elevation of CRP is reported to be associated with LV dysfunction and is a predictor of mortality and morbidity in patients with established heart failure, independent of ischemic/non-ischemic etiology and other established cardiovascular risk factors [17, 18].
Transforming growth factor β1 (TGF-β1) is a prosclerotic cytokine that is consistently implicated in organ fibrosis and hypertrophy [19, 20]. TGF-β1 is over-expressed in hypertrophic myocardium during the transition from stable hypertrophy to heart failure [21] and up-regulation of TGF-β1 correlates with the degree of fibrosis in the pressure overloaded heart [21]. However, Bugyei-Twum et al. [22] have previously reported that in a clinically relevant animal model of diabetes-induced heart failure [the diabetic m(Ren2)27 rat], with preserved ejection fraction, increased interstitial fibrosis and cellular hypertrophy are mediated by upregulated TGF-β1 activity and Smad2 phosphorylation [22]. However, the molecular mechanisms and inter-linked pathways by which hyperglycemia mediate the increased TGF-β1 activity and downstream canonical Smad signaling needs to be elucidated.
Genistein (4′,5′,7′-trihydroxyisoflavone) is a well-characterized multifunctional isoflavone found in soy beans is a strong antioxidant and biological activities including tyrosine kinase inhibitor [23, 24]. Dietary genistein has been shown to suppress endotoxin-induced inflammatory reaction in liver and intestine [25]. Genistein reportedly protects against a number of human chronic diseases, including cancer and diabetes [26, 27]. It would therefore be interesting and possibly fruitful to evaluate the possible beneficial antioxidant and anti-inflammatory effects of genistein in the setting of diabetes-induced cardiomyopathy. Therefore, in the present study, the potential of genistein to ameliorate hyperglycemia, cardiac oxidative stress, fibrosis, and hypertrophy in STZ-induced experimental diabetic rats will be investigated. In addition, genistein ability to suppress the inflammatory cytokine pathway and its downstream signaling targets will also be determined.
Materials and methods
Streptozotocin was obtained from HiMedia India. Enzyme-linked immunosorbent assay (ELISA) kits were purchased from Diaclone, France and RayBiotech, Inc, USA. Total antioxidant capacity assay kit was procured from Cayman Chemical Company, USA. Glycosylated hemoglobin (HbA1c) kits from Biosystems S.A. Costa Brava, Spain. Rabbit polyclonal NF-κB (p65) antibody was purchased from Abcam, Plc., UK and detection system for immunohistochemistry was obtained from Bio SB., USA.
Streptozotocin-induced diabetes
The protocol was approved by the Institutional Animal Ethics Committee (IAEC/2012-I/Prot. No.-02). Wistar albino rats of either sex (200–250 g) obtained from the Animal facility of the Institute were housed under controlled room temperature (21 ± 2 °C) with a 12/12 h light/dark cycle. The experimental rats received food and water ad libitum. Diabetes was induced with single intraperitoneal injection of streptozotocin (STZ, 45 mg/kg body weight) in 0.1 M citrate buffer (pH 4.5). Normal rats were injected with the equal volume of citrate buffer. The rats showing a blood glucose concentration greater than 300 mg/dl after 48 h STZ injection were considered diabetic and included in the study for 24 weeks.
Experimental groups
Age-matched healthy rats served as normal control (NC). Diabetic rats were randomly divided into diabetic control (DC) and genistein group (GNST) (n = 16/group). DC received normal diet and GNST group received genistein (300 mg/kg/day) treatment was initiated when blood glucose concentration was found to be more than 300 mg/dl after STZ injection and continued up to 24 weeks by oral administration to direct stomach intubation in a constant volume of 0.5 ml/kg body weight. One unit of insulin was administered subcutaneously twice a week or as desired. The dose of genistein was chosen based on recent published studies showing that genistein at 300 mg/kg was useful as pharmacological approach for the treatment of diabetic-induced cardiomyopathy [28–30]. The body weight of the rats was measured weekly using sensitive digital balance.
Estimation of biochemical parameters
At the end of 24 weeks after induction of diabetes, the rats were euthanized using CO2 chamber. The hearts were isolated for biochemical estimations, histopathological, and ultrastructural (TEM) studies. After removing the heart, atria and large vessels were carefully trimmed and weighed. A 10 % heart homogenate was prepared in 50 Mm phosphate buffer (pH 7.4). The homogenate was centrifuged at 7000 rpm at 4 °C for 15 min and a supernatant was obtained [3]. From the supernatant, the biochemical parameters CRP, TGF-β, TNF-α, and total antioxidant capacity were estimated. Blood samples were also analyzed for glucose and HbA1C levels in different groups.
Glycemic parameters
Weekly blood glucose was measured using Accu-Chek® Active Glucose Test Strips and Accu-Chek® meter (Roche Diagnostics India Pvt. Ltd) and this device was calibrated to confirm the accuracy before every use. HbA1c was estimated at the end of 24 weeks (Biosystems S.A. Costa Brava 30, Barcelona, Spain).
Inflammatory parameters
TNF-α (Diaclone, France), CRP (Assaypro, St. Charles, MO), and TGF-β (Qayee-Bio, China) levels in heart homogenate were estimated using enzyme-linked immunosorbent assay (ELISA) kit as per the manufacturer’s instructions.
Total antioxidant capacity assay
The total antioxidant capacity of the heart homogenate (10 µl of sample) was measured by a colorimetric technique using a kit from Cayman Chemical (Ann Arbor, MI). The assay is based on the ability of the sample to inhibit oxidation of 2,2′-azino-di-[3-ethylbenzthiazoline sulfonate] + (ABTS) by metmyoglobin; the antioxidants in the sample cause the decrease in absorbance at 750 nm and that represents the amount of ABTS+ produced. Each sample was measured in duplicate.
Histopathological studies
Hearts were immersion fixed in 10 % phosphate buffered formalin. The heart was serially sliced into 3-mm sections perpendicular to the axis of the heart from apex to base. These sections were embedded in paraffin, sectioned at 5 µm, and serial sections were stained with hematoxylin–eosin (H and E). All histological parameters were quantified by an experienced pathologist blinded to the identity of the sample being examined.
Ultrastructural examination by transmission electron microscopy
Hearts were cut into approximately 1 mm3 pieces, fixed in 2.5 % glutaraldehyde in 0.1 mol/L sodium phosphate buffer (pH 7.4) for 6 h at 4 °C, and osmicated in 1 % osmium tetroxide for 1 h at 4 °C. The specimens were then washed with phosphate buffer, dehydrated with graded acetone, and then embedded in araldite CY212 to make tissue blocks. Semithin (1 μm) as well as ultrathin Sections (70–80 nm) were cut by ultramicrotome (Ultracut E, Reichert, Austria). The sections were stained with uranyl acetate and lead acetate and examined under a transmission electron microscope (Morgagni 268D, Fei Co., The Netherlands) operated at 60 kV by a morphologist blinded to the groups studied. At least four hearts from each group were examined for ultrastructural changes.
Immunohistochemistry
For Immunohistochemical studies, 4-μm formalin fixed, paraffin-embedded left ventricle tissue sections were taken on polylysine-coated slides. Sections were deparaffinised, rehydrated, and endogenous peroxidase activity was blocked with H2O2 in methanol. Sections were pre-treated in citrate buffer (pH 6.0) for antigen retrieval in a microwave [31]. Sections were incubated overnight under humid conditions using an NF-κB rabbit polyclonal antibody (dilution 1:1000, Abcam, Plc., UK) at 4 °C. Next slides were washed three times in Tris buffer for 5 min each and followed by the detection system (Bio SB., USA) used which was System-HRP with diaminobenzidine as a chromogen. Slides were counterstained with hematoxylin. The expression of cell immunopositivity was assessed under light microscope.
Statistical analysis
All data are expressed as mean ± standard deviation (SD). Statistical comparisons were made using one-way-analysis of variance (ANOVA) with post hoc analysis using Dunnet multiple comparison test (Graph Pad Prism, Version-5). Value of p < 0.05 was accepted as statistically significant.
Results
Effect of genistein on body weight changes
Before the study, there was no significant difference in baseline body weight of the animals in different groups (Fig. 1c). Diabetic rats without genistein indicated a significant decrease in body weight compared with NC (p < 0.0001). Treatment of genistein prevented body weight loss in rats as compared with DC groups rats (p < 0.001).

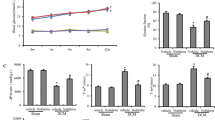
Genistein treatment for 24 weeks, a decreased blood glucose level. Values are mean ± SD; n = 6; *p < 0.001 vs diabetic, # p < 0.05, ## p < 0.01, ### p < 0.001 versus genistein-treated group. b Effects of genistein on glycated hemoglobin (HbA1c) c Increased body weight. Values are mean ± SD; n = 6; *p < 0.05, **p < 0.001 versus diabetic, # p < 0.05, ## p < 0.001 versus genistein-treated group
Other symptoms generally associated with the diabetic state, such as dull fur, hair loss, lack of grooming, inactivity, polydipsia, polyuria, and polyphagia, were also observed in the DC rats.
Effect of genistein on glycemic parameters
NC rats exhibited stable blood glucose levels (92.66 ± 6.08–115.67 ± 7.73 mg/dl) throughout the experimental period. Meanwhile, the fasting blood glucose levels of STZ-induced diabetic rats (457.33 ± 62.21–531.83 ± 35.02 mg/dl) were significantly higher than those of the NC rats, and this persistent hyperglycemia was accompanied by weight loss. The long-term treatment with genistein significantly (p < 0.001) prevented the rise in the blood glucose levels as compared with the DC rats (Fig. 1a).
In STZ-induced diabetic rats, genistein significantly (p < 0.0001) inhibited increase in HbA1c level after chronic daily drug administration when compared with diabetic group (Fig. 1b).
Effects of genistein on cytokines associated with diabetic cardiomyopathy
Figure 2a, b demonstrates that DC rats had significantly higher levels of TNF-α and CRP as compared to NC (p < 0.001). Genistein treatment was able to significantly decrease TNF-alpha and CRP in diabetic rats relative to DC (p < 0.001). The levels of TGF-β was significantly higher in diabetic rats as compared with NC (p < 0.001; Fig. 2c). Subsequent to STZ treated, genistein treatment significantly restored the levels of CRP (p < 0.0001).

a Decreased TNF-α, b restored CRP level c Pretreatment with genistein reduced TGF-β1 as compared to diabetic animals d total antioxidant capacity (TAC) in cardiac tissue was detected and expressed as micromoles, Values are mean ± SD; n = 6; *p < 0.01, ***p < 0.0001 versus diabetic, # p < 0.01, ## p < 0.001, ### p < 0.0001 versus genistein-treated group
Effects of genistein on total antioxidant capacity
The activity of total antioxidant capacity in heart homogenate of the diabetic group rats was significantly lower (p < 0.01) than that of the NC rats (Fig. 2d). Rats treated with genistein significant restored the antioxidant capacity of the myocardium (p < 0.01).
Effect of genistein on histopathological changes
Figure 3 shows the light micrograph of the normal architecture of the myocardium (without any fraying or infarction) in the NC group rats. Light micrograph of DC heart shows focal confluent necrosis of muscle fibers with inflammatory cell infiltration, edema, and myophagocytosis along with extravasation of red blood cells (Fig. 3b, c). The degree of myocardial damage in GNST-treated rats was significantly less than the DC rats. Mild edema with significant reduction in the myocardial necrosis was observed in the treatment group as compared to DC (Fig. 3d).

Histopathological observation of heart tissue: a normal cardiac myocytes with regular cross-striations, b diabetic group show focal mononuclear inflammation of myofibrils due to increased inflammatory stress (arrow), c diabetic heart showing focal area of infarction with necrosis of myocytes cells (arrow), and d genistein-treated rats heart showing a reversal of inflammation, focal infarction and moderate necrosis changes (n = 4; h, e; ×400). b Immunohistochemical staining for NF-κB (p65) in the left ventricle 24 weeks after disease induction. Sections were reacted with antibody specific for NF-κB (p65), color-developed with diaminobenzidine, and counterstained with hematoxylin. No expression was seen in NC (e). DC heart showing increased expression of NF-κB (p65) (f, g). In GNST-treated heart shows relatively lesser expression of NF-κB (p65) (h) (n = 4; ×400)
Effects of genistein on ultrastructural changes in the rat myocardium
TEM analysis showed the normal cardiomyocyte architecture in the hearts of the NC group. Regular-shaped cylinders, comprising of Z lines with diads, sarcomeres, elliptical nucleus, numerous mitochondria, and prominent myofilaments, were observed (Fig. 4a). In cardiomyocytes of untreated diabetic rats, edema was evident, and sarcomeres and myofilament arrangements were disordered and partially separated; meanwhile, the mitochondria were enlarged and rounded, with the cristae appearing disordered or disrupted (Fig. 4b, c). However, the damage to myocardial ultrastructure in GNST-treated rats was markedly attenuated as compared with that of the untreated diabetic group (Fig. 4d).

Effects of long-term treatment with genistein on the ultrastructure of rat hearts. The ultrastructure of heart tissues was detected by TEM from the left ventricle of the a NC group, b, c untreated diabetic group, and d genistein-treated diabetic group. Several mitochondria (M) exhibited swelling, fusion and disruption of cristae, and thinning of the Z lines were observed in the untreated diabetic group. Long-term treatment with genistein significantly improved these morphological changes
Effects of genistein on NF-κB
Immunohistochemical analysis of hearts from NC animals did not show localization (staining) for NF-κB in cardiac myocytes (Fig. 3e). In DC animals, NF-κB expression was markedly increased in the LV myocardium as compared with NC animals as determined by positive immunoreactivity for NF-κB (Fig. 3f, g). Comparatively lesser expression of NF-κB in cardiac myocytes was recorded in GNST-treated rats (Fig. 3h).
Discussion
Cardiomyopathy is one of the most prevalent cardiovascular complications of diabetes mellitus. Prominent defects of DCM include the prolonged duration of contraction and relaxation and reduced cardiac compliance [21]. Through compilation of several studies, various etiological causes for DCM such as hyperglycemia resulting in activation of oxidative stress and inflammatory cascade pathways have been identified [4, 5]. It is therefore plausible and logical, to identify pharmacological agents having antidiabetic activity with additional antioxidant and inflammatory properties for the management of diabetic cardiomyopathy. Such pharmacological agents if used as adjuvant to modern medicine may modulate the signal transduction pathways involved in inflammation as well as oxidative stress and attenuate the myocardial injury. With this point of view, the study was planned.
The cardioprotective effect of GNST on diabetes-induced myocardial injury was studied. The molecular basis for protection afforded by genistein was also studied. It has been reported that the soya phytoestrogen GNST reduces hyperglycemia and islet cell loss in alloxan-induced diabetic male Sprague–Dawley rats [32] and in STZ-induced diabetic male C57BL/6J mice [33, 34]. In addition, supplementation of GNST increased the plasma insulin level and decreased the HbA1c level in STZ-induced diabetic male Sprague–Dawley rats [35]. Guo et al., study also demonstrated that oral administration of GNST reduced the incidence as well as increased the time to onset of diabetes mellitus in genetically susceptible female mice when fed a soy- and alfalfa-free 5K96 diet [36]. The hypoglycemic effects of GNST as observed in the present study concur with previously published reports [32–37].
In the present study, body weight of DC rats was significantly decreased as compared to NC group rats. It may be hypothesized that the metabolic disturbances that occur in diabetes mellitus results in calorie deprivation which in turn leads to decrease in protein synthesis and promotes protein degradation in longer-lasting disease state [38]. However, treatment with genistein showed improvement in the body weight, which was statistically significant.
Inflammation characterized by elevated inflammatory cytokines production exists not only in the myocardium of animal models [39–41], but also in the serum of diabetic patients, even in those patients with good glycemic control [8, 42]. NF-κB is a pleiotropic transcription factor that controls the expression of several target genes, involved mainly in inflammation. Under hyperglycemic condition, the DNA binding activity of NF-κB p65 is enhanced, and its mRNA and protein expressions are also upregulated. The NF-κB activation led to the increased expression of inflammatory mediators, including pro-inflammatory cytokine like TNF-α and CRP. These inflammatory cytokines can attenuate cardiomyocyte contractility either directly through the immediate reduction of systolic cytosolic calcium and indirectly through attenuation of myofilament calcium sensitivity [43]. This detrimental effect may be reversible by clearance of the cytokine exposure. Similarly, diabetic rats in the present study showed significantly increased level of TNF-α, CRP, and expression of NF-κB in diabetic heart as compared to NC. However, GNST demonstrated significant anti-inflammatory activity as it prevented the increase in levels of TNF- α, CRP, and expression of NF-κB, [37, 44, 45].
Hyperglycemia can modulate both the metabolic and hemodynamic pathways to change growth factors [20]. The important role of growth factors in pathogenesis of long-term complications of diabetes was suggested by their increased concentrations in target tissue [9]. Ample evidence demonstrates elevated TGF-β1 expression during the transition from stable hypertrophy to heart failure in both experimental models and human heart failure [21, 46]. Canonical TGF-β1 signaling involves the receptor-activated Smad proteins (Smad2 and Smad3), which, upon phosphorylation, associate with Smad4, translocate to the nucleus and act as transcription factors [22, 46]. Similarly, diabetic rats in the present study showed significantly increased level of TGF-β1 in the myocardium as compared to NC. Interestingly GNST reduced the expression of TGF-β1 and prevented its deleterious effects [47, 48].
Several studies demonstrate that oxidative stress is associated with diabetic complications in both humans and animals [9, 10]. Oxidative stress is an imbalance between oxidants, such as ROS and antioxidants, and probably contributes to the development, progression, and complications of diabetes [10]. However, the mechanism underlying the development of structural and functional impairment in cardiomyopathy is still unknown, and the signal pathway that leads to the pathological changes of cardiomyocyte hypertrophy is still unclear. Oxidative stress often causes cell death via apoptosis and necrosis. A pathogenic role for oxidative stress in DCM is becoming increasingly apparent [9, 49–53]. Our results demonstrate that the oxidative stress resulting from hyperglycemia is a potential mechanism for diabetic cardiac complications. In this present study, it was observed that diabetes caused a significant decrease in TAC level, which indicates abnormal LV systolic and diastolic function. Treatment with genistein significantly increased myocardial TAC levels and augmented the endogenous reserve of the myocardium. Therefore under stressful conditions, the GNST-treated rats would be better prepared to with stand any such stressors such as hyperglycemia, as compared to the DC rats. This result confirms the antioxidant effect of genistein published by different authors [27, 37, 47, 48].
Diabetic cardiomyopathy is characterized by both systolic and diastolic dysfunction and altered cardiac structure [3, 54]. In the current study, both histological changes as well as ultrastructural changes of the myocardium were evaluated. Histopathological findings of the heart pre-treated with genistein showed a well preserved normal morphology of cardiac muscle with no significant evidence of necrosis, edema, and inflammation when compared to STZ-induced myocardial infracted heart and further reconfirmed the beneficial effects of genistein. Genistein treatment afforded significant cardioprotection as observed histopathologically and as determined by TEM. Genistein-treated protected against ultrastructural features of cell damage, including disorganization of myofibrils and aggregation of large mitochondria with swollen and broken cristae, in hearts of the 24-week-untreated diabetic rats. In conclusion, our present work confirmed that TNF-α, CRP, and TGF-β1 are involved in DCM and that genistein treatment ameliorated the progression of cardiac remodeling (fibrosis and hypertrophy) in experimental diabetes. The histopathological studies confirmed the cardioprotective effects of genistein. Cardioprotection seems to be mediated, at least in part, by inhibiting the activities of TNF-α, CRP, and TGF-β1. Moreover, genistein seemingly contributed to restoring the antioxidant capacity of the myocardium. Taken together, our results implicate the therapeutic potential of genistein as adjuvant therapy for treating early diabetic cardiomyopathy.
References
Dhalla NS, Liu X, Panagia V, Takeda N (1998) Subcellular remodeling and heart dysfunction in chronic diabetes. Cardiovasc Res 40:239–247
Mizushige K, Yao L, Noma T, Kiyomoto H, Yu Y, Hosomi N, Ohmori K, Matsuo H (2000) Alteration in left ventricular diastolic filling and accumulation of myocardial collagen at insulin-resistant prediabetic stage of a type II diabetic rat model. Circulation 101:899–907
Guan SJ, Ma ZH, Wu YL, Zhang JP, Liang F, Weiss JW, Guo QY, Wang JY, Ji ES, Chu L (2012) Long-term administration of fasudil improves cardiomyopathy in streptozotocin-induced diabetic rats. Food Chem Toxicol 50:1874–1882
Falcao-Pires I, Leite-Moreira AF (2012) Diabetic cardiomyopathy: understanding the molecular and cellular basis to progress in diagnosis and treatment. Heart Fail Rev 17:325–344
Hotamisligil GS (2006) Inflammation and metabolic disorders. Nature 444:860–867
Stratmann B, Tschoepe D (2011) The diabetic heart: sweet, fatty and stressed. Expert Rev Cardiovasc Ther 9:1093–1096
Ku PM, Chen LJ, Liang JR, Cheng KC, Li YX, Cheng JT (2011) Molecular role of GATA binding protein 4 (GATA-4) in hyperglycemia-induced reduction of cardiac contractility. Cardiovasc Diabetol 10:1–15
Wen HL, Liang ZS, Zhang R, Yang K (2013) Anti-inflammatory effects of triptolide improve left ventricular function in a rat model of diabetic cardiomyopathy. Cardiovasc Diabetol 12:1–11
Wang G, Li W, Lu X, Bao P, Zhao X (2012) Luteolin ameliorates cardiac failure in type I diabetic cardiomyopathy. J Diabetes Complicat 4:259–265
Rösen P, Nawroth PP, King G, Möller W, Tritschler HJ, Packer L (2001) The role of oxidative stress in the onset and progression of diabetes and its complications: a summary of a Congress Series sponsored by UNESCO-MCBN, the American Diabetes Association and the German Diabetes Society. Diabetes Metab Res Rev 17:189–212
Westermann D, Van Linthout S, Dhayat S, Dhayat N, Schmidt A, Noutsias M, Song XY, Spillmann F, Riad A, Schultheiss HP, Tschöpe C (2007) Tumor necrosis factor-alpha antagonism protects from myocardial inflammation and fibrosis in experimental diabetic cardiomyopathy. Basic Res Cardiol 6:500–507
Sara N, Edna S, Frederico P, Flávio R (2012) The role of inflammation in diabetic cardiomyopathy. Int J Infereron Cytokine Mediator Res 4:59–73
Esposito K, Nappo F, Marfella R, Giugliano G, Giugliano F, Ciotola M, Quagliaro L, Ceriello A, Giugliano D (2002) Inflammatory cytokine concentrations are acutely increased by hyperglycemia in humans: role of oxidative stress. Circulation 106:2067–2072
Patel S, Santani D (2009) Role of NF-kappa B in the pathogenesis of diabetes and its associated complications. Pharmacol Rep 4:595–603
Rutter MK, Meigs JB, Sullivan LM, D’Agostino RB Sr, Wilson PW (2004) C-reactive protein, the metabolic syndrome, and prediction of cardiovascular events in the Framingham Offspring Study. Circulation 110:380–385
Goyal BR, Mesariya P, Goyal RK, Mehta AA (2008) Effect of telmisartan on cardiovascular complications associated with streptozotocin diabetic rats. Mol Cell Biochem 314:123–131
Shah SJ, Marcus GM, Gerber IL, McKeown BH, Vessey JC, Jordan MV, Huddleston M, Foster E, Chatterjee K, Michaels AD (2006) High-sensitivity C-reactive protein and parameters of left ventricular dysfunction. J Card Fail 12:61–65
Kardys I, Knetsch AM, Bleumink GS, Deckers JW, Hofman A, Stricker BH, Witteman JC (2006) C-reactive protein and risk of heart failure: the Rotterdam Study. Am Heart J 152:514–520
Border WA, Noble NA (1995) Fibrosis linked to TGF-beta in yet another disease. J Clin Invest 2:655–656
Khan R, Sheppard R (2006) Fibrosis in heart disease: understanding the role of transforming growth factor-beta in cardiomyopathy, valvular disease and arrhythmia. Immunology 1:10–24
Hein S, Arnon E, Kostin S, Schonburg M, Elsasser A, Polyakova V, Bauer EP, Klovekorn WP, Schaper J (2003) Progression from compensated hypertrophy to failure in the pressure-overloaded human heart: structural deterioration and compensatory mechanisms. Circulation 7:984–991
Bugyei-Twum A, Advani A, Advani SL, Zhang Y, Thai K, Kelly DJ, Connelly KA (2014) High glucose induces Smad activation via the transcriptional coregulator p300 and contributes to cardiac fibrosis and hypertrophy. Cardiovasc Diabetol 13(89):1–12
Naderi GA, Asgary S, Sarraf-Zadegan N (2003) Anti-oxidant effect of flavonoids on the susceptibility of LDL oxidation. Mol Cell Biochem 246:193–196
Akiyama T, Ishida J, Nakagawa S, Ogawara H, Watanabe S, Itoh N (1987) Genistein, a specific inhibitor of tyrosine-specific protein kinases. J Biol Chem 12:5592–5595
Paradkar PN, Blum PS, Berhow MA, Baumann H, Kuo SM (2004) Dietary isoflavones suppress endotoxin-induced inflammatory reaction in liver and intestine. Cancer Lett 215:21–28
Park CE, Yun H, Lee EB, Min BI, Bae H, Choe W, Kang I, Kim SS, Ha J (2010) The antioxidant effects of genistein are associated with AMP-activated protein kinase activation and PTEN induction in prostate cancer cells. J Med Food 13:815–820
Gilbert ER, Liu D (2013) Anti-diabetic functions of soy isoflavone genistein: mechanisms underlying effects on pancreatic β-cell function. Food Funct 2:200–212
Nakajima M, Cooney MJ, Tu AH, Chang KY, Cao J, Ando A, An GJ, Melia M, de Juan E Jr (2001) Normalization of retinal vascular permeability in experimental diabetes with genistein. Invest Ophthalmol Vis Sci 42:2110–2114
Ramos JE, Al-Nakkash L, Peterson A, Gump BS, Janjulia T, Moore MS, Broderick TL, Carroll CC (2012) The soy isoflavone genistein inhibits the reduction in Achilles tendon collagen content induced by ovariectomy in rats. Scand J Med Sci Sports 5:e108–e114
Zeng X, Feng Y, Yang L, Huang Y, Zhou D, Sun J, Liu Y, Deng Y (2008) Single- and multiple-dose pharmacokinetics of genistein capsules in healthy chinese subjects: a phase I, randomized, open-label study. Curr Ther Res Clin Exp 4:318–333
Fouad AA, Al-Sultan AI, Yacoubi MT, Gomaa W (2010) Ameliorative effects of telmisartan in diabetic rats with indomethacin-induced gastric ulceration. Eur J Pharmacol 637:162–170
Yang W, Wang S, Li L, Liang Z, Wang L (2011) Genistein reduces hyperglycemia and islet cell loss in a high-dosage manner in rats with alloxan-induced pancreatic damage. Pancreas 40:396–402
Fu Z, Zhang W, Zhen W, Lum H, Nadler J, Bassaganya-Riera J, Jia Z, Wang Y, Misra H, Liu D (2010) Genistein induces pancreatic beta-cell proliferation through activation of multiple signaling pathways and prevents insulin-deficient diabetes in mice. Endocrinology 151:3026–3037
Elmarakby AA, Ibrahim AS, Faulkner J, Mozaffari MS, Liou GI, Abdelsayed R (2011) Tyrosine kinase inhibitor, genistein, reduces renal inflammation and injury in streptozotocin-induced diabetic mice. Vasc Pharmacol 55:149–156
Lee JS (2006) Effects of soy protein and genistein on blood glucose, antioxidant enzyme activities, and lipid profile in streptozotocin-induced diabetic rats. Life Sci 79:1578–1584
Guo TL, Germolec DR, Zheng JF, Kooistra L, Auttachoat W, Smith MJ, White KL, Elmore SA (2014) Genistein protects female nonobese diabetic mice from developing type 1 diabetes when fed a soy- and alfalfa-free diet. Toxicol Pathol (Apr 8. [Epub ahead of print] PubMed PMID: 24713318)
Behloul N, Wu G (2013) Genistein: a promising therapeutic agent for obesity and diabetes treatment. Eur J Pharmacol 698:31–38
Nemoto O, Kawaguchi M, Yaoita H, Miyake K, Maehara K, Maruyama Y (2006) Left ventricular dysfunction and remodeling in streptozotocin-induced diabetic rats. Circ J 70:327–334
Van Linthout S, Spillmann F, Riad A, Trimpert C, Lievens J, Meloni M, Escher F, Filenberg E, Demir O, Li J, Shakibaei M, Schimke I, Staudt A, Felix SB, Schultheiss HP, De Geest B, Tschope C (2008) Human apolipoprotein A-I gene transfer reduces the development of experimental diabetic cardiomyopathy. Circulation 117:1563–1573
Sun D, Shen M, Li J, Li W, Zhang Y, Zhao L, Zhang Z, Yuan Y, Wang H, Cao F (2011) Cardioprotective effects of tanshinone IIA pretreatment via kinin B2 receptor-Akt-GSK-3b dependent pathway in experimental diabetic cardiomyopathy. Cardiovasc Diabetol 10:1–8
Rajesh M, Mukhopadhyay P, Batkai S, Patel V, Saito K, Matsumoto S, Kashiwaya Y, Horvath B, Mukhopadhyay B, Becker L, Hasko G, Liaudet L, Wink DA, Veves A, Mechoulam R, Pacher P (2010) Cannabidiol attenuates cardiac dysfunction, oxidative stress, fibrosis, and inflammatory and cell death signaling pathways in diabetic cardiomyopathy. J Am Coll Cardiol 56:2115–2125
Snell-Bergeon JK, West NA, Mayer-Davis EJ, Liese AD, Marcovina SM, D’Agostino RB Jr, Hamman RF, Dabelea D (2010) Inflammatory markers are increased in youth with type 1 diabetes: the SEARCH Case–control study. J Clin Endocrinol Metab 95:2868–2876
Nian M, Lee P, Khaper N, Liu P (2004) Inflammatory cytokines and postmyocardial infarction remodeling. Circ Res 94:1543–1553
Ji G, Yang Q, Hao J, Guo L, Chen X, Hu J, Leng L, Jiang Z (2011) Anti-inflammatory effect of genistein on non-alcoholic steatohepatitis rats induced by high fat diet and its potential mechanisms. Int Immunopharmacol 6:762–768
Deodato B, Altavilla D, Squadrito G, Campo GM, Arlotta M, Minutoli L, Saitta A, Cucinotta D, Calapai G, Caputi AP, Miano M, Squadrito F (1999) Cardioprotection by the phytoestrogen genistein in experimental myocardial ischaemia-reperfusion injury. Br J Pharmacol 8:1683–1690
Hayashi H, Abdollah S, Qiu Y, Cai J, Xu YY, Grinnell BW, Richardson MA, Topper JN, Gimbrone MA Jr, Wrana JL, Falb D (1997) The MAD-related protein Smad7 associates with the TGFbeta receptor and functions as an antagonist of TGF beta signaling. Cell 7:1165–1173
Kim YS, Kim NH, Jung DH, Jang DS, Lee YM, Kim JM, Kim JS (2008) Genistein inhibits aldose reductase activity and high glucose-induced TGF-beta2 expression in human lens epithelial cells. Eur J Pharmacol 594:18–25
Kim H, Peterson TG, Barnes S (1998) Mechanisms of action of the soy isoflavone genistein: emerging role for its effects via transforming growth factor β signaling pathways1−3. Am J Clin Nutr 68:1418S–1425S
Agarwal A, Nick HS (2000) Renal response to tissue injury: lessons from heme oxygenase-1 gene ablation and expression. J Am Soc Nephrol 5:965–973
Ryter SW, Kim HP, Nakahira K, Zuckerbraun BS, Morse D, Choi AM (2007) Protective functions of heme oxygenase-1 and carbon monoxide in the respiratory system. Antioxid Redox Signal 9:2157–2173
Grieve DJ, Byme JA, Cave AC, Shah AM (2004) Role of oxidative stress in cardiac remodeling after myocardial infarction. Heart Lung Circ 13:132–138
Kameda K, Matsunaga T, Abe N, Hanada H, Ishizaka H, Ono H, Saitoh M, Fukui K, Fukuda I, Osanai T, Okumura K (2003) Correlation of oxidative stress with activity of matrix metalloproteinase in patients with coronary artery disease: possible role for left ventricular remodeling. Eur Heart J 24:2180–2185
Molavi B, Mehta JL (2004) Oxidative stress in cardiovascular disease: molecular basis of its deleterious effects, its detection, and therapeutic considerations. Curr Opin Cardiol 19:488–493
Boudina S, Abel ED (2007) Diabetic cardiomyopathy revisited. Circulation 115:3213–3223
Acknowledgments
Financial support provided by UKIERI and Department of Science and Technology (DST/INT/UK/P-39/2012), India is gratefully acknowledged. Facilities for electron microscopy availed at SAIF (DST), All India Institute of Medical Sciences, New Delhi, are acknowledged.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Gupta, S.K., Dongare, S., Mathur, R. et al. Genistein ameliorates cardiac inflammation and oxidative stress in streptozotocin-induced diabetic cardiomyopathy in rats. Mol Cell Biochem 408, 63–72 (2015). https://doi.org/10.1007/s11010-015-2483-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11010-015-2483-2