
Abstract
Context
Burrowing mammals play a role in rangeland disturbance worldwide, enhancing habitat for certain species while negatively affecting others. However, little is known concerning effects of disturbance spatial pattern on co-occuring fauna. In the North American Great Plains, colonial black-tailed prairie dogs (Cynomys ludovicianus) may enhance habitat for one suite of birds while degrading habitat for others.
Objectives
We examined the influence of prairie dogs on birds in a mosaic grassland–shrubland landscape. We evaluated how birds associated with shortgrass, midgrass, and sagebrush (Artemisia spp.) plant communities respond to spatial pattern of prairie dog disturbance and identified thresholds where abundance changes.
Methods
We surveyed bird abundance on prairie dog colonies of varying sizes and shapes, across colony edges into undisturbed habitat, and within undisturbed sagebrush in northeastern Wyoming. We modeled species responses to colony presence, distance to colony edge, and total area and edge density of colonies at four spatial scales (100 m, 225 m, 500 m, 1000 m).
Results
Sagebrush specialists like Brewer’s sparrow (Spizella breweri) and sage thrasher (Oreoscoptes montanus) were 4.5 times more abundant in undisturbed shrublands. Conversely, the shortgrass-specialist mountain plover (Charadrius montanus) was abundant on colonies but showed a non-linear response to colony edge, increasing in abundance up to 600 m from edges then declining further towards colony cores.
Conclusions
While some species may be broadly intolerant to disturbance, disturbance-dependent birds can display a “goldilocks syndrome” relative to disturbance size. As such, management for multiple species of conservation concern can be optimized relative to other goals by identifying thresholds associated with the effect of disturbance.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Rangeland ecosystems worldwide are shaped by disturbance processes including fire and grazing by mammalian herbivores (Davidson and Lightfoot 2007; Fuhlendorf et al. 2012). North American rangelands were drastically altered following European settlement as a result of conversion to rowcrops (Wright and Wimberly 2013), energy extraction (Allred et al. 2015), exurban development (Riebsame et al. 1996; Sala et al. 2017), introduction of exotic species (USDA 2010; DiTomaso et al. 2017), extirpation and control of undesirable native species (Miller et al. 2007; Sayre 2017), and alteration of historic disturbance regimes (Sayre 2017; Fuhlendorf et al. 2017). These changes have severely reduced habitat availability and quality for rangeland wildlife, making these species a key target for conservation. Restoration of historic disturbance regimes is especially important for rangeland wildlife that evolved in the context of these disturbances (e.g., Brawn et al. 2001; Fuhlendorf and Engle 2001). Throughout the Great Plains, disturbance caused by black-tailed prairie dogs (Cynomys ludovicianus), which are colonial, burrowing, herbivorous mammals, has been identified as a major driver of community structure in grasslands (Lomolino and Smith 2004; Augustine and Baker 2013). This oversized influence has led black-tailed prairie dogs to be labeled as both a keystone species (Kotliar et al. 1999) and an ecosystem engineer (Van Nimwegen et al. 2008). While the black-footed ferret (Mustela nigripes) is well known as a globally-imperiled species that is an obligate predator of prairie dogs (Dobson and Lyles 2000), many other species including herptiles (Shipley and Reading 2006), arthropods (Davidson and Lightfoot 2007), mammals (Lomolino and Smith 2004; Shipley and Reading 2006), and birds (Augustine and Baker 2013) rely on colonies for breeding habitat and food resources.
Despite the crucial role black-tailed prairie dogs play in these systems, they occupy less than 1% of their historic range (Miller and Cully 2001) and continue to experience lethal control because of potential competition with livestock (Derner et al. 2006; Miller et al. 2007). Prairie dog populations are further reduced by epizootics of sylvatic plague (Yersina pestis), which can lead to > 95% dieoff within colony complexes (Cully et al. 2010). Prairie dog declines due to lethal control and plague epizootics have been directly linked with declines in imperiled grassland birds including the mountain plover (Charadrius montanus; Augustine et al. 2008; Dinsmore and Smith 2010) and burrowing owl (Athene cunicularia; Desmond et al. 2000).
Restoration of black-tailed prairie dogs has benefitted many species but may negatively affect disturbance-intolerant birds. The black-tailed prairie dog’s propensity for pervasive vegetation clipping and burrow construction can be a detriment to species that rely on tall, dense grasses (Augustine and Derner 2015) or shrubs (Knick et al. 2005; Beck et al. 2012) for nesting and foraging. Specifically, sagebrush-associated birds may be affected if frequent prairie dog clipping and girdling kills sagebrush (Johnson-Nistler et al. 2004; Ponce-Guevara et al. 2016), which is slow to recover following disturbance (Baker 2006; Porensky et al. 2018). Much of the sagebrush steppe lacks prairie dogs entirely or is inhabited by white-tailed (Cynomys leucurus) or Gunnison’s (Cynomys gunnisoni) prairie dogs, which are less inclined to clip vegetation, live at far lower densities, and have smaller colony sizes than black-tailed prairie dogs (Hoogland 1995). However, the eastern portion of sagebrush distribution, where these shrubs coexist with grasses characteristic of the mixed-grass prairie (Porensky et al. 2018), overlaps with the distribution of black-tailed prairie dogs (Fig. 1). At this ecotone between the Great Plains and the sagebrush steppe, black-tailed prairie dogs engineer habitats that may benefit certain grassland species, but negatively impact sagebrush-associated birds.

Location of the TBNG within eastern Wyoming, USA, where the range of the black-tailed prairie dog overlaps the sagebrush steppe. Inset: Avian point count locations (n = 439) within sagebrush and across long-term and new prairie dog colony habitat in the Thunder Basin, surveyed between 2015 and 2017
Although previous research indicates that prairie dog disturbance affects sagebrush and shortgrass bird species differently (Augustine and Baker 2013), we do not yet understand at what scales these species respond, and whether there are thresholds beyond which habitat becomes unsuitable. Identification of these thresholds is of crucial import in rangelands where multiple bird guilds coexist with livestock grazing, because they imply that small changes to management or disturbance regimes in such landscapes may have large impacts on avian diversity.
We sought to address these issues by examining the influence of black-tailed prairie dog disturbance on priority bird species on the Thunder Basin National Grassland (TBNG). This mosaic grassland-sagebrush landscape is managed by the U.S. Forest Service, representing a considerable opportunity for the management and conservation of declining grassland and sagebrush birds. However, because National Grasslands are managed as multiple-use landscapes, understanding the role of fine-scale changes in disturbance amount and configuration is important for sustaining all uses, including livestock grazing and habitat for wildlife. The TBNG encompasses one of the largest contiguous tracts of public land managed for both wildlife conservation and livestock production in central North America, and the size and distribution of black-tailed prairie dog colonies has been monitored for the past two decades (Cully et al. 2010), creating a unique opportunity to examine how such variation influences the abundance of rangeland bird species.
We explored the relationship between disturbance and rangeland birds with three objectives in mind. First, we asked how both vegetation structure and composition changed as a function of spatial and temporal variation in the history of prairie dog disturbance, in particular evaluating whether these changes varied with distance to disturbed patch edge or were more marked in areas of long-term disturbance (Q1). Second, we examined the direct response of rangeland birds to prairie dog disturbance (Q2). Specifically, we were interested in the responses of shortgrass (‘disturbance-dependent’) species versus responses of species relying on less disturbed habitat (midgrass and sagebrush birds). Finally, we wanted to understand how these same species responded to both the composition and configuration of prairie dog disturbance at multiple spatial scales (Q3). We predicted that vegetation structure would differ substantially between disturbed and undisturbed patches, and as a result that bird communities would differ on and off colonies; however, it was unclear how each species would respond to non-binary features of disturbance, like distance to colony edge, or percent colony cover at various spatial scales.
Methods
Study area
Our study was conducted on public lands within the U.S. Forest Service (USFS)–Thunder Basin National Grassland in Converse, Weston, and Campbell counties, Wyoming, USA (Fig. 1). Mean annual precipitation ranges from 25 to 35 cm (Porensky et al. 2018) mainly falling as rain in the spring and summer. During our study (2015–2017) annual precipitation was 34 cm, 20 cm and 32 cm respectively (NOAA 2018). Summer high temperatures average around 27 °C (80 °F), but can exceed 38°.
Common graminoids included blue grama (Bouteloua gracilis), western wheatgrass (Pascopyrum smithii), needle-and-thread (Heterostipa comata), and threadleaf sedge (Carex filifolia). Wyoming big sagebrush (Artemisia tridentata wyomingensis) occurred in lower densities in this region than elsewhere in the sagebrush steppe (Chambers et al. 2016), but reached > 30% canopy cover in some areas (Porensky et al. 2018). Other common species included greasewood (Sarcobatus vermiculatus), broom snakeweed (Gutierrezia sarothrae), plains pricklypear (Opuntia polyacantha), and other sagebrush species (Artemisia spp.).
The entirety of our study area in the southern portion of the Thunder Basin represents over 1000 km2 (100,000 ha) of sagebrush, grassland, and prairie dog colonies, as well as some badlands and riparian areas. Black-tailed prairie dog colonies expanded over the duration of the study, with mean colony size increasing from 2015 through 2017 as colonies expanded (2015 = 0.7 km2, 2016 = 0.88 km2, 2017 = 2.68 km2). In 2017, the total extent of prairie dog colonies on public lands within our study area was > 160 km2, which at the time represented the largest known colony complex in the world. Individual colonies within the complex were up to 40 km2 in size, which is an order of magnitude larger than the size of prairie dog colonies typically found on National Grasslands in the western Great Plains over the past two decades (see Cully et al. 2010; Johnson et al. 2011). This wide range in prairie dog colony size over space and time within our study area provided a unique opportunity to examine how colony distribution and size influenced the grassland bird community.
Study design
To assess bird responses to prairie dog disturbance in this landscape, we used a point-transect-based sampling design (Fig. 1). Because TBNG is a patchwork of cover types, and our objective was to sample a wide range of locations that varied in terms of colony size and distance from the location to colony edges, we established transects using 3 different criteria. First, colony core transects (n = 10, 8 points per transect) were randomly placed with the constraint that transects fell entirely within prairie dog colonies. Second, colony edge transects (n = 41, 5–8 points per transect depending on colony size) were randomly located with the requirement that transects crossed the edge of a prairie dog colony with four points located outside the colony and one to four within the colony, depending on colony size. Third, sagebrush transects (n = 10, 8 points per transect) were located non-randomly in 10 known areas of extensive sagebrush habitat, which were identified during past surveys of greater sage-grouse (Centrocercus urophasianus), to ensure our surveys included portions of the landscape supporting spatially extensive stands of sagebrush. All transects contained 5–8 points spaced 250 m apart, for a total of 61 transects containing 439 survey points. We conducted two rounds of avian surveys between mid-May and late June each year from 2015 to 2017, surveying between sunrise and 10:00 AM on days with low wind and no rain (Pavlacky et al. 2017). By traveling to many points via off-road vehicle we ensured more effective detection of mountain plovers, which display more cryptic behavior in response to observers on-foot (Dinsmore et al. 2002). To adjust for detectability, we modeled avian abundance using Program DISTANCE (version 6.0) for seven focal species. We categorized species as either generalists or specialists within shortgrass, mid-grass, or sagebrush systems (Fig. 2), and compared models including time of survey, wind, temperature, observer, travel method (ATV versus on foot), and visual obstruction. The latter was incorporated to specifically address potential detectability differences between sites, as prairie dog colonies typically have lower visual obstruction (Duchardt et al. 2018).

Species names and four-letter alpha codes for shortgrass, midgrass, and sagebrush bird species in the TBNG, eastern Wyoming, USA, 2015–2017. “Generalists” in this sense display wider niche breadth and tolerances of other habitats (e.g., lark bunting utilized both midgrass and sagebrush), whereas specialists were found only within that habitat. Superscripts indicate Wyoming species of Conservation Need (A, WGFD 2010) and U.S. Forest Service Sensitive Species (B, U.S. Forest Service 2017)
Vegetation and landscape composition
We collected vegetation data at each survey point following point counts. We recorded line-point intercept data every meter along 30-m transects radiating from each point, perpendicular to the axis of the point count transect (Herrick et al. 2009). These data consisted of basal and canopy hits for perennial grasses, annual grasses, cacti, forbs, and shrubs. Ground cover categories included bare ground, litter, biological soil crust (BSC), and lichen, in addition to basal cover of vegetation classes. We measured visual obstruction, a metric incorporating both vegetation height and density, using a Robel pole (Robel et al. 1970) placed at 5-m increments along transects. In 2015, we collected shrub and cactus canopy cover data along these transects using the line-intercept method (Canfield 1941; Herrick et al. 2009). Because shrub canopy cover likely varies minimally over 1–2 years, we used these data to calculate percent cover of sagebrush and cactus at each point and used this value for all years.
Prairie dog colony boundaries in the Thunder Basin have been either partially or fully mapped by the USFS and partners since 2001 (Cully et al. 2010), mapping the perimeter of each colony using a GPS unit to walk between exterior active burrows (Sidle et al. 2012). Because most colony growth occurs in summer and early fall (Garrett and Franklin 1988; Milne-Laux and Sweitzer 2006), and birds select breeding habitat in the spring, we used mapped boundaries from the previous year to represent colony habitat in a given year (i.e., bird data collected in spring 2016 were paired with colony boundaries from fall 2015). Using these data, we generated multiple metrics of colony disturbance including distance to colony edge (positive inside, negative outside) as well as a binary measure of disturbance presence/absence. Prairie dog disturbance, especially that engendered by black-tailed prairie dogs, differs from many other sources of biotic or abiotic disturbance in that it is continuous and additive—areas that have been colonized for 10 years generally differ in terms of vegetation structure and composition from those only colonized for two (Garrett and Franklin 1988; Johnson-Nistler et al. 2004). Colonies have been expanding in Thunder Basin since 2006 (Cully et al. 2010), so those areas colonized for multiple years have experienced a longer period of grazing and burrowing pressure from prairie dogs than more recently colonized areas. As such, we also calculated the distance to edge for each point count location in each year, and a binary metric representing whether the location was colonized prior to or after 2013 (“long-term” colony cover).
Because we were also interested in how disturbance may affect birds at scales beyond an individual colony, we examined whether abundance was related to the proportion of the landscape occupied by prairie dogs surrounding any given point (hereafter referred to as landscape occupancy) or the density of prairie dog colony edges within the vicinity of any given point (hereafter edge density) at four spatial scales (100 m, 225 m, 500 m, 1000 m). The 100 m and 225 m scales contain the average territory size of the smallest (grasshopper sparrow [Ammodramus savannarum; Vickery 1996] and largest (mountain plover [Knopf and Wunder 2006]) focal species, while the larger scales may capture sensitivity to patch size during spring settlement (e.g., Hutto 1985).
Data analyses
To evaluate what vegetation characteristics best differentiate between prairie dog colonies and undisturbed habitat at a grassland-sagebrush ecotone (Q1), we used a logistic model to explain colony presence-absence as a function of a suite of characteristics of both vegetation structure (i.e., visual obstruction and bare ground) and composition (e.g., sagebrush cover). We modeled this same response for long-term colony cover and used a general linear model to examine distance to core colony.
To examine whether and how colony-associated birds responded to cover of their preferred habitat, and conversely how species preferring more dense vegetation (mid-grass and sagebrush birds) responded to prairie dog disturbance (Q2), we used a Generalized Additive Mixed Model framework in the gamm4 package in R (Wood and Scheipl 2017). Generalized additive models (GAMs) extend off generalized models using a non-parametric smoothing term to fit the data (Hastie and Tibshirani 1986) and are well-suited to detecting threshold responses (e.g., Large et al. 2013), and a mixed-model framework (GAMM) can be used to incorporate random effects (Wood 2011). Furthermore, they are well-suited to examining ecological patterns, which are rarely linear in nature (e.g., Bestelmeyer et al. 2011).
We tested three main hypotheses concerning the abundance of rangeland bird species: that prairie dog disturbance would have no effect (Q2H0), that current or long-term colony cover would explain variation in abundance but distance to edge (either inside or outside) would not (Q2H1C, Q2H1LT,), and finally that distance to either current or long-term edges would influence abundance (Q2H2LT, Q2H2C). We also compared both linear and non-linear models testing H2. We used an AIC model framework (Burnham and Anderson 2002) to evaluate the strength of these 7 models for each of our focal species, ranking models based on model weight. All models included a fixed effect of year and topographic roughness, as the latter has been shown to be important for habitat use for many of these species (Duchardt et al. 2018). We also incorporated a random effect of transect to address spatial autocorrelation (Legendre and Legendre 1998). We examined semivariograms and calculated Moran’s I (Moran 1950) for model residuals to ascertain that this variable did significantly reduce autocorrelation in the dataset. For species with < 2 observations in a given strata (i.e., inside or outside colony), we excluded data from that strata in our models.
We used an approach similar to that described above to compare hypotheses concerning avian response to landscape occupancy of prairie dogs and colony edge density at four spatial scales. Our hypotheses were that colony cover at broader spatial scales would have no effect beyond the presence or absence of disturbance, or distance to edge (Q3H0), that the total area (composition) of colony cover would influence abundance beyond the presence or absence of current (Q3H1c) or long-term (Q3H1LT) disturbance, and that current (Q3H2c) or long-term (Q3H2LT) edge density (configuration) would affect abundance. All models included a random effect of transect identity (see above) and a fixed effect of year and topographic roughness. We compared these models for each focal species using AICc (Burnham and Anderson 2002). We did not combine models representing Q2 and Q3, because correlation (r) between distance to edge and total colony cover in the landscape was high (0.6–0.73), which was unsurprising given larger colonies will have greater interior distance.
Results
Q1. Colony vegetation composition and structure
Prairie dog colonies differed substantially from undisturbed habitat, and were characterized by lower visual obstruction, less sagebrush and grass cover, and more forb cover (Table 1, See Electronic Supplementary material Appendix 1). Compared with undisturbed habitat, colonies also had substantially more bare ground and cactus cover. Areas of long-term prairie dog disturbance did not differ markedly from more recently colonized area in terms of bare ground but did have much lower visual obstruction and sagebrush cover. For survey points located on colonies, distance to the colony edge was not associated with variation in vegetation structure. Sagebrush and grass cover declined while forb cover increased within increasing distance from colony edge (i.e., as one moves toward the center of large colonies; Table 1).
Q2. and Q3. Avian abundance
Best-fitting models of detectability varied by species, with the basic hazard rate model describing detectability of sage thrashers and horned larks, while lark buntings were best described by the uniform function. Mountain plover detectability was explained by use of ATV on surveys and wind. Western meadowlark detectability varied with observer, while grasshopper sparrows varied by observer and start time. Finally, Brewer’s sparrow detectability varied with visual obstruction. The best model for each species had good fit based on a Chi squared test (P > 0.1).
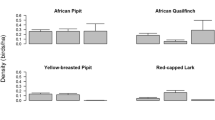
Horned larks, Brewer’s sparrows, and western meadowlarks (Sturnella neglecta) were the three most common species based on distance-adjusted densities across habitats (Fig. 3). Horned larks were the most abundant species on colonies (\(\bar{x}\) = 54 km−2, 95% CI = 51.2–57.2) and Brewer’s sparrows the most abundant in undisturbed habitat (\(\bar{x}\)= 69 km−2, 95% CI = 62.5–76.4). Raw distance-adjusted densities differed markedly between disturbed and undisturbed habitat for all species (Fig. 3). Mountain plovers occurred almost exclusively on colonies (2.9 km−2 on colonies [95% CI = 2.3–3.5], with only one observation off-colony). All other species were less abundant on versus off prairie dog colonies (Fig. 3), although western meadowlarks still reached moderate abundances on colonies (17 km−2, 95% CI = 16.1–18.1).

Avian densities (log scale) inside and outside current prairie dog colonies across 3 years in the TBNG, eastern Wyoming, USA, 2015–2017. Bars represent 95% CI
Abundances of all seven species were well-explained by models representing prairie dog disturbance, especially distance to colony edge, and there was little uncertainty in selection of the most parsimonious model for each species (Table 2; Fig. 4). Sage thrashers (Oreoscoptes montanus) and Brewer’s sparrows responded most strongly to long-term prairie dog disturbance with lower abundances on colonies (Q2H1LT), whereas grasshopper sparrows showed lower abundance on current colonies (Q2H1C). More generalist species showed a non-linear response to distance to colony edge, with lark buntings (Calamospiza melanocorys) declining with distance to current edge (Q2H2C, Fig. 4), and western meadowlarks declining with increasing distance (from negative outside to positive inside) to historic edge (Q2H2LT).

Top models of avian abundance (transformed to density per km2) in response to distance to current colony edge or historic colony edge of seven species in TBNG, eastern Wyoming, USA, 2015–2017. Envelopes in single-species figures represent 95% confidence envelopes. Grey shading indicates prairie dog colony. Bottom right figure combines models of abundance in response to current edge for all seven species (even where this was the second-best model). See Fig. 2 for alpha codes
Models for shortgrass-associated bird species also showed strong support for a non-linear relationship with distance to colony edge (Q2H2). Horned larks were more sensitive to historic edges, and although they were abundant throughout the study area were 4.6 times more abundant within colonies. Plovers were so rare off colonies (only 1 detection in 3 years of surveys) that their abundance could only be modelled as a function of distance to edge within colonies; we found that plover density at first increased with distance to edge, reaching maximum density at approximately 500 m, and declining sharply beyond 800 m (Fig. 4). Densities were generally high at distances of ~ 350–650 m from an edge. This relationship predicts that for an approximately circular colony, mountain plover density was maximized on colonies ~ 1.1 km2 (110 ha) in size and declined in abundance in the center of colonies > 1.5 km2.
Total long-term cover of prairie dog colonies (landscape occupancy) also affected abundance of five bird species, although no landscape occupancy models improved upon models from Question 2 for lark buntings or sage thrasher (Table 3). There was evidence of a linear positive effect of overall long-term colony cover for plovers at the 100 m and 225 m scale, correlating with territory size in this species (Q3H1LT), and non-linear increases in horned larks at the 225 m scale. Grasshopper sparrows and Brewer’s sparrows declined non-linearly with increasing long-term colony cover at the 500 m and 100 m scale, respectively, while meadowlarks declined linearly read at the 225 m scale. We did not find support for an effect of colony edge density on any species at any scale (Q3H2).
Discussion
Avian responses to prairie dog disturbance
We found that the presence versus absence of disturbance by prairie dogs was not enough to explain abundances of focal species in this system. Rather, landscape context of disturbance and disturbance duration influenced both sagebrush and grassland birds, albeit in different ways. For many of these species, this is the first example of landscape-scale response to prairie dog disturbance. For others, it provides evidence of new spatial patterns of abundance that prior research had not identified. Most notably, studies throughout the western Great Plains have shown that mountain plovers increase in abundance on black-tailed prairie dog colonies (Dinsmore et al. 2005; Goguen 2012; Augustine and Baker 2013), and are more abundant on colonies > 0.8 km2 in size, compared to colonies < 0.8 km2 (Goguen 2012; Augustine and Skagen 2014). This suggests that “bigger is always better” regarding the relationship between mountain plovers and prairie dog colonies. However, to our knowledge our study is the first to document a decline in mountain plover density towards the center of extremely large colonies, or a “goldilocks response” to distance to colony edge (Fig. 4). We note that “large” colonies in the previously mentioned studies varied from ~ 0.8 to 4.8 km2 (80–480 ha) in size. Average colony size throughout our study in Thunder Basin fell within this range (see Methods), whereas “large” colonies were 10 times larger. Indeed, plover density in Thunder Basin was maximal in the range of 350–650 m from a colony edge, was similar near colony edges and areas ~ 800 m from edges and declined below this level over distances of 800–1200 m from edges.
How this result translates into a relationship between plover density and colony size will depend heavily on colony geometry, but as a first approximation a circular colony with radius of 650 m (i.e., within the range of maximum plover abundance) would be 1.32 km2 in size, which corresponds to the mean size of large colonies supporting high plover densities in other localities (Dinsmore et al. 2005; Augustine et al. 2008; Goguen 2012; Augustine and Skagen 2014). In reality, colonies show high complexity in shape due to variation in vegetation and topography, so colonies with > 650 m to nearest colony edge will generally be much larger. For example, in 2015 the colony with highest mountain plover density was 3.2 km2 in size, but no point in the colony was greater than 650 m from an edge due to irregular colony shape. Thus, only when colonies begin exceeding many hundreds of hectares will they contain substantial area that exceeds 800 m from a colony edge.
The unique nature of our study area allowed us to detect a threshold beyond which very large colonies supported reduced mountain plover density. Our data do not allow us to directly test drivers of this response, but we here consider three potential mechanisms. Vegetation structure at colony cores was distinct from edges (Table 1), and plovers may avoid areas that are too sparse, especially if insect biomass is lower in these areas (Schneider et al. 2006). We did not detect avoidance above certain levels of bare ground, nor have other researchers (e.g., Knopf and Miller 1994; Goguen 2012), but measures of insect biomass on colonies may shed light on this idea in the future. Distance to colony edge may be a driver if edges provide resources that colony cores do not (Ries et al. 2004). It is unlikely that > 800 m is an infeasible distance for adults to travel for foraging opportunities given evidence plovers can move more than 1 km to preferred foraging sites (Woolley 2016). More likely, edges represent better cover for brood-rearing (Schneider et al. 2006), and potential thermoregulatory benefits for both chicks and adults (Graul 1975; Shackford 1996), but increased distance to these resources increases risk associated with reaching them, especially for unfledged chicks. Such benefits have been tied directly with shrubs (Schneider et al. 2006), which were much more abundant outside of colonies (\(\bar{x}\) = 1.3–7.3% cover). We also found that plover density declined near colony edges relative to areas 100–650 m from an edge, which could potentially be related to increased predation risk close to edges.
The lack of shrub cover within colonies also explains sagebrush bird avoidance of areas of long-term prairie dog disturbance. Long-term disturbance leads to prolonged clipping and girdling of sagebrush by prairie dogs, explaining extremely low sagebrush cover on these sites (\(\bar{x}\) = 0.6%) and resulting avoidance by sagebrush specialists like the Brewer’s sparrow and sage thrasher. It’s important to note that neither species responded to distance to colony edge, and only Brewer’s sparrows decreased with increasing long-term cover at a 100-m scale. These results indicate that sagebrush specialists overall show low sensitivity to the spatial arrangement of colonies in the landscape, responding mainly to local availability of sagebrush.
We also note that densities of Brewer’s sparrows and sage thrashers were lower in the TBNG (including undisturbed habitat) than many other areas throughout their range (Rotenberry et al. 1999; Reynolds 1999; Aldridge et al. 2011). It would be challenging to untangle the ubiquitous presence of disturbance in this landscape from the fact that it represents range-edge for both species (Rotenberry et al. 1999; Reynolds 1999). Species abundances are generally lower near range boundaries (Andrewartha and Birch 1954; Kirkpatrick and Barton 1997), and the fact that this is indeed a range edge for sagebrush birds is underscored by the absence of sagebrush sparrows (Artemisiospiza nevadensis) on these sites, a species that is found along with sage thrashers and Brewer’s sparrow throughout many portions of the sagebrush steppe (Martin 1998).
Generalists like western meadowlarks and lark buntings were sensitive to distance to colony edge (Fig. 4). Because colonies grow and expand into undisturbed habitat, more central, “core” areas tend to be older, and locations radiating from the core represent a chronosequence of decreasing age (Johnson-Nistler et al. 2004). The vegetational distinctiveness of these older areas helps explain the non-linear edge responses in many of Thunder Basin’s birds. Although both will nest near shrubs or taller grasses, lark buntings and western meadowlarks are primarily ground nesters and foragers. As such, they are less sensitive to the absence of shrubs within colonies and are tolerant of the low-intensity disturbance present at colony edges as long as some grass and litter cover is still present (Davis and Lanyon 2008; Augustine and Derner 2015).
While shortgrass birds increased with increasing long-term colony cover, midgrass and sagebrush birds mainly responded to long-term colony cover at more local scales, highlighting the importance of disturbance duration in this system. Conversely, few species responded to total colony cover at broader scales or to current colony cover, and none appeared sensitive to edge density. At broader scales, it appears that most avian species respond primarily to the extremely sparse cover of patches that have experienced multiple years of disturbance, and are less sensitive to short-term fluctuations in prairie dog disturbance.
Implications for prairie dog management
While few black-tailed prairie dog colony complexes reach the size of those observed in Thunder Basin (but see Ceballos et al. 2010), prairie dog conservation to this point has generally taken a “bigger is better” approach. Efforts to reintroduce endangered black-footed ferrets target complexes > 2000 ha (20 km2) in size (Roelle et al. 2005), with the underlying assumption that suitability for ferrets, and other colony associates, increases directly with size (Houston et al. 1986). Our results suggest that this may not be the case for the mountain plover, a species of conservation concern throughout its limited range (Birdlife International 2017). Further, our data show that sagebrush-associated birds are present but rare on prairie dog colonies—although these species may tolerate some large colonies within this landscape, their populations cannot be sustained without undisturbed sagebrush. The greater sage-grouse, another species of concern in this landscape, may also be negatively impacted if colonies become too large. Though sage-grouse have been observed using small colonies for lekking sites (D. Pellatz, personal communication), they rely on large patches of sagebrush for nesting, brood-rearing, and overwintering (Connelly et al. 2011; Knick and Hanser 2011), and this habitat declines with long-term prairie dog disturbance. Given the past and ongoing conversion of rangelands to croplands in the Great Plains (Wright and Wimberly 2013), conservation of these multiple bird guilds on a declining land base highlights the need to carefully consider the size and spatial dispersion of prairie dog disturbance on the landscape.
Beyond the responses of individual species to large colonies, colony size may be related to transmission of sylvatic plague, and thus the sustainability of prairie dog colonies in this landscape (Collinge et al. 2005; Johnson et al. 2011). Plague was introduced to North America over 100 years ago (Antolin et al. 2002) and is now among the major drivers of prairie dog declines. Plague vaccines (Abbott et al. 2012) and dusting to kill fleas that carry plague (Seery and Biggins 2003) have had some success in controlling these outbreaks, but there is evidence that the probability and intensity of outbreaks is greater as colony size increases (Collinge et al. 2005). It is important to note that “large” colonies in Collinge et al.’s study were 0.4–0.5 km2 in size, or two orders of magnitude smaller than large colonies in the Thunder Basin, and thus these effects may be magnified in the Thunder Basin landscape. While plague can be contracted by black-footed ferrets (Williams et al. 1994), it does not directly affect colony-associated avifauna (Antolin et al. 2002). However, large epizootics can decimate local prairie dog populations, and birds relying on engineered habitat structure (Augustine et al. 2008; Dinsmore and Smith 2010) or prairie dogs as a food resource (Seery and Matiatos 2000), often decline or disappear as a result. Thus, in the long-term, an important management consideration is how to maintain colonies in a spatial pattern that mitigates the “boom-and-bust” cycle of colony growth and plague, which has been documented in Thunder Basin and other landscapes over the past several decades (Cully et al. 2010).
A final key consideration for managing colony size and distribution in this landscape is the potential impact on livestock production and the local ranching community (Derner et al. 2006). Other studies have shown that stakeholders that coexist with prairie dogs generally consider them as pests (Lybecker et al. 2002), and ranchers especially see them as a threat to cattle production (Reading and Kellert 1993; Miller et al. 2007). While most stakeholders acknowledge a need for some prairie dogs for maintenance of wildlife populations, there can be disagreement concerning management, and concern for a loss of control over livestock management (Reading and Kellert 1993). In the Thunder Basin, rapid colony growth over a three-year period led to significant concerns within the ranching community (Ruckelshaus Institute 2017), and an undermining of trust between public and private stakeholders. Our findings regarding the threshold responses of both sagebrush-associated birds and mountain plovers to the spatial pattern of prairie dog disturbances suggest that management to minimize the presence of colonies containing areas > 800 m from colony edges is consistent with goals to conserve multiple avian guilds while also reducing conflict with livestock producers and reducing the potential for plague transmission across the landscape.
References
Abbott R, Osorio J, Bunck C, Rocke T (2012) Sylvatic plague vaccine: a new tool for conservation of threatened and endangered species? EcoHealth 9:243–250
Aldridge CL, Hanser SE, Nielsen SE et al (2011) Detectability adjusted count models of songbird abundance. In: Hanser SE, Leu M, Knick ST, Aldridge CL (eds) Sagebrush ecosystem conservation and management: ecoregional assessment tools and models for the Wyoming basins. Allen Press, Lawrence, pp 141–220
Allred BW, Smith WK, Twidwell D et al (2015) Ecosystem services lost to oil and gas in North America. Science 348:401–402
Andrewartha HG, Birch LC (1954) The distribution and abundance of animals. University of Chicago Press, Chicago
Antolin MF, Gober P, Luce B, Biggins DE, Van Pelt WE, Seery DB, Lockhart M, Ball M (2002) The influence of sylvatic plague on North American wildlife at the landscape level, with special emphasis on black-footed ferret and prairie dog conservation. In: Transactions of the North American Wildlife and Natural Resources Conference, vol 67, pp 104–127
Augustine D, Baker B (2013) Associations of grassland bird communities with black-tailed prairie dogs in the North American Great Plains. Conserv Biol 27:324–334
Augustine DJ, Derner JD (2015) Patch-burn grazing management, vegetation heterogeneity, and avian responses in a semi-arid grassland. J Wildl Manag 79:927–936
Augustine DJ, Dinsmore SJ, Wunder MB et al (2008) Response of mountain plovers to plague-driven dynamics of black-tailed prairie dog colonies. Landscape Ecol 23:689–697
Augustine D, Skagen S (2014) Mountain plover nest survival in relation to prairie dog and fire dynamics in shortgrass steppe. J Wildl Manag 78:595–602
Baker WL (2006) Fire and restoration of sagebrush ecosystems. Wildl Soc Bull 34:177–185
Beck JL, Connelly JW, Wambolt CL (2012) Consequences of treating Wyoming big sagebrush to enhance wildlife habitats. Rangel Ecol Manag 65:444–455
Bestelmeyer BT, Ellison AM, Fraser WR et al (2011) Analysis of abrupt transitions in ecological systems. Ecosphere. https://doi.org/10.1890/es11-00216.1
BirdLife International (2017) Charadrius montanus (amended version of 2016 assessment). The IUCN Red List of Threatened Species 2017
Brawn JD, Robinson SK, Iii FRT (2001) The role of disturbance in the ecology and conservation of birds. Annu Rev Ecol Syst 32:231–276
Burnham KP, Anderson DR (2002) Model selection and Multi-model inference: a practical-theoretical approach. Springer, New York
Canfield RH (1941) Application of the line intercept method in sampling range vegetation. J Forest 39:388–394
Ceballos G, Davidson A, List R et al (2010) Rapid decline of a grassland system and its ecological and conservation implications. PLoS One 5:e8562
Chambers JC et al (2016) Using resilience and resistance concepts to manage threats to sagebrush ecosystems, Gunnison sage-grouse, and greater sage-grouse in their eastern range: a strategic multi-scale approach. Gen Tech Rep RMRS-GTR-356. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, p 143
Collinge SK, Johnson WC, Ray C et al (2005) Landscape structure and plague occurrence in black-tailed prairie dogs on grasslands of the western USA. Landscape Ecol 20:941–955
Connelly JW, Rinkes ET, Braun CE (2011) Characteristics of greater sage-grouse habitats: a landscape species at micro and macro scales. In: Knick ST, Connelly JW (eds) Greater sage-grouse: ecology and conservation of a landscape species and its habitats, studies in Avian Biology (vol 38). University of California Press, Berkeley, pp 69–84
Cully JF, Collinge SK, Van Nimwegen RE et al (2010) Spatial variation in keystone effects: small mammal diversity associated with black-tailed prairie dog colonies. Ecography 33:667–677
Davidson AD, Lightfoot DC (2007) Interactive effects of keystone rodents on the structure of desert grassland arthropod communities. Ecography 30:515–525
Davis SK, Lanyon WE (2008) Western meadowlark (Sturnella neglecta), version 2.0. In: Poole AF (ed) The birds of North America. Cornell Lab of Ornithology, Ithaca
Derner JD, Detling JK, Antolin MF (2006) Are livestock weight gains affected by black-tailed prairie dogs? Front Ecol Environ 4:459–464
Desmond M, Savidge J, Eskridge KM (2000) Correlations between burrowing owl and black-tailed prairie dog declines: a 7-year analysis. J Wildl Manag 64:1067–1075
Dinsmore S, Smith M (2010) Mountain Plover responses to plague in Montana. Vector Borne Zoonotic Dis 10:37–45
Dinsmore SJ, White GC, Knopf FL (2002) Advanced techniques for modeling avian nest survival. Ecology 83:3476–3488
Dinsmore SJ, White GC, Knopf FL (2005) Mountain plover population responses to black-tailed prairie dogs in Montana. J Wildl Manag 69:1546–1553
DiTomaso JM, Monaco TA, James JJ, Firn J (2017) Invasive plant species and novel rangeland systems. In: Briske D (ed) Rangeland systems: processes, management, and challenges. Springer, Cham, pp 429–465
Dobson A, Lyles A (2000) Black-footed ferret recovery. Science 288:985–988
Duchardt CJ, Porensky LM, Augustine DM, Beck JL (2018) Disturbance shapes grassland bird communities on a grassland-sagebrush ecotone. Ecosphere 9(10):e02483. https://doi.org/10.1002/ecs2.248
Fuhlendorf SD, Engle DM (2001) Restoring heterogeneity on rangelands. Bioscience 51:625–632
Fuhlendorf SD, Engle DM, Elmore RD et al (2012) Conservation of pattern and process: developing an alternative paradigm of rangeland management. Rangel Ecol Manag 65:579–589. https://doi.org/10.2111/REM-D-11-00109.1
Fuhlendorf SD, Fynn RWS, McGranahan DA, Twidwell D (2017) Heterogeneity as the basis for rangeland management. In: Briske DD (ed) Rangeland systems: processes, management, and challenges. Springer, Cham, pp 169–176
Garrett MG, Franklin WL (1988) Behavioral ecology of dispersal in the black-tailed prairie dog. J Mammal 69:236–250
Goguen CB (2012) Habitat use by mountain plovers in prairie dog colonies in northeastern New Mexico. J F Ornithol 83:154–165. https://doi.org/10.1111/j.1557-9263.2012.00365.x
Graul WD (1975) Breeding biology of the mountain plover. Wilson Bull 87:6–31
Hastie TT, Tibshirani RR (1986) Generalized additive models. Stat Sci 1:297–310
Herrick JE, Van Zee JW, Havstad KM et al (2009) Monitoring manual for grassland, shrubland and savanna ecosystems, volume 1, quick start. University of Arizona Press, Tucson
Hoogland JL (1995) The black-tailed prairie dog: social life of a burrowing mammal. The University of Chicago Press, Illinois
Houston BR, Clark TW, Minta SC (1986) Habitat suitability index model for the black-footed ferret: a method to locate transplant sites. Great Basin Nat Mem 8:99–114
Hutto RL (1985) Habitat selection by nonbreeding, migratory landbirds. In: Cody ML (ed) Habitat selection in birds. Academic Press, New York, pp 455–476
Johnson TL, Cully JF, Collinge SK et al (2011) Spread of plague among black-tailed prairie dogs is associated with colony spatial characteristics. J Wildl Manag 75:357–368. https://doi.org/10.1002/jwmg.40
Johnson-Nistler CM, Sowell BF, Sherwood HW, Wambolt CL (2004) Black-tailed prairie dog effects on Montana’s mixed-grass prairie. J Rangel Manag 57:641–648
Kirkpatrick M, Barton NH (1997) Evolution of a species’ range. Am Nat 150:1–23
Knick ST, Hanser SE (2011) Connecting pattern and process in greater sage-grouse populations and sagebrush landscapes. In: Knick ST, Connelly JW (eds) Greater sage-grouse: ecology and conservation of a landscape species and its habitats, studies in avian biology, vol 38. University of California Press, Berkeley, pp 383–406
Knick ST, Holmes AL, Miller RF (2005) The role of fire in structuring sagebrush habitats and bird communities. Stud Avian Biol 30:1–13
Knopf FL, Miller BJ (1994) Charadrius montanus–montane, grassland, or bare-ground plover? Auk 111:504–506
Knopf FL, Wunder MB (2006) Mountain plover (Charadrius montanus). In: Poole A (ed) In the birds of North America online. Cornell Laboratory of Ornithology, Ithaca
Kotliar N, Baker B, Whicker A, Plumb G (1999) A critical review of assumptions about the prairie dog as a keystone species. Environ Manag 24:177–192
Large SI, Fay G, Friedland KD, Link JS (2013) Defining trends and thresholds in responses of ecological indicators to fishing and environmental pressures. ICES J Mar Sci 70:755–767
Legendre P, Legendre L (1998) Numerical ecology, 2nd edn. Elsevier, Amsterdam
Lomolino M, Smith G (2004) Terrestrial vertebrate communities at black-tailed prairie dog (Cynomys ludovicianus) towns. Biol Conserv 115:89–100
Lybecker D, Lamb BL, Ponds PD (2002) Public attitudes and knowledge of the black-tailed prairie dog: a common and controversial species. Bioscience 52:607–613
Martin JW, Carlson BA (1998) Sagebrush sparrow (Artemisiospiza nevadensis), version 20. In: Poole AF (ed) The birds of North America. Cornell Lab of Ornithology, Ithaca
Miller S, Cully J (2001) Conservation of black-tailed prairie dogs (Cynomys ludovicianus). J Mamm 82:889–893
Miller BJ, Reading R, Biggings D et al (2007) Prairie dogs: an ecological review and current biopolitics. J Wildl Dis 71:2801–2810
Milne-laux AS, Sweitzer RA (2006) Experimentally induced colony expansion by black-tailed Prairie dogs (Cynomys ludovicianus) and implications for conservation. J Mamm 87:296–303
Moran PAP (1950) Notes on continuous stochastic phenomena. Biometrika 37:17
National Oceanic and Atmospheric Administration [NOAA] (2018) National Centers for Environmental Information. http://www.ncdc.noaa.gov/cdo-web/datasets#GHCND. Accessed 04 June 2016
Pavlacky DC, Lukacs PM, Blakesley JA et al (2017) A statistically rigorous sampling design to integrate avian monitoring and management within Bird Conservation Regions. PLoS One 12:1–22
Ponce-Guevara E, Davidson A, Sierra-Corona R, Ceballos G (2016) Interactive effects of black-tailed prairie dogs and cattle on shrub encroachment in a desert grassland ecosystem. PLoS One 11:e0154748
Porensky LM, Derner JD, Pellatz D (2018) Plant community responses to historical wildfire in a shrubland-grassland ecotone reveal hybrid disturbance response. Ecosphere 9(8):e02363
Reading RP, Kellert SR (1993) Attitudes toward a proposed reintroduction of black-footed ferrets (Mustela nigripes). Conserv Biol 7:569–580
Reynolds TD, Rich TD, Stephens DA (1999) Sage thrasher (Oreoscoptes montanus), version 2.0. In: Pooler AF, Gill FB (eds) The birds of North America. Cornell Lab of Ornithology, Ithaca
Riebsame WE, Gosnell H, Theobald DM (1996) Land use and landscape change in the Colorado Mountains I: theory, Scale, and Pattern. Mt Res Dev 16:395
Ries L, Fletcher RJJ, Battin J, Sisk TD (2004) Ecological responses to habitat edges: mechanisms, models, and variability explained. Annu Rev Ecol Syst 35:491–522
Robel RI, Briggs JN, Dayton AD, Hulbert LC (1970) Relationships between visual obstruction measurements and weight of grassland vegetation. J Rangel Manag 23:295–297
Roelle JE, Godbey JL, Biggins DE (2005) Recovery of the Black-footed Ferret: progress and continuing challenges. In: Proceedings, symposium status black-footed ferret its habitat, Fort Collins, Color, p 288
Rotenberry JT, Patten MA, Preston KL (1999) Brewer’s Sparrow (Spizella breweri), version 2.0. In: Poole AF, Gill FB (eds) The Birds of North America. Cornell Lab of Ornithology, Ithaca
Ruckelshaus Institute (2017) Thunder basin National Grassland collaboration report. University of Wyoming, Laramie
Sala OE, Yahdjian L, Havstad K, Aguiar MR (2017) Rangeland ecosystem services: nature’s supply and humans’ demand. In: Briske D (ed) Rangeland systems: processes, management, and challenges. Springer, Cham, pp 467–489
Sayre NF (2017) The politics of scale: a history of rangeland science. University of Chicago Press, Chicago
Schneider SC, Wunder MB, Knopf FL, Jones CA (2006) Relationship between shrubs and foods in mountain plover habitat in Park County, Colorado. Southwest Nat 51:197–202
Seery D, Biggins D (2003) Treatment of black-tailed prairie dog burrows with deltamethrin to control fleas (Insecta: Siphonaptera) and plague. J Med Entomol 40:718–722
Seery D, Matiatos D (2000) Response of wintering buteos to plague epizootics in prairie dogs. West North Am Nat 60:420–425
Shackford JS (1996) The importance of shade to breeding mountain plovers. Bull Oklahoma Ornithol Soc 29:17–21
Shipley B, Reading R (2006) A comparison of herpetofauna and small mammal diversity on black-tailed prairie dog (Cynomys ludovicianus) colonies and non-colonized grasslands in Colorado. J Arid Environ 66:27–41
Sidle JG, Augustine DJ, Johnson DH et al (2012) Aerial surveys adjusted by ground surveys to estimate area occupied by black-tailed prairie dog colonies. Wildl Soc Bull 36:248–256. https://doi.org/10.1002/wsb.146
U.S. Department of Agriculture (USDA) (2010). National resources inventory rangeland resource assessment, natural resources conservation service. US Department of Agriculture (USDA), Washington, DC
US Forest Service (2017). Wildlife, fish, and sensitive plant habitat management: threatened, endangered and sensitive plants and animals. Forest Service Manual. Rocky Mountain Region, Denver CO
Van Nimwegen R, Kretzer J Jr, Cully JF (2008) Ecosystem engineering by a colonial mammal: how prairie dogs structure rodent communities. Ecology 89:3298–3305
Vickery PD (1996) Grasshopper Sparrow (Ammodramus savannarum), version 20. In: Poole AF, Gill FB (eds) The birds of North America. Cornell Lab of Ornithology, Ithaca
Williams ES, Mills K, Kwiatkowski DR et al (1994) Plague in a black-footed ferret (Mustela nigripes). J Wildl Dis 30:581–585
Wood SN (2011) Fast stable restricted maximum likelihood and marginal likelihood estimation of semiparametric generalized linear models. J R Stat Soc Ser B Stat Methodol 73:3–36
Wood S, Scheipl F (2017) gamm4: Generalized additive mixed models using ‘mgcv’ and ‘lme4’. R package version 0.2–5
Woolley CA (2016) Mountain plover breeding ecology: home-range size, habitat use, and nest survival in an agricultural landscape, Thesis. University of Colorado, Denver
Wright CK, Wimberly MC (2013) Recent land use change in the western Corn Belt threatens grasslands and wetlands. Proc Natl Acad Sci 110:4134–4139
Acknowledgements
Funding for this project was provided by the USDA Agricultural Research Service, UW Agriculture Experiment Station, and Laramie Audubon Society. We thank J. Hennig, S. Green and S. Rankins for collecting field data, I.G. for data processing, and M. Murphy and C. Tarwater for consultation about data analyses. We also thank the U.S. Forest Service and the Thunder Basin Grassland Prairie Ecosystem Association for help coordinating field efforts. Thanks also to A. Meyer for a photo of an elusive, if not local, grasshopper sparrow. All other photos and figures by C. Duchardt.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Duchardt, C.J., Augustine, D.J. & Beck, J.L. Threshold responses of grassland and sagebrush birds to patterns of disturbance created by an ecosystem engineer. Landscape Ecol 34, 895–909 (2019). https://doi.org/10.1007/s10980-019-00813-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-019-00813-y