
Abstract
Context
Land use changes have modified the extent and structure of native vegetation, resulting in fragmentation of native species habitat. Connectivity is increasingly seen as a requirement for effective conservation in these landscapes, but the question remains: ‘connectivity for which species?’.
Objective
The aim of this study was to develop and then apply a rapid, expert-based, dispersal guild approach where species are grouped on similar fine-scale dispersal behaviour (such as between scattered trees) and habitat characteristics.
Methods
Dispersal guilds were identified using clustering techniques to compare dispersal and habitat parameters elicited from experts. We modelled least-cost paths and corridors between patches and individual movement probabilities within these corridors for each of the dispersal guilds using Circuitscape. We demonstrate our approach with a case study in the Tasmanian Northern Midlands, Australia.
Results
The dispersal guild approach grouped the 12 species into five dispersal guilds. The connectivity modelling of those five guilds found that broadly dispersing species in this landscape, such as medium-sized carnivorous mammals, were unaffected by fragmentation while from the perspective of the three dispersal guilds made up of smaller mammals, the landscape appeared highly fragmented.
Conclusions
Our approach yields biologically defensible outputs that are broadly applicable, particularly for conservation planning where data and resources are limited. It is a useful first step in multi-species conservation planning which aims to identify those species most in need of conservation efforts.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Human driven land use changes have modified the extent and structure of native vegetation, resulting in fragmentation of native species habitat. Species movement through landscapes declines as habitat fragmentation increases, with decreased connectivity resulting in reduced population viability and increased extinction risk beyond that caused by habitat loss alone (Caughley 1994; Fischer and Lindenmayer 2006; Brook et al. 2008). Land use between habitat patches can strongly influence connectivity. The most common conservation method to address the impacts of habitat fragmentation is to increase habitat area or increase connectivity between remnant habitat patches through wildlife corridors (Brost and Beier 2012).
A common approach to connectivity modelling is based on least-cost path analysis and graph theory (Urban and Keitt 2001; Adriaensen et al. 2003; Saura and Torné 2009; Foltête et al. 2012). Least-cost paths represent the least costly path across a cost surface, where the values assigned to cells in the cost surface represent the relative energetic costs, difficulty, or mortality risk of moving through that particular land cover type (Adriaensen et al. 2003; Sawyer et al. 2011). Least-cost analysis can also be used to identify least-cost corridors which represent the accumulated costs of movement between two patches (McRae and Kavanagh 2011). The significance of patches and links within a connectivity network can be quantified using the graph theoretic approach and calculation of network measures (Minor and Urban 2008; Rayfield et al. 2011). Another common approach that has been applied to connectivity modelling is based on circuit theory, whereby the landscape is conceptualized as a conductive surface within an electrical circuit, characterising resistance to movement for every raster grid cell, considering current flow as analogous to individual movement probabilities (McRae et al. 2008).
Connectivity modelling has been applied to landscape features or land-facets (Alagador et al. 2012; Brost and Beier 2012), single species (LaRue and Nielsen 2008; Tournant et al. 2013) and multiple single species modelled separately (Beier et al. 2009; Brás et al. 2013). In this paper we develop a modelling approach that is an intermediate between single species models and the landscape features or land-facets approach, characterising connectivity for groups of species based on shared dispersal and habitat characteristics. We extend the guild concept, defined as a “group of species whose members exploit similar resources in a similar manner” (Park and Allaby 2013), to include species with similar dispersal characteristics as well as those that exploit similar resources in a similar manner. Using the guild approach, information about single species can be meaningfully aggregated into common groups, providing greater generalisability of the results (Blaum et al. 2011). Conversely, adopting empirical, single species approaches to understand many species’ needs can be practically impossible to implement within time frames and costs reasonable for conservation planning (Noon et al. 2009; Blaum et al. 2011; Rudnick et al. 2012). Our research complements existing approaches to multi-species connectivity modelling for conservation planning which include assessing the overlap between connectivity outputs from multiple single species models (Beier et al. 2009), identifying links between habitats occupied by multiple species with distinct distributions and/or dispersal requirements (Brás et al. 2013) and identifying corridors for a range of dispersal abilities as opposed to specific species (Drielsma et al. 2007; Cushman and Landguth 2012; Synes et al. 2015). Additionally, our methods can potentially be used in experimental studies examining the ecology of connectivity for multiple species (e.g. Haddad et al. 2003; Frey-Ehrenbold et al. 2013).
The objective of this study was to develop the dispersal guild concept and then apply it to conservation planning using an expert-based parameterisation of connectivity modelling using a combination of least-cost path, circuit theory and graph theory methods. The approach can be used as a filtering process for multi-species conservation planning to identify those species in greatest need of conservation efforts. We demonstrate this approach using the Tasmanian Northern Midlands, Australia as a case study using a connectivity modelling framework which describes fine-scale dispersal patterns (Lechner et al. 2015b, c). Through a process of engaging experts in workshops we identified the species of critical concern and the habitat and dispersal characteristics of each. These individual target species were then grouped into dispersal guilds using cluster analysis. Connectivity was then modelled for each of these groups based on their shared dispersal and habitat characteristics. We used Graphab (Foltête et al. 2012) to calculate graph-metrics and least-cost paths between patches and Linkage Mapper (with Circuitscape) (McRae et al. 2008; McRae and Kavanagh 2011) to characterise least-cost corridors and individual movement probabilities for each of the dispersal guilds. By modelling dispersal guilds it was possible to capture a range of responses to fragmentation and thus identify those groups of species that may benefit from habitat restoration based on the spatial and functional requirements for increased connectivity. We conclude by discussing the application of this method to conservation planning.
Dispersal guilds to connectivity analysis methods
Case study
We use a case study in the Tasmanian Northern Midlands to demonstrate the application of the dispersal guild approach to conservation, as this landscape represents a typical connectivity planning problem example, where the focus of conservation is on connecting small fragmented, predominantly woodland remnant habitat. The Tasmanian Northern Midlands is one of fifteen designated Australian Biodiversity Hotspots containing numerous endemic plants and nationally and state listed threatened plants and animals (Department of Environment 2014). Widespread land clearing has left native vegetation highly fragmented, with mostly small and scattered remnants (Mooney et al. 2010). A planned expansion of irrigation schemes in the region will likely lead to agricultural intensification across the landscape (Gadsby et al. 2013), and further fragmentation of habitat in the region.
Expert elicitation
The selection of species, their parameterisation and validation of the model outputs were driven by an expert elicitation approach, over three (approximately 1 day) workshops and through engaging individual experts separately. The aim of each of the three workshops were as follows:
-
1.
To identify target species of conservation concern and available spatial and ecological data.
-
2.
To identify dispersal and habitat characteristics for the target species to be used as inputs into dispersal guild analysis and connectivity modelling.
-
3.
To present and refine draft modelling outputs.
The workshop attendees represented a core group of experts responsible for parameterising and selecting all the target species. This group was made up of 14 individuals over three workshops, with eight to eleven individuals attending per workshop (including the authors). This group included ecologists from academia (7), non-government conservation organisations (4), and government conservation agencies (3). The experts had a broad knowledge of the study area and the species, either through conducting research in the area or engaging in on ground conservation activities. These experts included individuals from three conservation organisations (the Tasmanian Land Conservancy, Greening Australia and Bush Heritage Australia) and the state government who have all been working in the study area for many decades. In addition to the workshop, six other experts were consulted separately to validate parameter estimates derived by the workshop attendees. These other experts had completed or were completing their PhD on specific target species, in many cases within the study area and had strong field experience which included observing and/or trapping the target species. Finally, two recognised field ecologists from academia with broad experience in the ecology of connectivity in Australia were also consulted.
Target species
Species considered to be of importance for conservation in the Tasmanian Northern Midlands were chosen at the first expert workshop. This workshop identified ground-based mammals as a key target group for conservation. Candidate species were reduced to those that depend only on native woodlands, the original vegetation cover that was fragmented by clearing as distinct from native grasslands, non-native vegetation or pasture. For example, the Eastern Quoll (Dasyurus viverrinus) was excluded because it readily utilises cleared areas such as non-native pastures and improving landscape connectivity is unlikely to be as important for the conservation of this species as other factors such as managing introduced predators. Twelve ground based mammals were identified and then grouped into dispersal guilds using a cluster analysis in subsequent workshops.
Approach to characterising connectivity
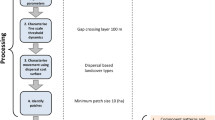
Our approach to characterising connectivity used in this paper is based on the GAP CLoSR framework for modelling fine-scale dispersal behaviour described in Lechner and Lefroy (2014) and Lechner et al. (2015a, b, c). The framework consists of three key parameters (Fig. 1):

Conceptual model of fine-scale connectivity behaviour of mammal fauna, where the likelihood of individuals moving between two patches is a function of two thresholds, the interpatch-crossing distance and gap-crossing distance, and the dispersal cost of landcover features (such as roads)
-
1.
Minimum patch size The minimum area of habitat that can support a population. Below this minimum patch size, vegetation no longer contributes to habitat but may aid in dispersal such as in the case of a wildlife corridor.
-
2.
Gap-crossing distance threshold The maximum distance a species will cross between connectivity elements (such as scattered trees), which limits the distances of open ground (gaps) individuals will move across.
-
3.
Interpatch-crossing distance The maximum total distance that a species can move between habitat patches using connectivity elements that are within the gap-crossing distance.
We also included additional parameters characterising land cover for habitat and connectivity elements. Habitat and connectivity elements were classified as either above 1 m (e.g. trees) or below 1 m (e.g. undergrowth) from ground level (Table 1).
The parameters (Table 1) for each of the 12 species were the result of a literature review and discussions with individual experts at the second workshop. These parameters were refined in the final workshop when the outputs from the model were presented to the expert group.
A critical component of the framework is the inclusion of fine-scale dispersal behaviour which is often absent from many common connectivity modelling approaches. In order for species to move long distances between patches there is a need for connectivity elements such as corridors, or stepping stones to facilitate movement (Fischer and Lindenmayer 2002; Van Der Ree et al. 2004).
Dispersal guilds and cluster analysis
Common dispersal and habitat characteristics of the target species were identified using cluster analysis to define the dispersal guilds. Two cluster analysis methods were used; a hierarchical clustering analysis in R using the pvclust package version 1.2-2 (Suzuki and Shimodaira 2006) and the Partitioning Around Medoids (PAM) algorithm (Kaufman and Rousseeuw 1990) in the Cluster package for R 2.15.2. A hierarchical cluster analysis is based on an agglomerative algorithm and produces a dendrogram or tree diagram used to illustrate the arrangement of the clusters. The PAM algorithm groups objects into clusters based on the number of clusters specified beforehand. In our study a range of cluster numbers were tested. To determine the stability of the clustering, which reflects the validity of a cluster solution (i.e. number and grouping), we calculated the average silhouette width, which provides a standard measure of cluster isolation (Kaufman and Rousseeuw 1990). High values of average silhouette width is one way to identify the appropriate number of clusters within which the data naturally falls. PAM uses a medoid as its measure of cluster centre as opposed to a centroid based on mean values. This method is more robust in the presence of noise and outliers than mean-based approaches (e.g. k-mean). The two cluster analysis methods have very different approaches and different graphical outputs, providing a method of testing whether clustering outputs are independent of the assumptions in the clustering methods.
To complete the cluster analysis, categorical variables including habitat above and below 1 m were converted to a matrix of zeros and ones as both cluster analysis methods require all variables to be continuous. The converted categorical data were then combined with the continuous data describing dispersal distances, and singular value decomposition was applied to create new continuous variables, each of which represents a composite of the original datasets.
The outputs of both cluster analysis methods were presented at the third expert workshop for validation. The cluster analysis provided an accessible graphical way of summarising the complex similarity and dissimilarity of ecological parameters between species that may have been difficult for an expert panel to identify from the raw data alone.
A single value for each of the habitat characteristics and dispersal thresholds is required to represent each dispersal guild for connectivity modelling. In this study we used average values for the continuous variables (e.g. interpatch-dispersal distance) and the most common value for the categorical variables (e.g. habitat).
Characterising habitat and dispersal-cost surfaces using spatial data
Habitat and dispersal-cost surfaces were produced for connectivity modelling to spatially represent the attributes of each dispersal guild. All spatial layers, regardless of their original spatial resolution, were aggregated to 50 m, the smallest pixel size at which the connectivity model would run for our study area.
Structural vegetation mapping from Tasmanian Land Conservancy vegetation data (5 m pixel size) was used to spatially represent habitat and connectivity elements (Sprod 2013). The spatial representation of the connectivity elements was used to create the dispersal cost-surface layer. Spatial data were based on TASVEG 2.0 mapping (DPIPWE 2013) and refined through interpretation of aerial imagery. Mapped polygons were refined and then attributed according to estimated density of vegetation structures above and below 1 m height (Table 1).
The above- and below 1 m vegetation classes were converted into two binary layers to spatially represent all combinations of habitat and connectivity described in Table 1. Habitat and connectivity were characterised as: (i) vegetation > 1 m; (ii) vegetation < 1 m; (iii) vegetation > 1 m AND vegetation < 1 m; and (iv) vegetation over 1 m OR vegetation under 1 m (i.e. either i, or ii) (Table 1; Fig. A.1). An AND function represents cases where vegetation above and below 1 m need to be present, while the OR function represents cases where any form of vegetation is present.
The dispersal-cost surface characterises how land cover type between habitat patches reduces or prevents movement. It is a result of combining the gap-crossing layer and the land cover resistance surface.
The gap-crossing distance threshold layer was simulated by creating the gap-crossing layer (see Lechner et al. (2015a, b, c) for more details). The gap-crossing layer identified distances between connectivity elements and patches beyond the movement threshold that act as barriers to dispersal. The input vegetation layers were buffered by half of the gap-crossing distance threshold (Fig. A.2). Where connectivity elements or patches fall within the gap-crossing distance threshold, buffers touch or overlap indicating connectivity between patches. Areas mapped outside the buffer area describe areas in which dispersal cannot take place (i.e. barriers).
Resistance to dispersal between patches is characterised by increased movement costs associated with land cover which is assumed to be linear. For example, if land cover with high dispersal resistance doubled the movement cost, the interpatch-crossing distance threshold would be reduced from 1.1 km to 550 m. In the study area we used generic land cover classes with dispersal costs assigned to each pixel based on expert opinion for small and medium mammals (Table A.1). We used the Land Use Tasmania spatial dataset (Brown 2011) for the spatial characterisation of resistance (Fig. A.3 caption for more details).
In the final step, the dispersal-cost surface was created by combining the binary gap-crossing layer with the resistance surface based on land cover. The dispersal-cost values are a function of: (a) pixel size (e.g. if the pixel size is 50 m and there is no resistance the cost should be 50 m); (b) land cover resistance (200 % resistance means a pixel size with of 50 m will have a value of 100 m); and (c) the presence of connectivity elements below the gap-crossing distance identified from the gap-crossing layer.
A summary of the processing rule set:
-
Connectivity elements take precedence over all other land cover classes, because dispersal cannot occur in the absence of structural connectivity;
-
The dispersal cost for each pixel was calculated as an average of all land covers.
The result was a resistance layer that recognises threshold dynamics by ensuring there was no dispersal where gaps are too large between connectivity elements, but still models cumulative costs where dispersal was considered possible but may be impeded by land use.
Connectivity modelling approach
Connectivity was modelled using Graphab (Foltête et al. 2012) a graph-network connectivity modelling software that uses least-cost paths and Linkage Mapper (McRae and Kavanagh 2011) for identifying least-cost corridors. The least-cost analysis was based on cumulative cost in relation to land cover resistance (Minor and Urban 2007; Dale and Fortin 2010; Etherington and Holland 2013).
Using the graph theoretic approach and the Graphab software we characterised patch isolation by identifying groups of patches that are linked to each other but isolated from other groups of patches. These groups of interlinked patches are known as components. The patterns in the size and shape of the components can be used to characterize fragmentation and isolation, and to locate barriers to connectivity. We also calculated a number of landscape-scale graph-metrics describing component characteristics: mean size of components (km2), size of the largest component (km2), number of components and the integral index of connectivity (IIC) dispersal metric (Minor and Urban 2008; Rayfield et al. 2011). The IIC measures the probability that two points randomly placed in a landscape are in habitat areas that can be reached (Pascual-Hortal and Saura 2006; Saura and Pascual-Hortal 2007). High values indicate more connected landscapes.
We then combined the least-cost corridors, representing areas with cost-weighted distances below the interpatch dispersal distance threshold with Circuitscape’s Pinch Point Mapper in the Linkage Mapper tool (see McRae and Kavanagh 2011). The Pinch Point Mapper characterises wildlife corridor pinch points based on random walk patterns between patches identified using circuit theory (McRae et al. 2008). Pinch-points (or choke-points) are areas where animal movement is funnelled within corridors and represent areas where linkages are most vulnerable to being severed (McRae et al. 2008). Using this combination of methods means we can characterise connectivity based on the two dispersal thresholds (interpatch and gap crossing) while characterising dispersal behaviour more realistically based on a random walk. This is not possible when modelling with Circuitscape in isolation as it does not allow for maximum dispersal distances.
Sensitivity analysis
The connectivity outputs can be assessed for sensitivity to the input data and parameterisation. A comparison of the modelled connectivity with and without dispersal costs from the gap-crossing layer and land use allows assessment of the importance of interpatch-crossing distance thresholds for landscape-scale connectivity.
The results can be interpreted in two ways:
-
1.
Does uncertainty in the mapping of fine-scale vegetation and land use effect connectivity outputs?
-
2.
Do corridors, vegetation and connectivity elements (e.g. scattered trees) make a key contribution to connectivity?
Further sensitivity analysis that characterises the number of components versus interpatch-crossing distance for all combinations of habitat input layers identifies the critical scales that need to be considered.
Tasmanian Northern Midlands case study
Expert elicitation of dispersal and habitat values for each species
A literature review revealed very few empirical studies that could be used to confidently characterise dispersal distances for our species. As a result, the values for the dispersal and habitat parameters for each species were derived through the consensus of an expert panel in a workshop environment (Table A.2). For some species experts outside of the workshop group were suggested by the panel and contacted or literature was identified. Values derived from outside experts and literature were reviewed by the panel. The experts agreed that the values represent only approximations, so the relative differences between species are likely to be more accurate than the absolute values. The experts also agreed that due to the lack of knowledge of our target species a general sensitivity analysis was useful for assessing the uncertainty on the elicited estimations.
Identify dispersal guilds and connectivity model input parameters
The dendrogram produced by the hierarchical cluster analysis qualitatively identified six groups made up of one to four species (Fig. 2). The output from the PAM clustering method set at six clusters found exactly the same membership as the hierarchical cluster analysis. The average silhouette widths (a method for assessing the strength of clustering) for six clusters calculated with the PAM clustering method was 0.591, which according to Kaufman and Rousseeuw (1990) indicates “reasonable structure has been found”. Average silhouette widths below 0.5 indicate weak structure that could be artificial or no structure at all. With five to seven clusters, all had greater than 0.5 average silhouette width (Fig. A.4b), so assigning between five and seven dispersal guilds was supported by this statistical analysis.

Hierarchical cluster analysis where height describes the similarity between individual species. Labelled dispersal guilds identified post hoc through discussion with experts. Each colour represents a dispersal guild. Note that the Ring-tailed Possum was included in the Arboreal dispersal guild based on expert opinion even though it is found in a separate cluster
The results from the cluster analysis were then presented to the expert group in workshop 3. The expert group agreed with the clustering except for the Ring-tailed Possum (Pseudocheirus peregrinus). They recommended that the Ring-tailed Possum should be grouped with the other arboreal species. It was grouped in a class of its own due to the fact that it is the only species which used connectivity elements over 1 m and elements under 1 m (Table A.2). The expert panel also agreed with the identification of the Tasmanian Devil (Sarcophilus harrisii) and Spotted-tailed Quoll (Dasyurus maculatus) as separate single-species dispersal guilds as their characteristics were unique.
Five dispersal guilds from the original six cluster groupings were agreed to by the expert panel. The expert panel labelled the guilds according to the ecological characteristics that united group members (Fig. 2; Table 2). The cluster analysis identified three guilds made up of smaller mammals with shorter interpatch-crossing and gap-crossing dispersal distances and smaller habitat requirements. Two separate single-species dispersal guilds were made up of medium carnivores with long, but distinct dispersal distances.
For each dispersal guild, a single value for each of the habitat characteristics and dispersal thresholds is required for connectivity modelling. For the two medium carnivore guilds their original values were used, while for the three other dispersal guilds the average values were used. In cases with categorical variables the combination of vegetation above- and/or below 1 m that was found in the majority of cases was used (Table 2).
Dispersal guild connectivity outputs
The connectivity analysis of the five dispersal guilds (Figs. 3, 4; Figs. A.5–A.9) showed large differences in the effects of fragmentation. For the two medium carnivores the landscape essentially appears connected. For the Tasmanian Devil, all patches were connected directly or indirectly to other patches (Fig. 3) and all but 4 patches were connected for the Spotted-tailed Quoll (Fig. A.6). Additionally, the least-cost corridor analysis showed that the medium-sized carnivore guilds with long interpatch and gap crossing dispersal distances utilises more of the matrix than the guilds with shorter dispersal distances (Figs. 3, 4).

Connectivity analysis for the “Medium Carnivore–Tasmanian Devil” dispersal guild with an interpatch-crossing distance threshold of 10,000 m. Black lines indicate least cost paths (LC) between connected patches. *Least-cost corridors width calculated with 10,000 m cost-weighted distance threshold. Pinch-points identified with Circuitscape within corridors

Connectivity analysis for the ‘Dense ground cover dependent’ dispersal guild with an interpatch-crossing distance threshold of 900 m. Component boundaries in blue are located at the midpoint between patches identifying groups of patches that are linked, red lines indicate least cost paths between connected patches. *Least-cost corridors width calculated with 900 m cost-weighted distance threshold. Pinch-points identified with Circuitscape within corridors. Inset illustrates that connectivity for this guild exists only at fine scales
From the perspective of the three dispersal guilds of smaller mammals, the landscape appears highly fragmented with the majority of the connected areas in the east and west of the study area and most patches in the central region isolated or connected to relatively few patches (Figs. A.5–A.9). The average component size was between 20 and 28 km2 (Table 3). The graph metric value reflects the visual patterns with the IIC having higher values for the medium carnivores versus the small mammals indicating greater connectivity at the landscape scale (Table 3).
Sensitivity analysis
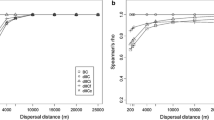
The sensitivity analysis found that all but one of the dispersal guilds,’Arboreal’, showed very similar connectivity outputs in terms of the component patterns regardless of the dispersal costs. In the case of medium mammals the landscape is always connected, but the availability of pathways vary and in the case of the two guilds of small mammals, ‘Not Woodland dependent and/or riparian’ and ‘Dense ground cover dependent’, most patches and the majority of the large components are isolated due to their relatively short interpatch-crossing distances. However, the ‘Arboreal’ dispersal guild showed large differences with or without the inclusion of fine-scale connectivity elements and land use. An assessment of the number of components versus interpatch-crossing distance for all combinations of habitat input layers found that at interpatch-crossing distances of approximately 1000 m there is a rapid drop off in connectivity within the landscape (Fig. 5). This analysis, however, did not include the impact of the gap-crossing threshold.

Interpatch-dispersal distance sensitivity analysis for vegetation a over 1 m, b over 1 m and under 1 m, and c under 1 m characterised by the number of components versus the interpatch-crossing distance. For this analysis the gap-crossing distance was excluded
Application of the results
For our specific case study we found that broadly dispersing medium-sized carnivores, are unlikely to have their landscape-scale connectivity increased with revegetation efforts in the Tasmanian Midlands because no or few habitat patches are considered isolated from their perspective. In such instances benefits may result from increasing the condition of habitat through restoration and critical components of wildlife corridors (i.e. pinch-points) or from addressing other threats such as devil facial tumour disease (Hawkins et al. 2006) or the effects of competition from introduced predators such as cats (Hollings et al. 2013). Where habitat patches for species are separated by more than the interpatch-crossing distance, revegetation may be effective, and our results provide a guide as to suitable areas for such activity. This might suit the dispersal guilds ‘Not Woodland dependent or riparian’ and ‘Dense ground cover dependent’ with average interpatch-crossing distances of 1000 and 900 m respectively. The corridor analysis (Figs. 3, 4) showed particularly striking differences in the use of the matrix between the medium carnivores and the small mammal dispersal guilds.
Connectivity was also examined for all species by plotting the number of components against the interpatch-crossing distance. This shows that the landscape becomes very fragmented for species with interpatch-crossing distances less than 1000 m. Four of the species tested (Long-tailed Mouse (Pseudomys higginsi), Swamp Rat (Rattus lutreolus), Eastern Pygmy Possum (Cercartetus nanus) and Little Pygmy Possum (Cercartetus lepidus)) have interpatch-crossing distances less than or equal to 1000 m. For those four species, the results show that the landscape is highly fragmented from their perspective. Reconnecting the landscape may have positive impacts on landscape movement of these species. A similar pattern can be seen when plotting the IIC versus the interpatch-crossing distance (Fig. A.10).
The outputs of our study could be refined with local knowledge of species distribution. In their current form, the outputs can provide general guidance as to where connectivity may be limiting (or not-limiting) and for which species connectivity may be limited and therefore aid in identifying where future conservation challenges may lie. The dispersal guild method can be expanded to assess future scenarios, barriers, and identify locations for revegetation (e.g. McRae et al. 2012; Foltête et al. 2014; Lechner et al. 2015a).
Application of dispersal guilds to conservation planning
The methods outlined in this study show great potential in conservation planning for targets that are intermediate between single species and general landscape features. The dispersal guild approach provides a way of using a relatively rapid assessment based on expert opinion to prioritise conservation effort for groups of species that view the landscape differently. Our approach to grouping species is supported by a recent empirical study by Brodie et al. (2014) which assessed single-species and multiple species habitat corridors in the tropical forests of Borneo and suggested that multispecies habitat connectivity plans should be tailored to groups of ecologically similar species to maximize effectiveness. Our approach presents a form of filtering in multispecies conservation planning by identifying those species that are likely to experience greatest benefit form conservation efforts (e.g. Noon et al., 2009). Such coarse filter approaches have been used previously for characterising connectivity for environmentally-similar habitats (Alagador et al. 2012) or land-facets (Beier and Brost 2010; Brost and Beier 2012), whereas our approach explicitly identifies which groups of species are affected by different aspects of fragmentation such as lack of connectivity elements, potential pinch-points and distances between patches. The method described in this paper also differs from existing, commonly applied single species approaches or other approaches aimed at modelling connectivity for multiple species (e.g. Beier and Brost 2010; Brás et al. 2013; Brodie et al. 2014; Koen et al. 2014).
Dispersal guilds could be used as part of a conservation action planning process to assess the viability of focal conservation targets (Poiani et al. 2000; The Nature Conservancy (2007) “Step 3: Assess Viability of Focal Conservation Targets” and CMP (2013) “1D. Analyze the Conservation Situation”) and identify the relative benefit different guilds would be likely to derive from alternative interventions (wildlife corridors, improved habitat quality, increased habitat patch size). For example, the case study graphically highlighted that investment in improved connectivity would be less effective than improving habitat area or quality for those species where the dispersal distance and gap-crossing threshold exceed the general level of fragmentation. The thresholds of connectivity and patterns of connectivity associated with each guild could be used in a triage approach, where conservation efforts are best directed at areas where investment is likely to produce the greatest benefit (Bottrill et al. 2008).
Our method can be applied rapidly, within approximately three months, providing experts with sufficient knowledge are available. The expert-based approach is particularly suited to connectivity modelling that uses simple thresholds such as the inter-patch and gap-crossing dispersal distance where species exhibit threshold dynamics such as a foray search strategy (Doerr et al. 2011; Smith et al. 2013) that can be modelled using cumulative cost approaches. It is important though to ensure that: (a) experts have sufficient knowledge for parameterising the model, (b) the required parameters are explained clearly to the experts and (c) it is understood that approximations are suitable for this process. While expert opinion is considered to be suboptimal for parameterising connectivity models (Clevenger et al. 2002; Zeller et al. 2012; Cushman et al. 2013) the alternative of empirical field based research would be a much greater undertaking for multiple species (Compton et al. 2007; Rudnick et al. 2012). Furthermore, the literature review, which was supported by data from a 2014 meta-analysis of all empirical studies of connectivity in Australia (Doerr et al. 2014), found very few empirical studies with the necessary parameters for our model or parameters that could be applied outside of the original study area due to locally unique characteristics. Thus in many cases an expert approach would be the only practical option.
Conclusions
Decision makers need tools that are sufficiently flexible and dynamic to assess connectivity without being too complex, difficult to use or time-consuming. The approach described here of characterising dispersal guilds provides greater generalizability than single species connectivity modelling, and is therefore likely to be better suited to a whole of landscape, multi-species approach to conservation planning. It provides a biologically defensible first pass connectivity assessment using available expert knowledge when the time or resources are not available to collect quantitative dispersal and distribution data for numerous species. The outputs provide a useful basis from which to prioritise conservation investment for connectivity and further research.
References
Adriaensen F, Chardon JP, De Blust G, Swinnen E, Villalba S, Gulinck H, Matthysen E (2003) The application of “least-cost” modelling as a functional landscape model. Landsc Urban Plan 64:233–247
Alagador D, Triviño M, Cerdeira JO, Brás R, Cabeza M, Araújo MB (2012) Linking like with like: optimising connectivity between environmentally-similar habitats. Landsc Ecol 27:291–301
Beier P, Brost B (2010) Use of land facets to plan for climate change: conserving the arenas, not the actors. Conserv Biol 24:701–710
Beier P, Majka DR, Newell SL (2009) Uncertainty analysis of least-wildlife for designing linkages. Ecol Appl 19:2067–2077
Blaum N, Mosner E, Schwager M, Jeltsch F (2011) How functional is functional? Ecological groupings in terrestrial animal ecology: towards an animal functional type approach. Biodivers Conserv 20:2333–2345
Bottrill MC, Joseph LN, Carwardine J, Bode M, Cook C, Game ET, Grantham H, Kark S, Linke S, McDonald-Madden E, Pressey RL, Walker S, Wilson KA, Possingham HP (2008) Is conservation triage just smart decision making? Trends Ecol Evol 23:649–654
Brás R, Cerdeira JO, Alagador D, Araújo MB (2013) Linking habitats for multiple species. Environ Model Softw 40:336–339
Brodie JF, Giordano AJ, Dickson B, Hebblewhite M, Bernard H, Mohd-Azlan J, Anderson J, Ambu L (2014) Evaluating multispecies landscape connectivity in a threatened tropical mammal community. Conserv Biol 29:122–132
Brook BW, Sodhi NS, Bradshaw CJA (2008) Synergies among extinction drivers under global change. Trends Ecol Evol 23:453–460
Brost BM, Beier P (2012) Comparing linkage designs based on land facets to linkage designs based on focal species. PLoS ONE 7:e48965
Brown M (2011) Land use Tasmania: a technical report outlining the creation of the Draft Tasmanian summer 2009/2010. Unpublished report DPIPWE, Tasmania
Caughley G (1994) Directions in conservation biology. J Anim Ecol 63:215–244
Clevenger AP, Wierzchowski J, Chruszcz B, Gunson K (2002) GIS-generated, expert-based models for identifying wildlife habitat linkages and planning mitigation passages. Conserv Biol 16:503–514
CMP (2013) Open standards for the practice of conservation, version 3. Conservation Measures Partnership
Compton BW, McGarigal K, Cushman SA, Gamble LR (2007) A resistant-kernel model of connectivity for amphibians that breed in vernal pools. Conserv Biol 21:788–799
Cushman SA, Landguth EL (2012) Multi-taxa population connectivity in the Northern Rocky Mountains. Ecol Model 231:101–112
Cushman SA, Mcrae B, Adriaensen F, Beier P, Shirley M, Zeller K (2013) Biological corridors and connectivity. Key Top Conserv Biol 2:384–404
Dale MRT, Fortin M-J (2010) From graphs to spatial graphs. Annu Rev Ecol Evol Syst 41:21–38
Department of Environment (2014) Australia’s 15 national biodiversity hotspots. Commonw Gov. http://www.environment.gov.au/biodiversity/conservation/hotspots/national-biodiversity-hotspots#hotspot4
Doerr VAJ, Doerr ED, Davies MJ (2011) Dispersal behaviour of Brown Treecreepers predicts functional connectivity for several other woodland birds. Emu 111:71–83
Doerr ED, Doerr VAJ, Davies MJ, Mcginness HM (2014) Does structural connectivity facilitate movement of native species in Australia’s fragmented landscapes?: a systematic review protocol. Environ Evid 3:1–8
DPIPWE (2013) TASVEG 3.0. Tasmanian vegetation monitoring and mapping program. Resource Management and Conservation Division, Hobart, Tasmania
Drielsma M, Manion G, Ferrier S (2007) The spatial links tool: automated mapping of habitat linkages in variegated landscapes. Ecol Model 200:403–411
Etherington TR, Holland PE (2013) Least-cost path length versus accumulated-cost as connectivity measures. Landscape Ecol 28:1223–1229
Fischer J, Lindenmayer DB (2002) The conservation value of paddock trees for birds in a variegated landscape in southern New South Wales. 1. Species composition and site occupancy patterns. Biodivers Conserv 11:807–832
Fischer J, Lindenmayer DB (2006) Habitat fragmentation and landscape change: an ecological and conservation synthesis. Island Press, Washington
Foltête JC, Clauzel C, Vuidel G (2012) A software tool dedicated to the modelling of landscape networks. Environ Model Softw 38:316–327
Foltête J-C, Girardet X, Clauzel C (2014) A methodological framework for the use of landscape graphs in land-use planning. Landsc Urban Plan 124:140–150. doi:10.1016/j.landurbplan.2013.12.012
Frey-Ehrenbold A, Bontadina F, Arlettaz R, Obrist MK (2013) Landscape connectivity, habitat structure and activity of bat guilds in farmland-dominated matrices. J Appl Ecol 50:252–261
Gadsby S, Lockwood M, Moore S, Curtis A (2013) Tasmanian midlands socio-economic profile. Hobart, Tasmania, Landscape and Policy Hub, University of Tasmania
Haddad NM, Bowne DR, Cunningham A, Danielson BJ, Levey DJ, Sargent S, Spira T (2003) Corridor use by diverse Taxa. Ecology 84:609–615
Hawkins CE, Baars C, Hesterman H, Hocking GJ, Jones ME, Lazenby B, Mann D, Mooney N, Pemberton D, Pyecroft S, Restani M, Wiersma J (2006) Emerging disease and population decline of an island endemic, the Tasmanian devil Sarcophilus harrisii. Biol Conserv 131:307–324
Hollings T, Jones M, Mooney N, McCallum H (2013) Wildlife disease ecology in changing landscapes: mesopredator release and toxoplasmosis. Int J Parasitol Parasites Wildl 2:110–118
Kaufman L, Rousseeuw PJ (1990) Finding groups in data: an introduction to cluster analysis. Wiley, New York
Koen EL, Bowman J, Sadowski C, Walpole AA (2014) Landscape connectivity for wildlife: development and validation of multispecies linkage maps. Methods Ecol Evol 5:626–633
LaRue MA, Nielsen CK (2008) Modelling potential dispersal corridors for cougars in midwestern North America using least-cost path methods. Ecol Model 212:372–381
Lechner AM, Brown G, Raymond CM (2015a) Modeling the impact of future development and public conservation orientation on landscape connectivity for conservation planning. Landscape Ecol 30:699–713
Lechner AM, Doerr V, Harris RMB, Doerr E, Lefroy EC (2015b) A framework for incorporating fine-scale dispersal behaviour into biodiversity conservation planning. Landsc Urban Plan 141:11–23
Lechner AM, Harris RMB, Doerr V, Doerr E, Drielsma M, Lefroy EC (2015c) From static connectivity modelling to scenario-based planning at local and regional scales. J Nat Conserv 28:78–88
Lechner AM, Lefroy EC (2014) General approach to planning connectivity from LOcal Scales to Regional (GAP CLoSR): combining multi-criteria analysis and connectivity science to enhance conservation outcomes at regional scale. Centre for Environment, University of Tasmania. www.nerplandscapes.edu.au/publication/GAP_CLoSR
McRae BH, Dickson BG, Keitt TH, Shah VB (2008) Using circuit theory to model connectivity in ecology, evolution, and conservation. Ecology 89:2712–2724
McRae BH, Hall SA, Beier P, Theobald DM (2012) Where to restore ecological connectivity? Detecting barriers and quantifying restoration benefits. PLoS ONE 7:e52604
McRae BH, Kavanagh DM (2011) Linkage mapper connectivity analysis software. The Nature Conservancy, Seattle
Minor ES, Urban DL (2007) Graph theory as a proxy for spatially explicit population models in conservation planning. Ecol Appl 17:1771–1782
Minor ES, Urban DL (2008) A graph-theory framework for evaluating landscape connectivity and conservation planning. Conserv Biol 22:297–307
Mooney C, Defenderfer D, Anderson M (2010) Reasons why farmers diversify northern midlands, Tasmania. Rural Industries Research and Development Corporation, Canberra
Noon BR, McKelvey KS, Dickson BG (2009) Multispecies conservation planning on U.S. Federal Lands. Models for Planning Wildlife Conservation in Large Landscapes. doi:10.1016/B978-0-12-373631-4.00003-4
Park C, Allaby M (2013) A Dictionary of Environment and Conservation, 2nd edn. Oxford University Press, Oxford
Pascual-Hortal L, Saura S (2006) Comparison and development of new graph-based landscape connectivity indices: towards the priorization of habitat patches and corridors for conservation. Landscape Ecol 21:959–967
Poiani KA, Richter BD, Anderson MG, Richter HE (2000) Biodiversity conservation at multiple scales: functional sites, landscapes, and networks. BioScience 50:133
Rayfield B, Fortin MJ, Fall A (2011) Connectivity for conservation: a framework to classify network measures. Ecology 92:847–858
Rudnick DA, Ryan SJ, Beier P, Cushman SA, Dieffenbach F, Epps C, Gerber LR, Hartter J, Jenness JS, Kintsch J, Merenlender AM (2012) The role of landscape connectivity in planning and implementing conservation and restoration priorities. Issue Ecol 16:1–23
Saura S, Pascual-Hortal L (2007) A new habitat availability index to integrate connectivity in landscape conservation planning: comparison with existing indices and application to a case study. Landsc Urban Plan 83:91–103
Saura S, Torné J (2009) Conefor Sensinode 2.2: a software package for quantifying the importance of habitat patches for landscape connectivity. Environ Model Softw 24:135–139
Sawyer SC, Epps CW, Brashares JS (2011) Placing linkages among fragmented habitats: do least-cost models reflect how animals use landscapes? J Appl Ecol 48:668–678
Smith MJ, Forbes GJ, Betts MG (2013) Landscape configuration influences gap-crossing decisions of northern flying squirrel (Glaucomys sabrinus). Biol Conserv 168:176–183
Sprod D (2013) A structural vegetation cover dataset for the Midlands of Tasmania. Unpublished report, Tasmanian Land Conservancy, Hobart
Suzuki R, Shimodaira H (2006) Pvclust: an R package for assessing the uncertainty in hierarchical clustering. Bioinformatics 22:1540–1542
Synes NW, Watts K, Palmer SCF, Bocedi G, Bartoń KA, Osborne PE, Travis JMJ (2015) A multi-species modelling approach to examine the impact of alternative climate change adaptation strategies on range shifting ability in a fragmented landscape. Ecol Inform 30:222–229
The Nature Conservancy (2007) Conservation action planning: developing strategies taking action, and measuring success at any scale. Overview of Basic Practices Version, February 2007
Tournant P, Afonso E, Roué S, Giraudoux P, Foltête JC (2013) Evaluating the effect of habitat connectivity on the distribution of lesser horseshoe bat maternity roosts using landscape graphs. Biol Conserv 164:39–49
Urban D, Keitt T (2001) Landscape connectivity: a graph-theoretic perspective. Ecology 82:1205–1218
Van Der Ree R, Bennett AF, Gilmore DC (2004) Gap-crossing by gliding marsupials: thresholds for use of isolated woodland patches in an agricultural landscape. Biol Conserv 115:241–249
Zeller KA, McGarigal K, Whiteley AR (2012) Estimating landscape resistance to movement: a review. Landscape Ecol 27:777–797
Acknowledgments
This project was funded by the Australian Government Sustainable Regional Development Program in conjunction with the National Environmental Research Program’s Landscapes and Policy research Hub. We would like to thank the following people who contributed to this research Amy Koch, Bronwyn Fancourt, Chris Johnson, Christine Fury, Erik Doerr, Felicity Faulkner, Gareth Davies, Kirsty Dixon, Kirstin Proft, Louise Gilfedder, Mat Appleby, Menna Jones, Neil Davidson, Nick Fitzgerald, Rebecca Harris, Sarah Maclagan, Shannon Troy and Veronica Doerr. We would also like to thank the two peer reviewers (Robby Marrotte and an anonymous reviewer) for their insightful and constructive comments which greatly improved the clarity of the paper.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Lechner, A.M., Sprod, D., Carter, O. et al. Characterising landscape connectivity for conservation planning using a dispersal guild approach. Landscape Ecol 32, 99–113 (2017). https://doi.org/10.1007/s10980-016-0431-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-016-0431-5