
Abstract
Context
Biodiversity in tropical region has declined in the last decades, mainly due to forest conversion into agricultural areas. Consequently, species occupancy in these landscapes is strongly governed by environmental changes acting at multiple spatial scales.
Objectives
We investigated which environmental predictors best determines the occupancy probability of 68 bird species exhibiting different ecological traits in forest patches.
Methods.
We conducted point-count bird surveys in 40 forest sites of the Brazilian Atlantic forest. Using six variables related to landscape composition and configuration and local vegetation structure, we predicted the occupancy probability of each species accounting for imperfect detections.
Results
Landscape composition, especially forest cover, best predicted bird occupancy probability. Specifically, most bird species showed greater occupancy probability in sites inserted in more forested landscapes, while some species presented higher occurrence in patches surrounded by low-quality matrices. Conversely, only three species showed greater occupancy in landscapes with higher number of patches and dominated by forest edges. Also, several species exhibited greater occupancy in sites harbouring either larger trees or lower number of understory plants. Of uttermost importance, our study revealed that a minimum of 54% of forest cover is required to ensure high (> 60%) occupancy probability of forest species.
Conclusions
We highlighted that maintaining only 20% of native vegetation in private property according to Brazilian environmental law is insufficient to guarantee a greater occupancy for most bird species. We recommend that policy actions should safeguard existing forest remnants, expand restoration projects, and curb human-induced disturbances to minimise degradation within forest patches.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Tropical forests safeguard the greatest number of species on Earth, yet have been recently experiencing extensive losses in biodiversity due to the escalating increase of anthropogenic activities (Haddad et al. 2015; Edwards et al. 2019). In fact, nearly four million hectares of tropical forest were lost in the last decade, and converted to other land uses types, mainly for commodity production (i.e., agriculture), but also for cattle pastures and urban areas (Curtis et al. 2018). From fungus to large mammals, several studies have reported that deforestation and the subsequent land use modification drive a massive shift in community structure within forest remnants, with pervasive consequences for forest functioning (Benchimol et al. 2017; Brinkmann et al. 2019; Püttker et al. 2020). Although studies at the community level are important for unveiling patterns of species distribution within disturbed habitats, by considering all species homogenously, studies can overlook the specific responses, once species exhibit distinct sensitivities to environmental changes (Valente and Betts 2019). For instance, large-bodied species that occupy higher levels in the food chain, such as vertebrate predators, present sudden population decrease in highly deforested landscapes, enhancing their extinction probability (Paviolo et al. 2016). Therefore, assessing the idiosyncratic responses of individual species is vital to identify the pivotal predictors of species persistence in disturbed tropical landscapes, and consequently contribute for conservation measures in the Anthropocene.
Several studies have investigated the patterns of species occurrence in anthropogenic forest landscapes (Benchimol and Peres 2014; Bhakti et al. 2018; Thomas et al. 2019), but most of them focused on patch size and isolation due to their conceptual association with the widely recognised Island Biogeography Theory (MacArthur and Wilson 1967). Yet a key limitation of this approach is that forest patches in fragmented landscapes are usually surrounded by a mosaic of different land use types, which strongly differ from the homogeneous aquatic matrix of oceanic islands. Therefore, other characteristics related to landscape structure may be even more important to determine species occupancy in forest remnants than these commonly used patch-scale metrics, such as patch size and isolation (Bhakti et al. 2018). For instance, the landscape forest cover was a key predictor explaining mammalian occupancy in human-modified landscapes in Colombia (Boron et al. 2019), whereas the matrix structure strongly affected patterns of small mammals’ persistence in Amazonian forest fragments (Santos-Filho et al. 2012). However, local features are also likely to affect species occupancy within forest patches for a wide range of biological groups. In particular, bat (Thomas et al. 2019) and bird occupancy (Bhakti et al. 2018) substantially declined in forest fragments with increasing tree basal area and lower tree diameter, respectively. Therefore, a multi-scale approach is required to accurately assess the effects of environmental changes on the persistence of species in human-modified landscapes (Hill and Hamer 2004).
Birds comprise a valuable group to examine patterns of species persistence in anthropogenic landscapes. This is broadly considered a keystone group in tropical forests given their important role in providing ecological services including seed dispersal, pollination, and invertebrate control (Şekercioḡlu 2006). In the tropics, birds have been frequently used as good indicators of habitat quality, with the absence or presence of certain species indicating high degree of disturbance of a forest patch (Şekercioḡlu et al. 2002). More specifically, local extinctions from forest fragments can lead to a myriad of cascade effects, with the shrinkage of ecological interactions essential to the maintenance of forest structure and functionality. For instance, a recent study using tropical seed-dispersal networks indicated that 10% of simulated bird species loss results in almost 40% decline of long-distance seed dispersal (Donoso et al. 2020), with pervasive consequences for plant recruitment. However, bird species exhibit different sensitivities to habitat perturbation, with certain ecological groups most prone to be affected in highly fragmented landscapes (Pizo and Toneti 2020). In particular, forest-dweller birds, like forest understory insectivorous species and large-bodied frugivores are most prone to disappear, whereas species that forage and use forest edges are favoured in highly deforested landscapes (Morante-Filho et al. 2018a; Pizo and Toneti 2020).
Although a growing number of studies in tropical landscapes has evaluated the influence of variables operating at multi-scales on birds, most has focused on community attributes such as richness, abundance and species composition (Uezu et al. 2005; Banks-Leite et al. 2013; Morante-Filho et al. 2016). However, these studies neglect the species’ identity and consequently the specific effect of environmental changes on their persistence. Indeed, species responses to habitat disturbance can be shaped by life-history traits, with those species highly susceptible to disturbance mainly characterised by large body size, low dispersion capacity, high trophic level, and high levels of habitat specialisation (Ewers and Didham 2006; Newbold et al. 2013). Consequently, the synergy between distinct disturbances acting at multi-scales and individual species’ traits should module the bird occupancy in deforested landscapes. Additionally, most studies ignore variability in species detectability, potentially leading to biased or misleading results (Mackenzie et al. 2002). In fact, imperfect detectability is extremely common, as several bird species present cryptic behaviour, occur in low densities or rarely vocalise, features that decrease their chance of being registered even with long survey periods (Alldredge et al. 2007). Therefore, accounting for imperfect detectability is key to provide more reliable estimates of species occupancy and should be favoured in achieving applied conservation goals.
Here, we investigated which environmental predictors evaluated at multi-scales best predict the occupancy probability of a wide number of bird species in 40 forest sites located in the threatened Brazilian Atlantic forest. Specifically, we used six variables related to landscape composition (i.e., forest cover and matrix quality), landscape configuration (i.e., number of patches and edge density) and local vegetation structure (i.e., number of understory plants and mean tree diameter). Then, we evaluated the influence of these variables on the occupancy patterns accounting for imperfect detections for 68 species exhibiting different ecological traits, mainly related to habitat specialisation and diet type. From this analysis, we were able to identify which environmental predictor best determines the occurrence of bird species in forest patches, and assessed if life-history traits can help to explain the observed patterns. Based on the results, we further estimated the occupancy probability of those species greatly affected by forest cover in two distinct landscape scenarios (20% and 40% of forest cover amount) to evaluate the potential effectiveness of conservation measures based on the Brazilian environmental law (see Metzger et al. 2010) and empirical studies (see Arroyo-Rodríguez et al. 2020), and identified the minimum amount of forest cover required to ensure high local occupancy for these species. We thus discussed the most important features to be considered in conservation strategies to safeguard bird species with different life-history traits in human-modified landscapes.
Methods
Study area
Our study area is located in the southeastern Bahia State, Brazil (Fig. 1), in a region dominated by the Brazilian Atlantic forest—a priority area for biodiversity conservation due to its very high level of endemism and species richness, in addition to high rates of deforestation. The regional climate, according to Köppen classification, is hot and humid without a dry season. Mean annual temperature is 24 °C, and annual rainfall averages 2,000 mm/yr, without a significant seasonal climatic variation (Thomas et al. 1998).

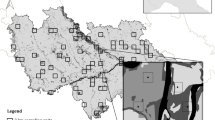
Sampled forest sites (red points) located in the southeastern Bahia State, Brazil. We show the land cover types within a 600 m radius from the centre of each site, and the environmental predictors calculated at landscape and local scales. Forest patches were highlighted in green, and cocoa agroforests, rubber tree and Eucalyptus sp. plantations and cattle pastures were represented by red, yellow, orange and grey colour within each landscape, respectively
We used high-resolution satellite images (i.e., Quick Bird and World View, both from 2011, and Rapid Eye from 2009–2010; with resolutions of 0.6 m, 0.5 m and 5 m, respectively) to obtain recent cloudless images for the study region. After image classification and digitalization at the scale of 1:10,000 using ArcGIS software, we developed a digital map that covered 3500 km2. We first identified 58 sites within forest remnants located at least 1 km apart from each other and > 150 m from the nearest forest edge. We thus excluded forest sites with unfeasible access, inserted in indigenous lands or highly mountainous forest. We then randomly selected 40 sites along a gradient of forest cover amount (Fig. 1), all exhibiting similar soil, topography, and floristic characteristics (Benchimol et al. 2017). All sites were located in forest patches isolated from each other by an anthropogenic matrix dominated by shade cacao plantations, rubber tree and Eucalyptus plantations, and/or cattle pastures.
Environmental predictors
We estimated four metrics related to landscapes structure within a 600-m radius (115-ha landscape) from each forest site (Appendix S1 in Supplementary Material). This landscape size can be considered biologically important for birds because included the home range of most Neotropical forest species (Robinson et al. 2000; Boscolo and Metzger 2009; Kennedy et al. 2017). Using this buffer size, we also avoided spatial overlapping among forest sites and ensured a large variation in several explanatory variables, which are necessary characteristics to provide accurate landscape-scale inferences (Eigenbrod et al. 2011). Finally, previous multi-scale analysis performed in several studies using this same dataset detected that 600-m radius was the most adequate spatial scale for predicting taxonomic (Morante-Filho et al. 2016), phylogenetic (Morante-Filho et al. 2018b) and functional (Matuoka et al. 2020) diversity of birds.
We used software FRAGSTATS© (McGarigal et al. 2012) to estimate two metrics related to (i) landscape composition—the percentage of old-growth and secondary forest (forest cover, hereafter), and an index of matrix quality; and two others related to (ii) landscape configuration—edge density and number of forest patches. To estimate matrix quality, we calculated an index that relates the percentage of each land-cover type within the matrix in relation to its relative quality. Because species presence declines along a land-use intensification (Fahrig 2003), the relative quality was ranked based on the vegetation structure of each land-cover type on a six-point scale (Garmendia et al. 2013): 1 (lowest quality, including water bodies, roads and human settlements), 2 (cattle pastures), 3 (croplands), 4 (Eucalyptus sp. plantations), 5 (rubber tree plantations) and 6 (shade-cocoa plantations). This ranking considers the positive effect of lower contrast between land use types when compared to forest, which could be able to provide additional habitat supplementary. Using this ranking, the index of quality matrix was calculated as: [(1 × % water, roads, human settlements and mangroves) + (2 × % cattle pastures) + (3 × % croplands) + (4 × % Eucalyptus sp. plantations) + (5 × % rubber tree plantations) + (6 × % shade-cocoa plantations)]/6.
We also characterized the vegetation structure in four 20 × 4-m forest plots, randomly located within each forest site, but maintaining a minimum inter-plot distance of 150 m. From January 2013 to April 2014, we recorded all woody plants in the understory (50–200 cm in height), all trees above the understory layer (i.e., with a diameter at breast height, DBH ≥ 5 cm), and the mean DBH of the trees in each plot. These three variables were summarized in a vegetation complexity index using a Principal Component Analysis (PCA), in which axis 1 from PCA explained 53% of the variation in vegetation structure (Appendix S2).
Bird surveys
In each site, we sampled bird species using point counts during three field campaigns: January–April 2013, May–September 2013 and October 2013–April 2014, which included the bird breeding season (September to January). In each campaign, all sites were visited twice—one in the morning (06:00 to 09:00 hs) and another in the afternoon (15:00 to 17:00 hs), with the order of sampling points being randomly chose to reduce temporal bias. On each site, we established four sampling points (50-m radius each, located 150–450 m apart from each other) to conduct 15 min of bird sampling through visual (8 × 42 binoculars) and auditory records. Total sampling effort per forest site was 6 h, distributed in six independent visits. We finally classified each bird species according to their habitat specialisation (i.e. forest dependent or non-forest dependent species) and trophic guild (i.e., insectivores, frugivores, nectivores and omnivores). For more details, see Table S2 in Appendix S3.
Statistical analyses
We firstly performed a Spearman's correlation analysis between all environmental predictors, and then we excluded both the number of trees and Axis 1 of PCA from further analysis, given its highly correlation (r ≥ 0.70) with other variables (Appendix S4).
We thus constructed matrices of detection (1) and non-detections (0) for each species in all 40 sites, combining data from all four sampling points to represent each visit on each surveyed site. We then used single-season occupancy models (MacKenzie et al. 2002) to estimate probabilities of occupancy (Ψ) and detection (p) for each species recorded in at least 25% of sites by using the software PRESENCE© (Hines 2006). This analytic approach explicitly accounts for failures in species detection, providing occupancy estimates that consider the probability of not recording the species in a site where it actually occurs. For this, we defined a set of simple models that might explain the site occupancy and the detection of bird species. Specifically, we included our landscape and local variables individually to model forest site occupancy, in addition to a single full model containing all these six potential predictors. Additionally, we developed two models of detection probability to control the sampling effect: (1) constant detection probability and (2) detection probabilities affected by AREA, which considers the total area of the surveyed patch within the buffer, given that the chance of recording a species might be reduced in larger forest sites. We also tested the null model, which assumes constant species presence and detection probabilities across time and forest sites. We thus used Akaike’s Information Criterion (AIC) to rank models and calculate Akaike weights (Burnham and Anderson 2002). When more than one model showed ∆AIC ≤ 2.00, we considered the best one as the simplest model (i.e., containing lower number of parameters).
Based in our best models showing the influence of forest cover on the estimates of occurrence in forest patches, we then estimated the occupancy probability for each species in two distinct scenarios of amount of forest cover at the landscape-scale: (i) 20% forest cover, based on the current Brazilian environmental law, which requires that private rural properties within the Atlantic forest domain protect a minimum of 20% of their total area (Metzger et al. 2010; Soares-Filho et al. 2014); and (ii) 40% forest cover, considering that a recent study, which combined theoretical concepts with empirical support, suggested that maintaining 40% of landscape habitat cover is the minimum amount required to safeguard biodiversity (see Arroyo-Rodríguez et al. 2020). We further estimated the minimum amount of forest cover required to ensure local occurrence for each species, considering a threshold of occupancy probability of 60%. We opted to use a conservative threshold (> 50%) to enhance the probability of ensuring species occurrence (Benchimol and Peres 2015). All graphs were carried out in R software (R Development Core Team 2019) using ggplot2 package.
Results
We observed 184 bird species considering all 40 forest sites, although only 68 species were recorded in at least 25% of sites. Of those, 42 species (62%) were forest birds, and 26 species (38%) were non-forest birds (Appendix S3). The most common forest species were Patagioenas speciosa, Phaethornis ruber and Tolmomyias flaviventris, recorded in 37, 35 and 33 forest sites, respectively. In addition, Megaryncgus pitangua, Euphonia violacea and Tangara palmarum, all non-forest species, were recorded, respectively, in 36, 35, and 35 sites. Also, insectivorous birds were the trophic guild most commonly recorded (32 species), followed by frugivores (26 species), and omnivores (8 species) (see Appendix S3).
Considering all 68 species, we did not obtain the occupancy estimate for only 5 species because the variance–covariance matrix could not be calculated successfully (Appendix S5). We observed that the mean observed occupancy probability (0.52 ± 0.19) was significantly (t = 3.80, df = 122.4, P = 0.0002) lower than the estimated probability (0.64 ± 0.19), which suggests that accounting for imperfect detectability enabled to decrease the number of false absences, providing more reliable occupancy values. Our results also indicated that the null model was included amongst the most parsimonious model (ΔAIC ≤ 2.00) for 23 species (Appendix S5), and we therefore considered that no other model best explained the pattern than chance. Hence, 38 species were affected by at least a single environmental predictor (see Fig. 2) and for only two species (Myiarchus ferox and Tangara cayana) the best model was composed by all variables (Appendix S5).

Panel plots for each bird species showing the influence of the best environmental predictor on the occupancy probability in forest patches (N = 38). Panels are ordered following composition landscape—i.e., amount of forest cover (n = 19 species) and quality matrix index (n = 6 species), configuration landscape—i.e., edge density (n = 2 species) and number of forest patches (n = 1 species), and local vegetation structure—i.e., mean tree diameter at breast height (n = 4 species) and number of understory plants (n = 6 species). Green and blue lines represent forest and non-forest species, respectively. The main trophic guild is shown by the illustration in each panel, which fruit, insect, flower and all together symbols indicate frugivores, insectivores, nectarivores and omnivores, respectively. Scientific names of each species is inserted above of panels. Bird pictures resource: https://www.hbw.com
In general, the occupancy probability of birds was mostly affected by landscape composition metrics. In fact, forest cover comprised the best predictor explaining occupancy patterns, especially for forest-dweller birds (n = 15 species; Appendix S5) that showed greater occurrence in sites inserted in more forested landscapes (Fig. 2). Yet, forest species responded differently to this forest cover gradient, with Glyphorynchus spirurus and Rhytipterna simplex requiring a minimum of 30% of forest cover to exhibit a great probability (i.e. ≥ 80%) to occur, whereas the majority of species demanding more than 60% of forest cover, including Herpsilochmus pileatus, Myrmotherula urostica, and Ceratopipra rubrocapilla. Conversely, non-forest species did not follow a general pattern following forest cover amount—two species (Camptostoma obsoletum and Elaenia flavogaster) substantially decreased in sites located in more forested landscapes, in contrast to Coereba flaveola and Saltator maximus, which exhibited greater occupancy estimates in these landscapes. Additionally, six species were affected by matrix quality—mostly those showing higher occurrence probability in forest patches surrounding by matrices of low quality (Fig. 2).
Our findings evidenced that only three species were influenced by landscape configuration (Fig. 2). In particular, these bird species showed greater occupancy probability in forest patches inserted in more fragmented landscapes; i.e. with higher number of patches and dominated by forest edges. Finally, local variables were important predictors of occupancy probability for ten species, with mean DBH and the number of understory plants appearing in the best model for three and seven species, respectively (Fig. 2). For instance, Ramphastos vitellinus and Tangara cyanomelas, both forest frugivorous birds, showed greater occupancy probability in forest patches harbouring larger trees (Fig. 2). Also, Ramphocaenus melanurus, Sittasomus griseicapillus and Thamnophilus ambiguus, all forest insectivorous birds, showed greater occupancy probability in sites containing more understory plants, in contrast to non-forest frugivorous birds, Nemosia pileata and Tangara cayana, which showed an opposite pattern (Fig. 2).
Our results also showed that most forest species have a low probability of occupying sites inserted in deforested landscapes (i.e. 20% of forest cover, Table 1). In addition, we observed a slight increase of occupancy probability of forest birds in more forested landscapes (i.e. 40% of forest cover). In contrast, all non-forest species presented an opposite occurrence pattern, indicating high occupancy in all predicted scenarios (Table 1). We also observed that, on average, 54% was the lowest estimated forest cover at the landscape required to ensure a > 60% occupancy probability of forest species (Table 1). However, this value varied among the species evaluated. For instance, Crypturellus soui—forest frugivorous species—showed a > 60% occupancy probability in landscapes with at least 70% of forest cover, while Rhytipterna simplex—forest insectivorous birds—required only 20% of forest cover (Table 1). In contrast, non-forest species exhibited high occupancy rates (> 60%) in deforested landscapes. Indeed, non-forest birds required, on average, only 31% forest cover at the landscape-scale. For instance, our results evidenced that 12.9% is the minimum forest cover at the landscape required to ensure a 60% occupancy probability of Saltator maximus—a non-forest omnivorous birds (Table 1).
Discussion
Our study clearly shows the greater importance of landscape composition over landscape configuration and local vegetation structure in explaining occupancy patterns of birds in forest patches of the threatened Brazilian Atlantic forest. Considering those species influenced by any environmental predictor, we unveil that 63% of birds had their occupancy probability best explained by landscape composition. Specifically, forest cover was the best predictor for nearly half of the evaluated species, although we detected a great variation in the minimum amount required to ensure greater occupancy of these species. By contrast, landscape configuration weakly explained occupancy patterns, with only two species being positively influenced by edge density and a single species by number of patches. Also, local variables exerted a strong influence for a quarter of species, suggesting that more conserved patches (i.e., exhibiting larger trees and lower number of understory plants) are prone to guarantee the incidence of species. Finally, we observed that the current legislation for the Brazilian Atlantic forest is unlikely to ensure bird occupancy in forest patches. As discussed below, it is primordial that studies consider environmental predictors acting at multi-scales to best understand patterns of species occupancy in fragmented landscapes and hence provide valuable information for bird conservation.
Determinants of bird occupancy in forest patches
Our findings highlighted that forest cover at the landscape was the key predictor for forest birds’ occupancy in forest patches. In fact, forest cover has been widely recognized to be a pivotal driver of biodiversity patterns in fragmented landscapes, especially because this metric is positively related to habitat amount and landscape connectivity for a wide range of species, in particular forest-dweller species (Fahrig 2003). Similar result was observed in a study performed in the south-eastern Australia, which indicated that forest amount was almost six times more important for bird occupancy in woodland patches than landscape configuration (Lindenmayer et al. 2020). Yet, we observed great variation in species’ responses to forest cover. Considering all species, most birds present low probability (< 30%) of occurring in forest patches inserted in highly deforested landscapes (see Appendix S6); however, distinct patterns are unveiled when assessing individual responses. For instance, the insectivore G. spirurus and the frugivore C. rubrocapilla, both forest species, showed about 80% and 20% occupancy probability, respectively, in landscapes with at least 30% of forest cover. Other species, mainly non-forest birds, presented pronounced occurrence (> 80%) in deforested landscapes. In particular, these non-forest species are better adapted to disturbed habitats (Bregman et al. 2014), and present high vagility, which allows them to explore different land cover types across the landscape (Julliard et al. 2006). For instance, E. flavogaster—a non-forest omnivorous species that showed high occupancy probability in deforested landscapes—is commonly recorded foraging in urban green areas, agricultural areas and forest edges (Sick 1984).
The matrix quality also explained occupancy patterns for six bird species, with species greatly varying in their specific responses. It is well-know that matrix type can drive colonization and extinction dynamics of birds in fragmented forest landscapes (Kennedy et al. 2017), with some sensitive species likely to occur in unsuitable fragments if the surrounding matrix offers supplementary resources (Antongiovanni and Metzger 2005). However, we unveil that only one forest-dweller species increased the occurrence probability in fragments surrounded by matrices with a greater proportion of rubber tree and shade-cocoa plantations (i.e., greater quality), the understory insectivores Myrmotherula axillaris. Conversely, the other species were less prone to be found in patches inserted in more friendly matrices. Several birds are often seen foraging in agricultural, urban and/or forest edges (Sick 1984; Morante-Filho et al. 2018a), indicating they are greatly favoured by the land use intensification in human-disturbed areas.
Our results provide strong support that landscape configuration poorly predicted the incidence of studied birds. In fact, only two species were substantially affected by edge density. Considering that some generalist species show a greater preference for foraging and breeding on fragment boundaries (Ewers and Didham 2006), we expected that most non-forest species would have their occupancy probability increased in landscapes exhibiting greater edge density. This was the case of Turdus leucomelas, in which a telemetry study demonstrated that this thrush species is indeed favoured in sites exhibiting greater edge amount (Da Silveira et al. 2016). Corroborating other studies (Pavlacky et al. 2012; Lindenmayer et al. 2020), our additional metric of landscape configuration (i.e. number of patches) was an unimportant predictor, affecting solely a unique species. This metric is strongly related to habitat fragmentation, given that greater number of patches within a landscape is expect to create subpopulations, thus favouring both metapopulation dynamics and species persistence in highly fragmented landscapes (Hanski 1999). Two plausible hypotheses can explain the observed weak effect of landscape configuration in our study. Firstly, the configuration effects (e.g. increasing in forest edges and patches within the landscape) can be minimized due to the high mobility exhibited by most bird species, enabling their use of different patches scattered in the landscape (Lindenmayer et al. 2020). Secondly, we observed a small range on the number of patches across studied landscapes (Appendix S1), which may lead to less variation than what would be required to show detectable responses in bird species (Lindenmayer et al. 2020).
We also detected that local vegetation structure significantly explained patterns of occurrence for several forest bird species. Particularly, forest sites containing larger trees were prone to ensure greater probability for R. vitellinus and T. cyanomelas, both frugivorous species. These sites likely exhibit greater fruit availability, given that large trees usually produce key resources for faunal species (Laurance et al. 2000). Also, big trees usually present other key resources for birds, such as more suitable cavities for nesting and sheltering, either created by excavators or naturally present in live trees, which are frequently used by a wide array of bird species (Lindenmayer et al. 2012; Bhakti et al. 2018). Indeed, forest patches retaining larger trees would favour both the foraging and breeding individuals of R. vitellinus, a large-bodied species (360 g) that depends on fleshy fruits and cavities for breeding (Sick 1984). Additionally, the number of understory plants also affected several bird species, yet patterns substantially distinct were observed when general or individual responses are evaluated (see Appendix S6 and Fig. 2). For forest species, individual responses can be explained by its dietary specificity. For instance, while Cacicus haemorrhous and Dendrocincla turdina were more prone to occur in forest sites containing lower number of understory plants, three insectivorous forest birds (Ramphocaenus melanurus, Sittasomus griseicapillus, Thamnophilus ambiguous) enhanced their probability of occurrence in sites containing more understory plants. In fact, disturbed forests commonly present dense vegetation mainly composed by pioneer plants, which can favour generalist insects (Guimarães et al. 2014), therefore providing greater resource availability for those insectivorous birds. Additionally, variables at local scale also predicted occupancy patterns for some non-forest birds. Three recorded non-forest frugivorous, Dacnis cayana, N. pileata and T. cayana, showed greater incidence in more disturbed sites (i.e., presenting either smaller trees or lower number of understory plants). Indeed, these species are frequently recorded in urban and open areas (Pena et al. 2017). Based on our results, we provide further evidence that, as the occurrence probability of certain species is highly sensitive to local disturbance, management actions at the landscape scale can be insufficient to ensure their occupancy in human-modified landscapes.
The minimum forest amount for bird occupancy
According to the Brazilian environmental law, which determines that private rural properties within the Atlantic forest domain should protect only 20% of their total area (Metzger et al. 2010; Soares-Filho et al. 2014), our results showed that most studied forest species exhibit a weak probability of occupying sites inserted in such highly deforested landscapes. For instance, H. pileatus and M. urosticta—both forest insectivorous birds and endemic species of Brazilian Atlantic forest, presented low occupancy probability, 15% and 9%, respectively, in landscapes with only 20% forest cover. Even if we consider that 40% is the minimum habitat amount in tropical landscapes required to maintain biodiversity (as suggested by Arroyo-Rodríguez et al. 2020), only a slight increase of occupancy probability for most forest birds is observed in our landscapes. This is an alarming finding especially because to ensure > 60% occupancy probability of forest birds in forest patches it would be required almost three-fold landscape forest cover than suggested by the current Brazilian environmental law (Soares-Filho et al. 2014). To aggravate this scenario, Brazil has been facing a serious environmental crisis that has contributed to the increase in deforestation in several Brazilian biomes (Escobar 2020). Specifically, in the 2018–2019 period, the Atlantic forest has lost 14,500 ha, 27% more than the previous period (i.e., 2017–2018; SOS Mata Atlântica and INPE 2020). Considering that 53% of native vegetation occurs in private properties in Brazil (Soares-Filho et al. 2014), our results underline the importance to discuss the current laws and elaborate more effective environmental legislation aiming to preserve wildlife species.
Conclusions
Our study revealed that environmental factors operating at multiple spatial scales are critical to determine the occupancy of bird species in forest patches inserted in human-modified landscapes. In particular, occupancy probability of most bird species was strongly determined by landscape composition, primarily the forest amount at the landscape. Therefore, bird conservation efforts towards the Atlantic forest should not only safeguard the existing forest remnants, but also require to expand forest cover via restoration projects within severely deforested landscapes. Additionally, measures for mitigating the local structure degradation within forest patches are also required to safeguard bird species, given that local vegetation features played a key role for several species. Indeed, human-induced disturbances frequently occur in tropical forests, including acute (i.e. selective logging, wildfires) and chronic (i.e. firewood and non-timber exploitation) disturbances, inducing to marked changes in forest structure (Barlow et al. 2016) with severe consequences for fauna. Specifically, the demise of bird ecological groups in disturbed patches, such as large frugivorous birds and understory insectivorous species, can disrupt ecological roles performed by species, including seed dispersion and arthropod control, which are vital to the forest functionality in the long term (Şekercioḡlu 2006). Therefore, we recommend that policy actions go further than preventing deforestation, and should also focus on minimising within-forest disturbance by engaging many stakeholders such as government, land farmers and institutions.
References
Alldredge MW, Simons TR, Pollock KH (2007) Factors affecting aural detections of songbirds. Ecol Appl 17(3):948–955
Antongiovanni M, Metzger JP (2005) Influence of matrix habitats on the occurrence of insectivorous bird species in Amazonian forest fragments. Biol Conserv 122(3):441–451
Arroyo-Rodríguez V, Fahrig L, Tabarelli M, Watling JI, Tischendorf L, Benchimol M, Melo FPL, Morante-Filho JC, Santos BA, Arasa-Gisbert R, Arce-Peña N, Cervantes-López MJ, Cudney-Valenzuela S, Galan-Acedo C, San-José M, Vieira IMG, Slik JWF, Nowakowski J, Tscharntke T (2020) Designing optimal human-modified landscapes for forest biodiversity conservation. Ecol Lett 23:1404–1420
Banks-Leite C, Ewers RM, Metzger JP (2013) The confounded effects of habitat disturbance at the local, patch and landscape scale on understorey birds of the Atlantic Forest: implications for the development of landscape-based indicators. Ecol indic 31:82–88
Barlow J, Lennox GD, Ferreira J, Berenguer E, Lees AC, Nally RM, Thomson JR, Ferraz SFB, Louzada J, Oliveira VHF, Parry L, Solar RRC, Vieira ICG, Aragão LEOC, Begotti RA, Braga RF, Cardoso TM, Oliveira RC Jr, Souza CM Jr, Moura NG, Nunes SS, Siqueira JV, Pardini R, Silveira JM, Vaz-de-Mello FZ, Veiga RCS, Venturieri A, Gardner TA (2016) Anthropogenic disturbance in tropical forests can double biodiversity loss from deforestation. Nature 535(7610):144–147
Benchimol M, Peres CA (2014) Predicting primate local extinctions within “real-world” forest fragments: a pan-neotropical analysis. Am J Primatol 76(3):289–302
Benchimol M, Peres CA (2015) Predicting local extinctions of Amazonian vertebrates in forest islands created by a mega dam. Biol Conserv 187:61–72
Benchimol M, Mariano-Neto E, Faria D, Rocha-Santos L, Pessoa MS, Gomes FS, Custodio D, Cazetta E (2017) Translating plant community responses to habitat loss into conservation practices: forest cover matters. Biol Conserv 209:499–507
Bhakti T, Goulart F, de Azevedo CS, Antonini Y (2018) Does scale matter? The influence of three-level spatial scales on forest bird occurrence in a tropical landscape. PLoS ONE 13(6):e0198732
Boron V, Deere NJ, Xofis P, Link A, Quiñones-Guerrero A, Payan E, Tzanopoulos J (2019) Richness, diversity, and factors influencing occupancy of mammal communities across human-modified landscapes in Colombia. Biol Conserv 232:108–116
Boscolo D, Metzger JP (2009) Is bird incidence in Atlantic forest fragments influenced by landscape patterns at multiple scales? Landsc Ecol 24(7):907–918
Bregman TP, Şekercioḡlu CH, Tobias JA (2014) Global patterns and predictors of bird species responses to forest fragmentation: implications for ecosystem function and conservation. Biol Conserv 169:372–383
Brinkmann N, Schneider D, Sahner J, Ballauff J, Edy N, Barus H, Bambang I, Budi SW, Qaim M, Daniel R, Polle A (2019) Intensive tropical land use massively shifts soil fungal communities. Sci Rep 9(1):1–11
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach. Springer, New York
Curtis PG, Slay CM, Harris NL, Tyukavina A, Hansen MC (2018) Classifying drivers of global forest loss. Science 361(6407):1108–1111
Da Silveira NS, Niebuhr BBS, de Lara MR, Ribeiro MC, Pizo MA (2016) Effects of land cover on the movement of frugivorous birds in a heterogeneous landscape. PLoS ONE 11(6):e0156688
Donoso I, Sorensen MC, Blendinger PG, Kissling WD, Neuschulz EL, Mueller T, Schleuning M (2020) Downsizing of animal communities triggers stronger functional than structural decay in seed-dispersal networks. Nat Commun 11(1):1–8
Edwards DP, Socolar JB, Mills SC, Burivalova Z, Koh LP, Wilcove DS (2019) Conservation of tropical forests in the anthropocene. Curr Biol 29(19):1008–1020
Eigenbrod F, Hecnar SJ, Fahrig L (2011) Sub-optimal study design has major impacts on landscape-scale inference. Biol Conserv 144:298–305
Escobar H (2020) Deforestation in the Brazilian Amazon is still rising sharply. Science 369:613–613
Ewers RM, Didham RK (2006) Confounding factors in the detection of species responses to habitat fragmentation. Biol Rev 81(1):117–142
Fahrig L (2003) Effects of habitat fragmentation on biodiversity. Annu Rev Ecol Evol Syst 34:487–515
Garmendia A, Arroyo-Rodríguez V, Estrada A, Naranjo EJ, Stoner KE (2013) Landscape and patch attributes impacting medium-and large-sized terrestrial mammals in a fragmented rain forest. J Trop Ecol 29(4):331–344
Guimarães CDC, Viana JPR, Cornelissen T (2014) A meta-analysis of the effects of fragmentation on herbivorous insects. Environ Entomol 43:537–545
Haddad NM, Brudvig LA, Clobert J, Davies KF, Gonzalez A, Holt RD, Lovejoy TE, Sexton JO, Austin MP, Collins CD, Cook WM, Damschen EI, Ewers RM, Foster NL, Jenkins CN, King AJ, Laurance WF, Levey DJ, Margules CR, Melbourne BA, Nicholls AO, Orrock JL, Song D-X, Townshend JR (2015) Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci Adv 1(2):e1500052
Hanski I (1999) Habitat connectivity, habitat continuity, and metapopulations in dynamic landscapes. Oikos 87:209–219
Hill JK, Hamer KC (2004) Determining impacts of habitat modification on diversity of tropical forest fauna: the importance of spatial scale. J Appl Ecol 41(4):744–754
Hines JE (2006) PRESENCE. Page Software to estimate patch occupancy and related parameters, U.S. Patuxent Wildlife Research Center, Maryland
Julliard R, Clavel J, Devictor V, Jiguet F, Couvet D (2006) Spatial segregation of specialists and generalists in bird communities. Ecol Lett 9:1237–1244
Kennedy CM, Zipkin EF, Marra PP (2017) Differential matrix use by Neotropical birds based on species traits and landscape condition. Ecol Appl 27(2):619–631
Laurance WF, Delamônica P, Laurance SG, Vasconcelos HL, Lovejoy TE (2000) Rainforest fragmentation kills big trees. Nature 404(6780):836–836
Lindenmayer DB, Blanchard W, McBurney L, Blair D, Banks S, Likens GE, Franklin JF, Laurance WF, Stein JAR, Gibbons P (2012) Interacting factors driving a major loss of large trees with cavities in a forest ecosystem. PLoS ONE 7(10):e41864
Lindenmayer DB, Blanchard W, Foster CN, Scheele BC, Westgate MJ, Stein J, Crane M, Florance D (2020) Habitat amount versus connectivity: an empirical study of bird responses. Biol Conserv 241:108377
MacArthur RG, Wilson EO (1967) The theory of island biogeography. Princeton University Press, Princeton
McGarigal K, Cushman SA, Ene E (2012) FRAGSTATS v4: spatial pattern analysis program for categorical and continuous maps. Computer software program. University of Massachusetts, Amherst
MacKenzie DI, Nichols JD, Lachman GB, Droege S, Royle JA, Langtimm CA (2002) Estimating site occupancy rates when detection probabilities are less than one. Ecology 83(8):2248–2255
Matuoka MA, Benchimol M, Morante-Filho JC (2020) Tropical forest loss drives divergent patterns in functional diversity of forest and non-forest birds. Biotropica 52:738–748
Metzger JP, Lewinsohn T, Joly CA, Verdade LM, Rodrigues RR (2010) Brazilian law: full speed in reverse. Science 329:276–277
Morante-Filho JC, Arroyo-Rodríguez V, Faria D (2016) Patterns and predictors of β-diversity in the fragmented Brazilian Atlantic forest: a multiscale analysis of forest specialist and generalist birds. J Anim Ecol 85(1):240–250
Morante-Filho JC, Arroyo-Rodríguez V, Pessoa MDS, Cazetta E, Faria D (2018a) Direct and cascading effects of landscape structure on tropical forest and non-forest frugivorous birds. Ecol Appl 28(8):2024–2032
Morante-Filho JC, Arroyo-Rodríguez V, de Andrade ER, Santos BA, Cazetta E, Faria D (2018b) Compensatory dynamics maintain bird phylogenetic diversity in fragmented tropical landscapes. J Appl Ecol 55(1):256–266
Newbold T, Scharlemann JP, Butchart SH, Şekercioğlu ÇH, Alkemade R, Booth H, Purves DW (2013) Ecological traits affect the response of tropical forest bird species to land-use intensity. Proc R Soc B 280(1750):20122131
Paviolo A, De Angelo C, Ferraz KM, Morato RG, Pardo JM, Srbek-Araujo AC, Beisiegel BM, Lima F, Sana D, Silva MX, Velázquez MC, Cullen L, Crawshaw P Jr, Jorge MLSP, Galetti PM, Di Bitetti MS, Paula RC, Eizirik E, Aide TM, Cruz P, Perilli MLL, Souza ASMC, Quiroga V, Nakano E, Pinto FR, Fernández S, Costa S, Moraes EA Jr, Azevedo F (2016) A biodiversity hotspot losing its top predator: the challenge of jaguar conservation in the Atlantic Forest of South America. Sci Rep 6(1):1–16
Pavlacky DC, Possingham HP, Lowe AJ, Prentis PJ, Green DJ, Goldizen AW (2012) Anthropogenic landscape change promotes asymmetric dispersal and limits regional patch occupancy in a spatially structured bird population. J Anim Ecol 81(5):940–952
Pena JC, Martello F, Ribeiro MC, Armitage RA, Young RJ, Rodrigues M (2017) Street trees reduce the negative effects of urbanization on birds. PLoS ONE 12(3):e0174484
Pizo MA, Tonetti VR (2020) Living in a fragmented world: birds in the Atlantic forest. Condor. https://doi.org/10.1093/condor/duaa023
Püttker T, Crouzeilles R, Almeida-Gomes M, Schmoeller M, Maurenza D, Alves-Pinto H, Metzger JP (2020) Indirect effects of habitat loss via habitat fragmentation: a cross-taxa analysis of forest-dependent species. Biol Conserv 241:108368
R Development Core Team (2019) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Austria
Robinson WD, Brawn JD, Robinson SK (2000) Forest bird community structure in central Panama: influence of spatial scale and biogeography. Ecol Monog 70(2):209–235
Santos-Filho M, Peres CA, Da Silva DJ, Sanaiotti TM (2012) Habitat patch and matrix effects on small-mammal persistence in Amazonian forest fragments. Biod Conserv 21(4):1127–1147
Şekercioḡlu ÇH, Ehrlich PR, Daily GC, Aygen D, Goehring D, Sandí RF (2002) Disappearance of insectivorous birds from tropical forest fragments. Proc Natl Acad Sci 99(1):263–326
Şekercioḡlu ÇH (2006) Increasing awareness of avian ecological function. Trends Ecol Evol 21:464–471
Sick H (1984) Ornitologia brasileira. Editora Universidade de Brasília, Brasilia
Soares-Filho B, Rajão R, Macedo M, Carneiro A, Costa W, Coe M, Rodrigues H, Alencar A (2014) Cracking Brazil's forest code. Science 344(6182):363–364
SOS Mata Atlântica, INPE (2020) Atlas dos remanescentes florestais da Mata Atlântica: período 2018–2019. Relatório técnico, São Paulo
Thomas WW, De Carvalho AMV, Amorim AMA, Garrison J, Arbeláez AL (1998) Plant endemism in two forests in southern Bahia, Brazil. Biod Conserv 7(3):311–322
Thomas JP, Reid ML, Jung TS, Barclay RM (2019) Site occupancy of little brown bats (Myotis lucifugus) in response to salvage logging in the boreal forest. For Ecol Manag 451:117501
Uezu A, Metzger JP, Vielliard JM (2005) Effects of structural and functional connectivity and patch size on the abundance of seven Atlantic Forest bird species. Biol Conserv 123(4):507–519
Valente JJ, Betts MG (2019) Response to fragmentation by avian communities is mediated by species traits. Divers Distrib 25(1):48–60
Acknowledgements
We thank the reviewer and editor for all the suggestions and improvements on the manuscript. We thank several members of the Applied Ecology & Conservation lab for their help in the field work; local landowners for allowing us to work in their properties. We thank the financial support provided by Universidade Estadual de Santa Cruz (UESC/PROPP; 00220-1100.1039 and 00220-1100.1171), Fundação de Amparo à Pesquisa do Estado da Bahia (FAPESB, CNV0028/2011) and Conselho Nacional de Desenvolvimento Científico e Técnológico (CNPq; 69014416); the later also granted a fellowship to DF (CNPq: 307990/2018-4). This is the publication number 36 of the REDE SISBIOTA (CNPq 563216/2010-7).
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Morante-Filho, J.C., Benchimol, M. & Faria, D. Landscape composition is the strongest determinant of bird occupancy patterns in tropical forest patches. Landscape Ecol 36, 105–117 (2021). https://doi.org/10.1007/s10980-020-01121-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-020-01121-6