
Abstract
Context
The ecological interplay between edge and matrix effects along forest-matrix interfaces is closed linked to landscape processes modulating biodiversity and ecosystem services provision in disturbed landscapes. Improving such knowledge is therefore essential to design more efficient land use management in multifunctional landscapes.
Objectives
Estimate the avian-mediated Ecosystem Service Provision (ESP) in contrasting types of forest-matrix interfaces in tropical landscapes, and examine how local habitat and landscape attributes can predict ESPs.
Methods
We sampled bird assemblages in forest-pasture (FP) and forest-eucalyptus plantation (FE) interfaces, and estimated their potential as pest control, seed dispersal and pollination agents across human-modified landscapes in southeastern Brazil. Using Random Forest algorithm, we also quantified the relative importance of local vs. landscape attributes in predicting ESPs.
Results
The overall ESPs was higher in FP than FE interfaces. Habitat generalist birds were important potential seed disperser and pollinator agents at both FP and FE. At forest edges, landscape forest cover best predicted pest control services, whereas the density of rural homesteads best predicted potential seed dispersal and pollination services. Local habitat features were particularly important in predicting all ESP in pastures.
Conclusions
We highlight the importance of matrix type and matrix habitat structure in modulating avian ESPs across forest-matrix interfaces in human-altered landscapes and suggest that forest cover and rural homestead density are key elements in multifunctional landscapes that consider avian ESP in both forest and matrix habitats.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Birds are involved in many ecological functions that are essential for the maintenance of Earth’s ecosystems (Sekercioglu et al. 2016). In both natural and anthropogenic habitats, birds provide direct and indirect ecosystem benefits to humans. In natural ecosystems, birds often play a pivotal role in ecosystem functioning (e.g. via mutualistic and antagonistic interactions), contributing to a balanced homeostasis of healthy ecosystems (Sekercioglu et al. 2016). In croplands, avian insectivores enhance crop yields by reducing pest abundance (Maas et al. 2016), particularly in areas near forests (Karp et al. 2013) or embedded within forest landscapes (Boesing et al. 2017). In fallow lands, avian seed dispersal agents generate substantial seed shadows, accelerating regeneration trajectories and contributing to ecosystem resilience (Bregman et al. 2016; Carlo and Morales 2016). Many tropical plant species are pollinated by avian nectivores that can also act as important pollinator agents in croplands (Sekercioglu et al. 2016).
Although birds are essential in maintaining important ecosystem functions and services, they are often severely threatened by human activities. In tropical regions, habitat loss and fragmentation have been considered the most important drivers of bird population declines (Haddad et al. 2015). Edge creation in tropical forests worldwide has severely contributed to edge-related bird declines (Pfeifer et al. 2017). Understanding the ecological roles of rapidly proliferating forest edges is critical, particularly considering that half of all remaining forest cover globally is now within 500 m of the nearest edge (Haddad et al. 2015). This pervasive pattern of edge domination is aggravated in the once continuous > 1.3 million km2 Atlantic Forest biome (Muylaert et al. 2018), nearly 50% of which is now within 100 m from any edge (Ribeiro et al. 2009). Incorporating meaningful estimates of context-specific ecosystem service provision (ESP) (i.e. the benefits people obtain from ecosystems: IPBES assessments, United Nations Environment Programme) can therefore be an important strategy in achieving more efficient land use management within fragmented forest landscapes (Sekercioglu et al. 2016).
In addition to the effects of anthropogenic edges, the matrix type (Kennedy et al. 2016; Biz et al. 2017; Boesing et al. 2018), landscape-wide forest cover (Banks-Leite et al. 2014), and landscape composition and heterogeneity (Carrara et al. 2015; Lee and Martin 2017) are important co-factors regulating bird persistence in human-modified landscapes. However, the poorly understood interactions between edge effects and matrix effects along forest-matrix interfaces is also a crucially important ecological process modulating bird persistence (Blitzer et al. 2012; Tscharntke et al. 2012; Boesing et al. 2018). The ability of birds and other organisms to spill over into neighbouring habitat patches (hereafter, cross-habitat spillover) can provide important cues on how forest species persist within human-altered landscapes (Boesing et al. 2018), enabling a better understanding of the consequences of anthropogenic habitats on ecosystem functioning and service provisioning (Tscharntke et al. 2012).
Several recent studies have examined the effects of land use intensification on avian foraging guilds. In tropical regions, avian insectivores, frugivores and nectarivores have been singled out as most sensitive trophic guilds to land-use change compared to other functional groups (Sekercioglu 2012; Newbold et al. 2013; Bregman et al. 2016). Also, specialists of core forest habitats are often replaced by habitat generalists in highly disturbed habitats (De Coster et al. 2015; Morante-Filho et al. 2015). However, little attention has been given to the effects of landscape modification on the strength of particular functions linked to ESP (Luck et al. 2012). Although recent studies have provided valuable insights on how land use change affects avian-mediated ES (e.g. Muñoz et al. 2013; Bregman et al. 2016; Bovo et al. 2018; Morante-Filho et al. 2018), none of them provide meaningful quantification on avian-mediated ESP (Wenny et al. 2011). Additionally, even less data are available on patterns of cross-habitat spillover by bird communities in tropical forest-matrix boundaries (Blitzer et al. 2012; Tscharntke et al. 2012; but see Boesing et al. 2018).
In southeast Brazil, cattle pastures and eucalyptus monocultures are among the most dominant anthropogenic matrix habitat types (Brockerhoff et al. 2012), and their relative importance to ESP by vertebrates remains poorly known (Sekercioglu et al. 2016). Although several studies have focused on the effects of tree plantations on bird communities, little is known about the relative importance of eucalyptus plantations compared to other ubiquitous matrix types such as livestock pastures (Lindenmayer et al. 2008; de Camargo Barbosa et al. 2017). Here, we used diet-related traits to assess differences on the potential flow of three key ecosystem services (insect pest control, seed dispersal and pollination) provided by birds inhabiting forest-pasture and forest-eucalyptus interfaces. We also examine which landscape and local habitat variables can best predict avian-mediated ESP within three types of habitat edges (forest, pasture and eucalyptus). Given that (1) eucalyptus plantations typically retain tall trees and a fairly closed canopy, and provide lower structural contrast with adjacent natural forests, and (2) birds inhabiting forest edges could be tolerant of external matrix habitats sharing similar vegetation structure, we expect that ESP by birds at forest-eucalyptus interfaces should be higher than those at forest-pasture interfaces.
Methods
Study region
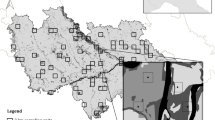
This study was conducted in a ~ 300,000-ha region of the state of São Paulo, southeast Brazil (W23°04′09″, S46°32′87″; Fig. 1), consisting of landscapes containing wide gradients of forest habitat loss and land use heterogeneity. The region was once entirely covered by the Atlantic Forest (Muylaert et al. 2018), but was subjected to intense resource exploitation leading to conversion of most natural vegetation into cropland, pastures and urban settlements (Morellato and Haddad 2000). Most of the study area is currently comprised of cattle pastures and eucalyptus plantations (Eucalyptus spp), with smaller areas of sugar-cane and other agricultural crops, including maze, citrus, and other fruit cultivation. In contrast to vast areas dominated by large-scale mechanized farming elsewhere in Brazil, our study landscapes are dominated by large numbers of smallholders and higher land-use heterogeneity because of a more dissected topography (Oliveira and Fontes 2000). Natural vegetation in this region consists primarily of uneven-aged but mature (>100 years-old) secondary forest fragments of varying patch sizes (most of which are highly disturbed), wetlands and regeneration areas (i.e. abandoned lands in early successional stage). The Köpper’s climate classification for the study region is Cwa (humid subtropical with a dry winter and hot summer), and elevation ranges between 700 and 1700 masl (Oliveira and Fontes 2000).

Study region showing the 32 bird sampling sites (black dots) surrounded by circular landscapes in southeastern Brazil (2014–2015). The green patches and white background represent native forest cover and the non-forest matrix, respectively. The inset higher-resolution landscapes (upper right) illustrate two examples of paired point-counts in forest-pasture (left) and forest-eucalyptus interfaces (right), both within 1.2-km radius circular landscapes classified according to land use types. The different colours represent different land use types. (Color figure online)
Sampling design
Bird assemblages were recorded at forest-matrix interfaces using 50-m fixed-radius point counts (Bibby et al. 2002). We sampled birds within 16 forest-eucalyptus and 16 forest-pasture interfaces, amounting to 32 sampling landscapes across the study region. Each sampling landscape included two paired sampling points, one within the forest edge and the other within the adjacent habitat matrix (eucalyptus or pasture). All paired sampling points were mirrored, i.e. they were located 140–200 m apart from each other on opposite directions at 70–100 m from the forest margin (Fig. 1).
At each sampling point, we recorded any bird seen or heard during 10 min per visit. Point-counts were repeated three times (amounting to 30 observation minutes per point) on different days from the onset of sunrise to the first three daylight hours, amounting to a total of 192 point-counts. All surveys took place during two breeding seasons (15 paired point-counts sampled from September 2014 to January 2015; and 17 paired point-counts sampled from October to December 2015). To minimize spatial autocorrelation, sampling sites were spaced by at least 2 km. To avoid potential spatio-temporal sampling biases (from an atypical annual climate, for instance), bird sampling was spatially randomized across the entire study area in both sampling years (i.e. not restricted to a particular portion of the study area at each sampling year). Sampling landscapes were selected to ensure a wide gradient of both forest cover (12–92%) and land use heterogeneity (Shannon diversity index; range 0.6–1.8).
Quantifying avian-mediated ecosystem services
We developed three community-based metrics related to the ability of bird assemblages to provide ecosystem services: pest control (PC), seed dispersal (SD) and pollination (PO). As such, we compiled dietary data (from Wilman et al. 2014) on the average proportion of invertebrates, fruits and nectar consumed by each bird species, which are used as proxies of potential avian provision of pest control, seed dispersal and pollination services, respectively. For each species, the ESP value was therefore estimated as the species absolute local abundance multiplied by the percentages of invertebrates, fruits and nectar in the diet. For each local species recorded, we define absolute abundance the maximum number of individuals recorded (seen or heard) among the three temporal repeats at each point-count site. At the assemblage level, the ESP metric (in aggregate and for each ES) was then calculated as the sum of ESP values for all species sampled at each assemblage. These metrics likely provide meaningful estimates of the strength of ecological functions and/or ES flows induced by any given bird assemblage (Luck et al. 2012). We also computed the bird species richness (SR) for each sampling point within landscapes as the total number of species detected.
All metrics aggregated for whole assemblages were also examined for either forest specialist and habitat generalist bird species. To do so, three highly experienced field ornithologists provided scores based on lifetime cumulative observations (> 60 years in total) on levels of forest dependency for all bird species detected [scores ranging from 0 (lowest) to 10 (highest)]. These scores reflected the probability of each species occurring within closed-canopy forest environments. According to the field knowledge of the experts consulted, those species with average forest occupancy values ranging from 0.0 to 7.0 were defined as habitat-generalists, whereas species scoring 7.1 or higher values were defined as forest-specialists. In any case, this threshold value produced a very accurate classification on bird habitat preference according to field experts.
Local and landscape predictors
Two local variables were considered as predictors of avian-mediated ESP: type of adjacent matrix and habitat structure. While the first is a categorical variable (pasture or eucalyptus), which was considered only when sampling points were located within forest sites, the latter was a simplified measure of vegetation structure and other important local environmental features that are known to affect habitat use by birds (e.g. presence of regenerating vegetation in eucalyptus, number of scattered trees in pastures, tree density in forests). From the local habitat variables sampled, we ran a principal component analysis (PCA) for each habitat type, wherein habitat structure values were defined as the resulting scores from the first axis that best explained the total variation in the data (forest 55.2%, eucalyptus 59.9% and pasture 51.3%). Higher habitat structure values therefore indicate overall higher structural complexity of matrix habitats. Details on local habitat sampling and the PCA analysis are provided in Supplementary Information S1.
As landscape predictors, we considered seven landscape variables: percentage of forest cover, edge density, Shannon diversity index of the matrix mosaic, percentage of area under silviculture (i.e. Eucalyptus plantation), percentage of pasture area, patch density of rural homesteads, and patch density of water bodies (e.g. lakes and reservoirs). Edge density and Shannon diversity are frequently used proxies of landscape heterogeneity (Fahrig et al. 2011). We defined rural homesteads as small patches containing warehouses, farmhouses, scattered trees, woodlots (including mainly exotic fruit trees), small ponds, cropland and livestock facilities. We included this metric because rural dwellings typically cluster several key avian resources (e.g. perches, nest sites, food) within relatively small areas. Functioning as stepping stones between habitat patches or even suitable habitat for several bird species (mainly habitat generalists), these dwellings are expected to facilitate landscape connectivity (Bennett et al. 2006) and ecosystem service provision by birds (Mitchell et al. 2013). Water bodies are usually surrounded by wetlands or riparian habitats that host high invertebrate abundance (Taft and Haig 2005) and may also act as stepping stones or foraging habitat for several bird species (mainly insectivores and/or habitat generalists). These landscape features may also favour avian ESP along forest-matrix interfaces within agriculture-dominated landscapes. We incorporated the percentage of silviculture and pasture areas in the analyses because these matrix types may regulate landscape connectivity for forest-specialist and generalist birds, respectively (de Camargo Barbosa et al. 2017), thereby influencing ESP along habitat interfaces.
To compute the landscape metrics, we first produced land cover and land use maps within 1200-m radial buffers from each forest sampling site using high-resolution images (ArcGIS 10.3 basemap imagery, DigitalGlobe satellites 2010–2011; scale of 1:5000). To do so, we considered 14 land-use cover classes (Fig. 1): forest, pasture, eucalyptus plantations, second-growth, wetland, cropland (mainly maze), sugar-cane, open-water, urban areas, rural homesteads, urban or suburban homesteads, paved roads, buildings, and bare soil. All ambiguous land cover in terms of image interpretation were checked and validated in the field. Once buffers were classified, we used Fragstats v.4 (McGarigal et al. 2012) to compute all landscapes metrics. We considered the 1200-m circular buffer size because it was the spatial scale (between 300 and 1200-m) that best explained landscape predictors to bird richness and abundance according to Random Forest algorithms (see Supporting Information S2).
Data analysis
We used pairwise t-tests to examine differences in ESP by bird assemblages between adjacent habitats (forest-pasture and forest-eucalyptus), and Mann–Whitney tests to examine differences between non-adjacent habitats (forest-forest or pasture-eucalyptus). To assess whether species straddled the boundaries between adjacent habitats, and the magnitude of ES potentially provided across forest-matrix interfaces, we computed the species richness and aggregate ESP values for species recorded only in forest, only in the matrix (either pasture or eucalyptus), and in both habitats. To calculate aggregate ESP estimates, we considered the incidence frequency, rather than the abundance, of all species throughout our study region as a quantitative measure within each habitat type. For any given species using any two adjacent habitat types, we were deliberately conservative by considering the lowest incidence frequency value across forest-matrix interfaces.
To assess the relative importance of environmental predictors of avian-mediated ESP, we used Random Forest algorithm (randomForest package in R) (Cutler et al. 2007; Triviño et al. 2011; Crisci et al. 2012). We ran 36 regression tree models, with nine predictor variables (two local-scale and seven landscape-scale) explaining four response variables (SR, PC, SD and PO) within three habitat types (forest, pasture and eucalyptus). Each model contained seven predictor variables, six of which (habitat structure, forest cover, edge density, Shannon diversity, water bodies and rural homesteads) were included in all models, whereas type of adjacent habitat, pasture cover and eucalyptus cover were included only in the forest, pasture and eucalyptus models, respectively.
We used Random Forest algorithms to compute the relative importance of predictors because they have been considered as the most powerful, accurate and simple estimators of variable importance (Cutler et al. 2007; Crisci et al. 2012). Using regression trees, one can calculate the relative importance among a large number of predictors and complex non-linear model relationships (Cutler et al. 2007; Triviño et al. 2011; Crisci et al. 2012). Moreover, this tool has a simple, intuitive interface, and can provide superior, or at least comparable, predictive power compared to generalized linear models (GLMs) or other conventional statistical modelling approach, even with small sample sizes (Cutler et al. 2007). All analyses were conducted in the R environment, version 3.3.1 (R Development Core Team 2016).
Results
In total, we recorded 200 bird species across all 32 sampling landscapes (64 paired point counts), 83.5% of which were found in forest edges, 52.2% in pastures, and only 24.5% in eucalyptus plantations. From all species detected, 54% were considered as forest-specialists and 46% as habitat-generalists. In terms of major dietary modes, 89% of all species consumed invertebrates, 50% fruits, and only 10% floral nectar (mainly hummingbirds).
Cross-habitat spillover and avian-mediated ESP
Overall, ESP modulated by birds was significantly higher in forest sites than in either matrix types (pasture or eucalyptus), except for pollination which did not show significant differences across forest-pasture interfaces (z = − 1.60, P = 0.10) (Table 1, Fig. 2). In general, there were no differences in mean ESP between the two types of forest interfaces (adjacent to pasture or to eucalyptus), except for habitat-generalists which had higher species richness and pest-control potential in forests adjacent to pastures than those adjacent to eucalyptus (z = − 2.87, P = 0.004 and z = − 2.11, P = 0.03, respectively, Table 1, Fig. 2). The total number of forest-specialist species in the two forest edge types was similar (84 and 91 species in forests adjacent to pastures and eucalyptus, respectively). However, the overall bird richness restricted to forest edges (i.e. those using only forest and not the adjacent matrix) was substantially higher in forests adjacent to eucalyptus (92 species) than in forests adjacent to pastures (68 species) (Fig. 3).

Species richness and potential ecosystem service provision (ESP) by birds within forest-pasture (N = 16) and forest-eucalyptus interfaces (N = 16), across human-modified landscapes in Brazil. Values are shown as means and standard deviations. (Color figure online)

Species richness and the potential ecosystem services provision (pest control, seed dispersal and pollination) by birds (all species, forest specialists and habitat generalists) in both forest-pasture and forest-eucalyptus interfaces across human-modified landscapes in the southeastern Brazil. Values indicate the total ecosystem services provision (ESP) by species restricted to a single habitat (forest, pasture or eucalyptus) or straddling the boundaries between habitats. (Color figure online)
Pasture sites exhibited significantly higher ESP estimates than eucalyptus stands (Table 1, Fig. 2). Overall, birds shared higher levels of SR and ESP across forest-pasture than across forest-eucalyptus interfaces, especially in relation to habitat-generalist seed dispersal vectors (Fig. 3). Eucalyptus plantations per se hosted very little or no unique ESP, which in all cases were provided by bird species that also used adjacent forests (Fig. 3). In contrast, pastures retained a significant pool of locally-unique countryside species, but all of those were habitat-generalists (Fig. 3).
The mean rate of ESP loss from forest to pasture was 35.3% (± 22.6%, standard deviation) for SR, 35.1% (± 25.4%) for PC, and 35.4% (± 31.7%) for SD. Despite a marked variation, there was actually a gain of 48% (± 123.1%) in estimated pollination at pastures sites (i.e. higher mean values in pastures than in adjacent forests). In contrast, there were consistent and substantial losses in ESP from forest to neighbouring eucalyptus plantations, in the order of 77.0% (± 6.9%), 82.8% (± 6.7%), 81.0% (± 14.9%), and 72.6% (± 17.5%) for SR, PC, SD and PO, respectively.
In general, forest-specialists provided higher overall contributions to ESP within forest sites, whereas habitat-generalists provided higher contributions in the adjacent matrix. However, both functional groups provided similar seed-dispersal contributions within forest areas, and in terms of SR and PC within eucalyptus plantations (Table 2). At forest sites, habitat-generalists provided an even higher contribution than forest-specialists in terms of PO services (Tables 1, 2).
Local and landscape effects on avian-mediated ESP
Regression trees performed by Random Forest algorithms showed a wide range of results, with the amount of variance explained ranging widely across models (Fig. 4, Table 2). At forest sites, overall forest cover was the best predictor of SR and PC, whereas rural homesteads were the best predictor of SD and PO (Fig. 4, Table 2). Forest cover was also the best predictor of all ESP provided by forest specialists at forest edges. In eucalyptus plantations, although models yielded very low amounts of variation explained (Table 2), metrics related to landscape heterogeneity—mainly Shannon diversity index—were the best predictors of all measures of ESP (Fig. 4, Table 2). In pastures, local habitat structure was the best predictor of all ESP metrics. Considering habitat-generalists across all habitats, ESP was best explained by rural homesteads, edge density, and Shannon index for matrix diversity. In particular, all ESP induced by habitat-generalists along forest edges was best explained by rural homesteads. Details of the Random Forest analysis, including all non-linear relationships between predictor and response variables are available in Supporting Information S2.

The relative importance of local and landscape predictors (different colours) on species richness and the potential ecosystem service provision (pest control, seed dispersal, pollination) by birds in forest edges, pastures and eucalyptus plantations across human-modified landscapes in southeastern Brazil. IncNodePurity values indicates the total decrease in node impurities averaged over all regression trees (i.e. higher values indicate higher predictive power). (Color figure online)
Discussion
Cross-habitat spillover and avian-mediated ESP
Forest environments are widely known to retain higher productivity and heterogeneity than structurally simplified and highly managed anthropogenic habitats, such as pastures or tree plantations (Newbold et al. 2015). However, comparing the two dominant types of matrix habitats examined here, eucalyptus monoculture had significant lower avian-induced ESP than pastures. Moreover, the overall loss of ESP was almost twice higher from forest to neighbouring eucalyptus than from forest to neighbouring pastures. These differences can be related primarily to local habitat structure within these two matrix types and, in any case, forest sites retained significantly higher levels of ESP than either their adjacent eucalyptus or pastures sites.
Local habitat structure was a decisive determinant of all ESP metrics at pasture sites (Table 2, Fig. 4). Given that both biotic and abiotic factors and proximity to water bodies are likely associated to resource availability for habitat-generalist countryside birds in man-made grazelands (Mahood et al. 2012), this likely induced higher ESP values in pastures compared to eucalyptus stands. Although frugivores may be unaffected by the spatial structure of tree cover in pastures near primary forest edges in the Amazon (Lasky and Keitt 2012), other studies have shown the importance of relict features in pastures to countryside birds (Mahood et al. 2012). Intensively managed pastures (e.g. those lacking scattered isolated trees), for instance, typically retain lower bird diversity than neglected pastures supporting lower livestock densities and containing higher structural complexity and scattered clusters of trees and shrubs (Mahood et al. 2012). Given that our sampling design excluded intensively managed homogeneous cattle pastures, the higher ESP estimates for pastures, mainly in terms of seed dispersal and pollination services, likely applies primarily to pastures with some degree of structural complexity.
The higher vegetation structural contrast between forest and pastures may contribute to the higher overall ESP values across these habitats compared to forest-eucalyptus interfaces (Table 1). Forest edges adjacent to open habitats are more exposed to sunlight, favouring light-demanding plant species, such as shrubs and pioneer trees (e.g. Cecropia spp.) (Watling and Orrock 2010). A greater availability of fleshy fruits and flowers (Oosterhoorn and Kappelle 2000; Galetti et al. 2003) may attract frugivorous and nectivorous birds and, consequently, facilitate higher provision of seed dispersal (Morante-Filho et al. 2018) and pollination services along these habitat interfaces. Apart from that, shorter distances between forest edges and pastures induce higher frugivore bird diversity (Pizo and dos Santos 2011), and this likely contributes to the higher levels of avian ESP at forest-pastures boundaries.
Bird assemblage responses to commercial tree plantations are relatively well investigated worldwide. Tropical tree plantations may improve landscape connectivity to forest birds (Zurita and Bellocq 2010; de Camargo Barbosa et al. 2017), and support diversified bird assemblages (Volpato et al. 2010), provided that plantations are older, lightly managed and include at least some native vegetation (Millan et al. 2015). However, contrary to some of these studies, our findings show low bird species richness within eucalyptus stands near forest remnants, with an average of only 9 species per site (amounting to 49 of the 200 species across all sites), and forest-specialists avoided this matrix type more frequently than we expected. Although we accounted for local habitat variation, this may occur because our eucalyptus stands were cultivated primarily for commercial exploitation, even-aged (4–6 years old), homogeneous and largely lacked native understorey vegetation. This is one of the most hostile scenarios for bird species richness within eucalyptus plantations (Millan et al. 2015).
Bird species composition in eucalyptus plantations is often a nested subset of species inhabiting quasi-natural forests (Wethered and Lawes 2005). In addition to the dearth of resources available for birds, simplified eucalyptus stands would at best provide redundant resources, which are already available in adjacent forests. This may explain the higher richness of forest-specialists restricted exclusively to forest sites adjacent to eucalyptus (92 species) compared to forests adjacent to pastures (68 species) (Fig. 3). A large number of forest-specialists was consistently found in forest patches than in neighbouring eucalyptus. However, eucalyptus plantations could still serve to enhance habitat connectivity for forest-specialists (de Camargo Barbosa et al. 2017), despite their low habitat value for most resident bird species.
Although our results on ESP by birds in matrix habitats are valid only for edges influenced by forest fragments, similar ESP estimates could be much lower than those reported here for large core areas of homogeneous pastures and eucalyptus stands. The lack of natural vegetation or stepping stones typical of extensive homogeneous matrix areas is hostile to diversified bird communities that could otherwise provide high ES interactions (Pizo and dos Santos 2011).
In general, habitat generalists were widely represented as potential ES providers in both pasture and eucalyptus patches (Table 2). Moreover, they were of comparable importance to forest-specialists at forest sites in terms of seed dispersal and pollination services (Table 2). Indeed, birds exhibiting wide habitat-breadth can be important seed dispersal agents, actively accelerating tropical forest regeneration (Carlo and Morales 2016), as well as important providers of pest control services in farmland near forest fragments (Boesing et al. 2017). Due to their relatively high abundance, ability to use a broad range of habitat types, and move through both natural and anthropogenic habitats (Table 1, Fig. 3), habitat generalists can play a key role in ecosystem service delivery across forest-matrix interfaces. Although seed dispersal services provided by forest-specialist birds cannot be ecologically compensated by habitat-generalist birds (Morante-Filho et al. 2018), the latter can still be helpful in providing SD services across highly disturbed landscapes. We therefore reinforce that special attention should be given to relatively ubiquitous habitat-generalist birds and other organisms, if the primary goal is to enhance ecosystem services in highly degraded landscapes (Pizo 2007).
Local and landscape effects on avian-mediated ESP
At forest edges, landscape-wide forest cover was the best predictor of SR and PC services by entire bird assemblages and of SR and all ESP by forest-affiliated species (Table 2, Fig. 4). This is consistent with the influence of landscape-wide forest cover on bird persistence (Banks-Leite et al. 2014). However, given that the vast majority of studies have not assessed the landscape structure affecting birds exclusively at forest edges, our results suggest that forest cover can similarly affect both core and edge habitats in birds, particularly in terms of the species richness and ecological functions performed by forest species.
Although landscape metrics related to land use heterogeneity (edge density and Shannon diversity index) were the best predictors of species richness and all ESP at eucalyptus plantations, the explanation power of our models was too low and not sufficiently informative (Table 2). In contrast, rural homesteads were the best predictor of SD and PO services by entire bird assemblages, and the best predictor of all ESP by habitat-generalists along forest edges. These results suggest that, beyond landscape heterogeneity per se, a higher density by rural homesteads throughout an otherwise uniform landscape can enhance the landscape connectivity for avian-mediated seed dispersal and pollination services. The presence of scattered trees and small woodlots, for example, likely offer important resources for many habitat generalist species, including thrushes (Turdus spp.), tanagers (Tangara spp.), hummingbirds (mainly Eupetomena macroura), and bananaquits (Coereba flaveola). These species are both ubiquitous and highly abundant in open-habitat areas but also serve important functions as seed and pollen vectors in human-modified landscapes (Pizo 2007).
Conclusions
We have shown that the erosion of bird species in highly altered neotropical landscapes and, consequently, the ecosystem services inherently flowing from their trophic interactions can be almost twice as severe across forest edges when they occur next to commercially exploited eucalyptus plantations than under comparable settings next to cattle pastures. The habitat complexity of pastures (e.g. presence of relict scattered trees) can also positively affect the potential provision of ecosystem services by birds, rather than surrounding landscape attributes. In addition, ESP by birds along forest edges was most affected by forest cover for forest-specialists, and nearby rural homesteads for habitat-generalists. The prevalence of such rural homesteads in agricultural landscapes may positively disrupt spatial homogenization across otherwise monotonous farmland matrix vegetation, thereby favouring infiltration of habitat-generalist birds that potentially provide pollination and seed dispersal services. This can apparently contribute to a higher forest regeneration capacity (i.e. higher spatial resilience) across highly disturbed and fragmented landscapes. Our results highlight not only the importance of matrix type, its habitat structure and landscape attributes in maintaining ES provision by birds along forest-matrix interfaces, but also the need for further studies quantifying the potential provision of ES in anthropogenic landscapes.
Given the high complexity of ecological processes operating within our study landscapes, we were unable to account for important features related to ESP, such as (1) the quality of species-specific interactions promoting the effective flow of ES, (2) any detrimental ecosystem disservices provided by some species (e.g. seed dispersal and pollination of invasive and alien species; predation on natural enemies of agricultural pests), and (3) the direct effects of bi-directional spillover patterns on ecosystem processes from forest to matrix areas and vice-versa (Blitzer et al. 2012). Despite such limitations, however, we believe that the metrics used here can be useful in assessing wider patterns of ESP by vertebrate and invertebrate communities at larger spatial scales (Eigenbrod et al. 2010), where more accurate in situ field measurements would be too labour-intensive and inefficient. A better understanding of the flows of ecosystem functions and services can inform management strategies that enhance the overall resilience of anthropogenic landscapes.
Finally, our data brings further evidence regarding conservation issues in land use management that augmenting (1) local habitat complexity within the wider habitat matrix, (2) landscape-scale forest cover, and (3) elements such as rural homesteads that disrupt the spatial homogenization typical of modern agricultural landscapes will likely enhance avian-mediated ESPs, which can ensure the regeneration capacity and multifunctionality of hyper-disturbed landscapes in the tropics.
References
Banks-Leite C, Pardini R, Tambosi LR, Pearse WD, Bueno AA, Bruscagin RT, Condez TH, Dixo M, Igari AT, Martensen AC, Metzger JP (2014) Using ecological thresholds to evaluate the costs and benefits of set-asides in a biodiversity hotspot. Science 80(345):1041–1045. https://doi.org/10.1126/science.1255768
Bennett AF, Radford JQ, Haslem A (2006) Properties of land mosaics: implications for nature conservation in agricultural environments. Biol Conserv 133:250–264. https://doi.org/10.1016/j.biocon.2006.06.008
Bibby CJ, Burgess ND, Hill DA, Mustoe SH (2002) Bird census techniques, 2nd edn. Academic Press, London, UK
Biz M, Cornelius C, Metzger JPW (2017) Matrix type affects movement behavior of a Neotropical understory forest bird. Perspect Ecol Conserv 15:10–17. https://doi.org/10.1016/j.pecon.2017.03.001
Blitzer EJ, Carsten FD, Holzschuh A, Klein AM, Rand TA, Tscharntke T (2012) Spillover of functionally important organisms between managed and natural habitats. Agric Ecosyst Environ 146:34–43. https://doi.org/10.1016/j.agee.2011.09.005
Boesing AL, Nichols E, Metzger JP (2017) Effects of landscape structure on avian-mediated insect pest control services: a review. Landscape Ecol. https://doi.org/10.1007/s10980-017-0503-1
Boesing AL, Nichols E, Metzger JP (2018) Land use type, forest cover and forest edges modulate avian cross-habitat spillover. J Appl Ecol 55:1252–1264. https://doi.org/10.1111/1365-2664.13032
Bovo AAA, Ferraz KMPMB, Magioli M, Alexandrino ER, Hasui E, Ribeiro MC, Tobias JA (2018) Habitat fragmentation narrows the distribution of avian functional traits associated with seed dispersal in tropical forest. Perspect Ecol Conserv 1:1. https://doi.org/10.1016/j.pecon.2018.03.004
Bregman TP, Lees AC, MacGregor HEA, Darski B, de Moura NG, Aleixo A, Barlow J, Tobias JA (2016) Using avian functional traits to assess the impact of land-cover change on ecosystem processes linked to resilience in tropical forests. Proc R Soc B Biol Sci. https://doi.org/10.1098/rspb.2016.1289
Brockerhoff EG, Jactel H, Parrotta JA, Ferraz SFB (2012) Role of eucalypt and other planted forests in biodiversity conservation and the provision of biodiversity-related ecosystem services. For Ecol Manag 301:43–50. https://doi.org/10.1016/j.foreco.2012.09.018
Carlo TA, Morales JM (2016) Generalist birds promote tropical forest regeneration and increase plant diversity via rare-biased seed dispersal. Ecology 97:1819–1831. https://doi.org/10.1890/15-2147.1
Carrara E, Arroyo-Rodríguez V, Vega-Rivera JH, Schondube JE, de Freitas SM, Fahrig L (2015) Impact of landscape composition and configuration on forest specialist and generalist bird species in the fragmented Lacandona rainforest, Mexico. Biol Conserv 184:117–126. https://doi.org/10.1016/j.biocon.2015.01.014
Crisci C, Ghattas B, Perera G (2012) A review of supervised machine learning algorithms and their applications to ecological data. Ecol Modell 240:113–122. https://doi.org/10.1016/j.ecolmodel.2012.03.001
Cutler DR, Edwards TC, Beard KH, Cutler A, Hess KT, Gibson J, Lawler JJ (2007) Random forests for classification in ecology. Ecology 88:2783–2792. https://doi.org/10.1890/07-0539.1
de Camargo Barbosa KV, Knogge C, Develey PF, Jenkins CN, Uezu A (2017) Use of small Atlantic Forest fragments by birds in Southeast Brazil. Perspect Ecol Conserv 15:42–46. https://doi.org/10.1016/j.pecon.2016.11.001
De Coster G, Banks-Leite C, Metzger JP (2015) Atlantic forest bird communities provide different but not fewer functions after habitat loss. Proc R Soc B Biol Sci 282:20142844. https://doi.org/10.1098/rspb.2014.2844
Eigenbrod F, Armsworth PR, Anderson BJ, Heinemeyer A, Gillings S, Roy DB, Thomas CD, Gaston KJ (2010) The impact of proxy-based methods on mapping the distribution of ecosystem services. J Appl Ecol 47:377–385. https://doi.org/10.1111/j.1365-2664.2010.01777.x
Fahrig L, Baudry J, Brotons L, Burel FG, Crist TO, Fuller RJ, Sirami C, Siriwardena GM, Martin JL (2011) Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecol Lett 14:101–112. https://doi.org/10.1111/j.1461-0248.2010.01559.x
Galetti M, Alves-Costa CP, Cazetta E (2003) Effects of forest fragmentation, anthropogenic edges and fruit colour on the consumption of ornithocoric fruits. Biol Conserv 111:269–273. https://doi.org/10.1016/S0006-3207(02)00299-9
Haddad NM, Brudvig LA, Clobert J, Davies KF, Gonzalez A, Holt RD, Lovejoy TE, Sexton JO, Austin MP, Collins CD, Cook WM, Damschen EI, Ewers RM, Foster BL, Jenkins CN, King AJ, Laurance WF, Levey DJ, Margules CR, Melbourne BA, Nicholls AO, Orrock JL, Song DX, Townshend JR (2015) Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci Adv 1:1–9. https://doi.org/10.1126/sciadv.1500052
Karp DS, Mendenhall CD, Sandí RF, Chaumont N, Ehrlich PR, Hadly EA, Daily GC (2013) Forest bolsters bird abundance, pest control and coffee yield. Ecol Lett 16:1339–1347. https://doi.org/10.1111/ele.12173
Kennedy CM, Zipkin EF, Marra PP (2016) Differential matrix use by Neotropical birds based on species traits and landscape condition. Ecol Appl 27:619–631. https://doi.org/10.1002/eap.1470
Lasky JR, Keitt TH (2012) The effect of spatial structure of pasture tree cover on avian frugivores in eastern Amazonia. Biotropica 44:489–497. https://doi.org/10.1111/j.1744-7429.2012.00857.x
Lee MB, Martin JA (2017) Avian species and functional diversity in agricultural landscapes: does landscape heterogeneity matter? PLoS ONE 12:1–21. https://doi.org/10.1371/journal.pone.0170540
Lindenmayer DB, Cunningham RB, MacGregor C, Crane M, Michael D, Fischer J, Montague-Drake R, Felton A, Manning A (2008) Temporal changes in vertebrates during landscape transformation: a large-scale “natural experiment”. Ecol Monogr 78:567–590. https://doi.org/10.1890/07-0945.1
Luck GW, Lavorel S, Mcintyre S, Lumb K (2012) Improving the application of vertebrate trait-based frameworks to the study of ecosystem services. J Anim Ecol 81:1065–1076. https://doi.org/10.1111/j.1365-2656.2012.01974.x
Maas B, Karp DS, Bumrungsri S, Darras K, Gonthier D, Huang JCC, Lindell CA, Maine JJ, Mestre L, Michel NL, Morrison EB, Perfecto I, Philpott SM, Sekercioglu CH, Silva RM, Taylor PJ, Tscharntke T, Van Bael SA, Whelan CJ, Williams-Guillén K (2016) Bird and bat predation services in tropical forests and agroforestry landscapes. Biol Rev 91:1081–1101. https://doi.org/10.1111/brv.12211
Mahood SP, Lees AC, Peres CA (2012) Amazonian countryside habitats provide limited avian conservation value. Biodivers Conserv 21:385–405. https://doi.org/10.1007/s10531-011-0188-8
McGarigal K, Cushman SA, Ene E (2012) FRAGSTATS v4: spatial pattern analysis program for categorical and continuous maps. University of Massachusetts, Amherst. http://www.umass.edu/landeco/research/fragstats/fragstats.html. Accessed 15 July 2016
Millan CH, Develey PF, Verdade LM (2015) Stand-level management practices increase occupancy by birds in exotic Eucalyptus plantations. For Ecol Manag 336:174–182. https://doi.org/10.1016/j.foreco.2014.10.005
Mitchell MGE, Bennett EM, Gonzalez A (2013) Linking landscape connectivity and ecosystem service provision: current knowledge and research gaps. Ecosystems 16:894–908. https://doi.org/10.1007/s10021-013-9647-2
Morante-Filho JC, Arroyo-Rodríguez V, de Souza Pessoa M, Cazetta E, Faria D (2018) Direct and cascading effects of landscape structure on tropical forest and non-forest frugivorous birds. Ecol Appl. https://doi.org/10.1002/eap.1791
Morante-Filho JC, Faria D, Mariano-Neto E, Rhodes J (2015) Birds in anthropogenic landscapes: the responses of ecological groups to forest loss in the Brazilian Atlantic forest. PLoS ONE 10:1–18. https://doi.org/10.1371/journal.pone.0128923
Morellato LPC, Haddad CFB (2000) Introduction: the Brazilian Atlantic Forest. Biotropica 32:786–792
Muñoz JC, Aerts R, Thijs KW, Stevenson PR, Muys B, Sekercioglu CH (2013) Contribution of woody habitat islands to the conservation of birds and their potential ecosystem services in an extensive Colombian rangeland. Agric Ecosyst Environ 173:13–19. https://doi.org/10.1016/j.agee.2013.04.006
Muylaert RL, Vancine MH, Bernardo R, Oshima JEF, Sobral-Souza T, Tonetti VR, Niebuhr BB, Ribeiro MC (2018) Uma nota sobre os limites territoriais da Mata Atlântica. Oecol Aust 22(3):302–311. https://doi.org/10.4257/oeco.2018.2203.09
Newbold T, Hudson LN, Hill SLL, Contu S, Lysenko I, Senior RA, Boerger L, Bennett DJ, Choimes A, Collen B, Day J, De Palma A, Diaz S, Echeverria-Londono S, Edgar MJ, Feldman A, Garon M, Harrison MLK, Alhusseini T, Ingram DJ, Itescu Y, Kattge J, Kemp V, Kirkpatrick L, Kleyer M, Correia DLP, Martin CD, Meiri S, Novosolov M, Pan Y, Phillips HRP, Purves DW, Robinson A, Simpson J, Tuck SL, Weiher E, White HJ, Ewers RM, Mace GM, Scharlemann JPW, Purvis A (2015) Global effects of land use on local terrestrial biodiversity. Nature 520:45–50. https://doi.org/10.1038/nature14324
Newbold T, Scharlemann JPW, Butchart SHM, Sekercioglu CH, Alkemade R, Booth H, Purves DW (2013) Ecological traits affect the response of tropical forest bird species to land-use intensity. Proc R Soc B Biol Sci 280:20122131. https://doi.org/10.1098/rspb.2012.2131
Oliveira AT, Fontes MAL (2000) Patterns of floristic differentiation among Atlantic Forests in southeastern Brazil and the influence of climate. Biotropica 32:793–810. https://doi.org/10.1111/j.1744-7429.2000.tb00619.x
Oosterhoorn M, Kappelle M (2000) Vegetation structure and composition along an interior-edge-exterior gradient in a Costa Rican montane cloud forest. For Ecol Manag 126:291–307. https://doi.org/10.1016/S0378-1127(99)00101-2
Pfeifer M, Lefebvre V, Peres CA, Wearn O, Marsh C, Banks-Leite C, Butchart S, Arroyo-Rodríguez V, Barlow J, Cerezo A, Cisneros L, D'Cruze N, Faria D, Hadley A, Klingbeil B, Kormann U, Lens L, Range GM, Morante-Filho JC, Olivier P, Peters S, Pidgeon A, Ribeiro D, Scherber C, Schneider-Maunoury L, Struebig M, Urbina-Cardona N, Watling JI, Willig M, Wood E, Ewers R (2017) Creation of forest edges has a global impact on forest vertebrates. Nature 551:187–191. https://doi.org/10.1038/nature24457
Pizo MA (2007) Frugivory by birds in degraded areas of Brazil. In: Dennis AJ, Schupp EW, Green RJ, Westcott DW (eds) Seed dispersal: theory and its application in a changing world. CABI Publishing, Wallingford, pp 615–627
Pizo MA, dos Santos BTP (2011) Frugivory, post-feeding flights of frugivorous birds and the movement of seeds in a Brazilian fragmented landscape. Biotropica 43:335–342. https://doi.org/10.1111/j.1744-7429.2010.00695.x
R Development Core Team (2016) R: a language and environment for statistical computing, R Foundation for Statistical Computing, Vienna
Ribeiro MC, Metzger JP, Martensen AC, Ponzoni FJ, Hirota MM (2009) The Brazilian Atlantic Forest: how much is left, and how is the remaining forest distributed? Implications for conservation. Biol Conserv 142:1141–1153. https://doi.org/10.1016/j.biocon.2009.02.021
Sekercioglu CH (2012) Bird functional diversity and ecosystem services in tropical forests, agroforests and agricultural areas. J Ornithol 153:153–161. https://doi.org/10.1007/s10336-012-0869-4
Sekercioglu CH, Wenny DG, Whelan CJ (eds) (2016) Why birds matter: Avian ecological function and ecosystem services. University of Chicago Press, Chicago
Taft OW, Haig SM (2005) The value of agricultural wetlands as invertebrate resources for wintering shorebirds. Agric Ecosyst Environ 110:249–256. https://doi.org/10.1016/j.agee.2005.04.012
Triviño M, Thuiller W, Cabeza M, Hickler T, Araújo MB (2011) The contribution of vegetation and landscape configuration for predicting environmental change impacts on iberian birds. PLoS ONE 6(12):e29373. https://doi.org/10.1371/journal.pone.0029373
Tscharntke T, Tylianakis JM, Rand TA, Didham RK, Fahrig L, Batáry P, Bengtsson J, Clough Y, Crist TO, Dormann CF, Ewers RM, Fründ J, Holt RD, Holzschuh A, Klein AM, Kleijn D, Kremen C, Landis DA, Laurance W, Lindenmayer D, Scherber C, Sodhi N, Steffan-Dewenter I, Thies C, van der Putten WH, Westphal C (2012) Landscape moderation of biodiversity patterns and processes—eight hypotheses. Biol Rev 87:661–685. https://doi.org/10.1111/j.1469-185X.2011.00216.x
Volpato GH, Prado VM, dos Anjos L (2010) What can tree plantations do for forest birds in fragmented forest landscapes? A case study in southern Brazil. For Ecol Manag 260:1156–1163. https://doi.org/10.1016/j.foreco.2010.07.006
Watling JI, Orrock JL (2010) Measuring edge contrast using biotic criteria helps define edge effects on the density of an invasive plant. Landscape Ecol 25:69–78. https://doi.org/10.1007/s10980-009-9416-y
Wenny DG, DeVault TL, Johnson MD, Kelly D, Sekercioglu CH, Tomback DF, Whelan CJ (2011) The need to quantify ecosystem services provided by birds. Auk 128:1–14. https://doi.org/10.1525/auk.2011.10248
Wethered R, Lawes MJ (2005) Nestedness of bird assemblages in fragmented Afromontane forest: the effect of plantation forestry in the matrix. Biol Conserv 123:125–137. https://doi.org/10.1016/j.biocon.2004.10.013
Wilman H, Belmaker J, Simpson J, de la Rosa C, Rivadeneira MM, Jetz W (2014) EltonTraits 1.0: Species-level foraging attributes of the world’s birds and mammals. Ecology 95(7):2027
Zurita GA, Bellocq MI (2010) Spatial patterns of bird community similarity: bird responses to landscape composition and configuration in the Atlantic forest. Landscape Ecol 25:147–158. https://doi.org/10.1007/s10980-009-9410-4
Acknowledgements
FMB is grateful to Brazil’s Ministry of Education (CAPES) for his Ph.D. studentship and FAPESP (Fundação de Amparo à Pesquisa do Estado de São Paulo, Grant Numbers 2013/19732-1, 2013/50421-2 and 2016/15376-4) for all financial support. Danilo Freire assisted with statistical analyses. Instituto Florestal do Estado de São Paulo granted a research permit (260108-009.965/2014) to work at Parque Estadual do Itaberaba. Field ornithologists Eduardo Alexandrino, Fabio Schunk and Carlos Gussoni participated in the expert survey. We are grateful to Júlia Assis and Instituto de Pesquisas Ecológicas (IPE) for logistic support during fieldwork. MAP is supported by a research Grant from the Brazilian Research Council (CNPq). MCR thanks FAPESP (Process #2013/50421-2), CNPq (Processes #312045/2013-1; #312292/2016-3) and PROCAD/CAPES (Project #88881.068425/2014-01) for their financial support. We are thankful for all the comments and suggestions by the anonymous reviewers.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Barros, F.M., Peres, C.A., Pizo, M.A. et al. Divergent flows of avian-mediated ecosystem services across forest-matrix interfaces in human-modified landscapes. Landscape Ecol 34, 879–894 (2019). https://doi.org/10.1007/s10980-019-00812-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-019-00812-z