
Abstract
Context
Land use changes and intensification have been amongst the major causes of the on-going biodiversity decline in Europe. A better understanding and description of how different levels of land use intensity affect biodiversity can support the planning and evaluation of policy measures.
Objectives
Our study investigates how land use-related landscape characteristics affect bird diversity, considering different spatial scales and species groups with characteristic habitat use.
Methods
We used breeding bird census data from 2693 observation points along 206 transects and applied a random effects hurdle model to describe the influence of the landscape characteristics altitude, forest proportion, patch density, land cover diversity, and land use intensity on avian species richness.
Results
Land use intensity and related landscape characteristics formed an important explanatory variable for bird richness. Increasing land use intensity was accompanied by a decrease in bird species richness. While forest bird richness decreased with a decreasing amount of forest cover, farmland species richness increased. This led to a bird diversity peak in extensively used semi-open landscapes. The influence of land cover diversity on species richness was small. Increasing patch density had positive effects on forest birds, but affected farm birds negatively. The strongest correlation between land use-based indicators and bird diversity was determined using spatial indicators at a close range around observation points (100–500 m radius).
Conclusions
Our results assist interpretation of the Pan-European Common Bird Indices and emphasize the importance of using multifaceted and thoroughly selected indicators in the context of biodiversity monitoring and decision-making support.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
In the past millennia farming and settlement activities transformed the European landscape from a mostly woodland-dominated landscape to a mosaic of meadows, pastures, arable fields, hedges, woods, and settlements. While most natural habitats were cleared away, secondary open and semi-open habitats were created and populated by plants and animals adapted to them (Tscharntke et al. 2012). Those habitats and species contribute greatly to the current European biodiversity (Pimentel et al. 1992; Tscharntke et al. 2005). During the last few decades land use change in general (Sala et al. 2000; Tasser et al. 2008; Zimmermann et al. 2010) and the ongoing agricultural intensification in particular (Benton et al. 2003; Culman et al. 2010) have prompted a widespread decline in biodiversity. Furthermore, Dullinger et al. (2013) demonstrated that effects of land use change on biodiversity might currently be seriously underestimated as the reaction of many plant and animal populations lags behind contemporary environmental degradation. For birds—one of the best observed and well known taxa—population decline in cultivated landscapes has been very well documented and mainly associated with agricultural intensification (Krebs et al. 1999; Donald et al. 2001; Wretenberg et al. 2010).
While intensified land use is doubtless one of the main causes of biodiversity loss, low-intensity land-use systems may be an essential element in large-scale biodiversity protection (Tscharntke et al. 2005; Wright et al. 2012; Baudron and Giller 2014; Wehrden et al. 2014). International policies have adopted various measures to counteract the developments that led to a depletion of biodiversity and natural resources. The highly financed European agri-environment schemes (AES) are an example of steps aimed to reduce the negative influence of agriculture on biodiversity by paying farmers to apply environmentally friendly management practices. However, evaluation of AES has shown mixed results, and it is still largely unclear whether and how AES contribute to policy objectives aimed at halting the depletion of biodiversity (Butler et al. 2009; Wilson et al. 2010; Kleijn et al. 2011). The effects of AES differ between taxa and seem to be influenced by the applied scale (Gonthier et al. 2014).
Biodiversity is not only affected by local habitat availability and quality, but also by the surrounding landscape features (Concepción et al. 2008; Concepción et al. 2012). For the development of efficient and wise biodiversity policies it is essential to understand how land use-related landscape characteristics affect biodiversity patterns on the local and the landscape scale (Concepción et al. 2008; Tscharntke et al. 2012; Kirchner et al. 2015). Kleijn et al. (2011) argue that conservation strategies should be monitored within the context of land use in the wider countryside. Gottschalk et al. (2007), Whittingham et al. (2007) and Batary et al. (2011) propose that agri-environmental programmes should consider the nature of the landscapes of the regions in which they are implemented, as well as the type of species groups they target. Properly designed biodiversity and environmental indicators can serve as urgently needed tools to support the planning and evaluation of policy measures (Walpole et al. 2009; Vackár et al. 2012). Our understanding of how different levels of land use intensity affect biodiversity is still limited and often restricted to the field scale. Biodiversity analysis with a landscape perspective including the whole land use intensity spectrum and considering different spatial scales could promote a better understanding of these complex interrelationships (Walz and Syrbe 2013).
The present study investigated the relationship between anthropogenic land use characteristics and bird species richness in cultural landscapes. Wu (2010) discussed different definitions of cultural landscapes from a landscape ecology perspective and emphasized the usefulness of the concept, especially when used in the context of a landscape modification gradient. We use the term “cultural landscapes” for organically evolved landscapes, resulting from the interaction between man and his natural environment (UNESCO 1996) including urbanised landscape. While agricultural intensification has been identified as the main driver in the decline of farmland birds (Donald et al. 2001, 2006), only few studies have focused on the general relation between land use intensity (ranging from natural habitats to urbanised landscapes) and bird species diversity (e.g. Loss et al. 2009). Advancing the understanding of this relationship on different spatial scales (from local to regional) was a major goal of our study. We used Austria-wide breeding bird census data to analyse the influence of land use-related landscape characteristics on total avian richness and on species groups with characteristic habitat use (farmland species, forest species, and other species). The environmental indicator distance to nature (D2N) as described by Rüdisser et al. (2012) served as a measure of the degree of habitat change caused by anthropogenic land use. To analyse the relationship between avian diversity and landscape characteristics (altitude, forest proportion, patch density, land cover diversity, and D2N) we applied a generalised linear mixed model (Zuur et al. 2009).
Methods
Bird species data
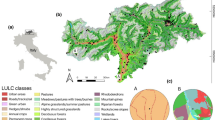
Data on breeding bird species were provided by the Common Bird Monitoring Programme (CBM) of BirdLife Austria for the year 2008. The Austrian CBM data contribute to the Pan-European Common Bird Monitoring Scheme that uses large-scale and long-term monitoring data to measure mean population changes in breeding bird populations across Europe (Gregory et al. 2005). The bird surveys were carried out twice in a predetermined 15-day period during the reproductive period at fixed observation points (n = 2693) along transects (n = 206) within the cultural landscape in Austria (Fig. 1). Surveys were conducted by expert volunteers using the point count method (Bibby et al. 2000). At each point count all visually and acoustically detected birds were recorded. Observations were performed for 5 min and started 2–3 min after the observer arrived at each observation point. The number of observation points per transect ranged between seven and 20, with a median of 12. The 1st and 3rd quartile was 10 and 15, respectively. No surveys were conducted on rainy or strong wind days. The observer always visited the same transects. For further information about the Austrian Bird Monitoring Programme, see Teufelbauer (2010).
To check whether the transects used for the Austrian CBM were representative of the cultural landscape Frühauf and Teufelbauer (2006, 2008) compared the count points with points randomly distributed over the cultural landscape in Austria. They used CORINE land cover data (EEA—European Environment Agency 2007) and GIS data from the Austrian Integrated Administration and Control System—used to administer benefits payments made to farmers under the Common Agricultural Policy—to calculate proportions of land cover types (forest, settlements, water etc.) as well as types of agricultural use (arable, grassland, wine, etc.) within 200 m of each observation point. Furthermore, the most common sub-categories (e.g., maize, summer and winter cereals, set-aside, hay meadows cut once or twice a year, pastures) and agri-environmental schemes (e.g., organic farming, special conservation measures, integrated production) were considered. Differences in distribution with respect to land cover types or types of agricultural use between observation points and randomly chosen points were not statistically significant (Frühauf and Teufelbauer 2008).

Localisation of the representative transects (n = 206) in Austria
Our analysis used the original CBM dataset (ALL), which includes all recorded species (n = 175) and three species groups defined by the European Bird Census Council (EBCC; http://www.ebcc.info). These groups have been used since 2007 for calculation of national and Pan-European common bird indices (Voříšek et al. 2008; EEA—European Environment Agency 2009). The species were selected on the basis of predominant regional habitat use and include 20 characteristic farmland species (FARM), 37 characteristic forest species (FOREST), and 49 other species (OTHER) (EBCC 2012). For the list of species and their classification, see Appendix 1 in supplementary material. The number of species detected at each point is termed species richness (cf. Gotelli and Colwell 2001) and was estimated for all species habitat groups separately.
Gradient of land use intensity and other landscape characteristics
As a measure of land use intensity and spatial land use characteristics we used the composite index D2N developed by Rüdisser et al. (2012). This index is based on a combination of the indicators degree of naturalness and distance to natural habitat and is an appropriate measure of the degree of habitat change caused by anthropogenic land use. Based on a high-resolution naturalness map Rüdisser et al. (2012) produced a spatially comprehensive indicator map. This map was used to estimate D2N at the CBM observation points. D2N is a continuous variable ranging from 0 to 1 and can be used to classify landscape types as proposed: (a) natural or near-natural landscapes (0 < D2N ≤ 0.06, n = 959): these are landscapes where natural land cover is dominant. Predominating ecosystems might be altered, but correspond with the naturally expected; (b) extensively cultivated landscapes with a substantial amount of natural elements (0.06 < D2N ≤ 0.25, n = 1020): these are agriculturally cultivated landscapes with a high degree of natural or near-natural landscape elements such as natural forests (at least 50 %); (c) cultivated landscapes (0.25 < D2N ≤ 0.35, n = 267): agriculturally intensively used landscapes with a substantial extension of natural habitats; (d) intensively cultivated landscapes (0.35 < D2N ≤ 0.65, n = 382): these are agriculturally intensively used landscapes with no or a very small amount of natural habitats; (e) urbanised landscapes (0.65 < D2N ≤ 1, n = 65): landscapes with a high degree of soil sealing (more than 50 %) (Rüdisser et al. 2012). For all species with more than ten sightings we analysed the landscape type preference by calculating mean D2N using all counting points at which the specific species occurred.
To control for the influence of known confounding factors affecting bird richness on the landscape scale (Böhning-Gaese 1997; Farina 1997; Atauri and de Lucio 2001; Desrochers et al. 2011; Bar-Massada et al. 2012) we used altitude (altitude), patch density (PD), land cover diversity (LCD) and percentage of forest coverage (forest) as additional variables. Altitude at count point was estimated using a high-resolution digital terrain model (10 m). The landscape metrics were calculated from a digital land use and land cover (LULC) map for Austria (Rüdisser and Tasser 2011). As this raster map with a cell size of 25 m combines a set of different land use and land cover information, we will subsequently use the term LULC when referring to this dataset. Patch density delineates the number of distinctive LULC patches per unit area, and land cover diversity describes the number of different LULC types per area (McGarigal and Marks 1995). All metrics were calculated for the area surrounding each observation point. To evaluate whether the relationship between landscape characteristics and species richness was sensitive to the applied scale, we estimated D2N, PD, forest and LCD for a radius of 100, 500, 1000 and 5000 m around each observation point.
Austria and hence our study region contains the Ecoregions Alps conifer and mixed forests; Western European Broadleaf forests, Central European mixed forests, and Pannonian mixed forests (Olson et al. 2001). These ecoregions were added as fixed effects to avoid a possible omitted variable bias. All explanatory variables were checked for multicollinearity using the variance inflation factor (VIF).
Modelling the influence of landscape characteristics on species richness
Data analysis aimed to understand and describe the relationship between bird species richness and landscape characteristics. It is reasonable to assume that the species richness of different habitat groups might follow different trends (Gregory et al. 2005), and therefore we divided bird communities into the groups described above (FOREST, FARM and OTHER). As species richness is a count variable, the relationship between landscape characteristics and species richness was investigated using count data models. Due to a considerable number of zeros a hurdle model was considered an appropriate model approach. Compared to zero-inflated models, the hurdle model assumes that zero observations are all true non-occurrences. That is, a zero observation is recorded because the species does not occur at a site, either due to its habitat characteristics or because it does not saturate its entire suitable habitat by chance (Martin et al. 2005). In contrast, a false-zero occurs when the observer fails to detect a species that occurs at a site. Ease of detectability (easy to recognize and easy to monitor) is one of the selection criteria for the common bird indicator (see “Bird species data” section). Furthermore, a zero observation in our case would mean that not a single bird of any possible species (FOREST = 37, FARM = 20 or OTHER = 49) was detected on either one of the two visits. Hence, we assume that the number of false-zeros in our study is negligible. Additionally, it makes sense to allow different data-generating processes (DGPs) in order to determine whether, on the one hand, the habitat is at all suitable for specific bird groups and, on the other hand, for determining which factors favour an increase in species richness. Although the same variables were used for both DGPs, the variables may very differently influence species richness or the presence of a species group (cf. Cunningham and Lindenmayer 2005; Huggett 2005; Radford et al. 2005). The multilevel structure of our data (counting points are aligned along a transect) required the use of a random effects hurdle model. The random intercept accounted for the dependence of the observation units within one transect in order to obtain valid estimates.
To confirm the statistical suitability of the hurdle model, we not only worked from the theoretical considerations mentioned above but we estimated further count data models also involving zero-inflated models. The model employing the Poisson distribution (‘Poisson model’) is commonly applied with count data. However, a characteristic of the Poisson distribution is the fact that the mean is the same as the variance, i.e. equidispersion is assumed and modelling over-dispersion, often observed in practice, is not possible. In order to overcome this restriction the negative-binomial distribution was employed to allow different estimates of the variance and the mean (‘negative-binomial model’). When dealing with count data more zeros may be observed than predicted by the Poisson or negative-binomial distribution. When this is the case the excess of zeros may be modelled either with a zero-inflated model or with a hurdle model, depending on the assumptions underlying the data-generating process of the zeros. Both types of model (each assuming either the Poisson or the negative-binomial distribution of the outcome) were developed to cope with zero-inflated outcome data with over-dispersion (negative-binomial distribution) or without (Poisson distribution). For zero-inflated models (‘zero-inflated Poisson model’ or ‘zero-inflated negative-binomial model’) theory suggests that the zeros are generated by two independent processes, whereas the hurdle model (‘hurdle model’) assumes that all zero data are from one structural source as described above.
For model evaluation information criteria (Schwarz 1978; Akaike 1981), the log-likelihood ratios, deviances, the squared correlation between the observed species richness, and the estimated values (R2), McFadden’s Pseudo Rsquare (McFadden 1973) and residual diagnostics were applied. As theoretical reasons and statistical arguments favour the hurdle model, we now describe this model in more detail. The hurdle model consists of two parts: a left-truncated count component and a right-censored hurdle component.
The first part is a binary (presence/absence) logit model:
where p is the probability of an event occurring given x, \(\left( {\frac{p}{1 - p}} \right)\) is the odds of species presence given x, \(x_{k}\) is the kth independent variable, \(b_{k}\) the kth fixed regression coefficient, \(k = 1, \ldots ,K\) and \(b_{0}\) the random intercept (Zuur et al. 2009). The parameters in this part of the model can be interpreted in terms of odds ratios and standardized variable changes. For a standard deviation change in \(x_{k}\) (\(s_{k}\)), the odds are expected to change by a factor of \(\exp (b_{k} s_{k} )\), with all other variables remaining constant:
The second part is a count model using the negative binomial distribution, \(E[y|x] = \exp (xb)\). Again, a standardized variable change in the variable \(x_{k}\) can be computed as \(\exp (b_{k} s_{k} )\), where \(s_{k}\) is the standard deviation of variable \(x_{k}\).
In order to obtain the maximum likelihood estimates the log-likelihood function is maximized. To estimate the generalised linear mixed models (GLMM) the package glmm.admb (Skaug et al. 2013) of the statistical software R was used (Bolker 2008). The covariance matrix was obtained as the numerically determined Hessian matrix and the significance of the regression coefficients was assessed via Wald tests.
The models were estimated for each species subset and all four spatial scales (100, 500, 1000 and 5000 m around each counting point). Statistical analysis was performed in R 3.0.1; calculation of landscape metrics and all related data handling was conducted in ArcGIS 9.3.
Results
The 2008 monitoring counts detected 175 bird species. Avian species richness at observation points ranged from 1 to 29 with an average of 10.5 and a standard deviation (SD) of 4.7 considering all recorded species. Chaffinch (Fringilla coelebs) showed the highest frequency of occurrence, occurring in 1919 of 2693 sampling points (71.3 %), followed by Blackcap (Sylvia atricapilla) with 61.1 % and Common Blackbird (Turdus merula) with 55.7 %. All other species had a relative frequency of occurrence of less than 50 %.
D2N at observation points comprised the whole possible range from 0 to 1 with an average of 0.18 (SD 0.17). Mean D2N values for the 100 m radius for bird species ranged from 0.05 (0.04 SD) (Ficedula albicollis) to 0.47 (SD 0.18) (Perdix perdix). All species from the group FARM achieved mean D2N values above 0.2, and all species from the group FOREST occurred at points with a mean D2N below 0.16. For details, see Appendix 1 in supplementary material.
Separate analysis of the species groups with predominant habitat use (FOREST, FARM or OTHER) revealed that species richness of those groups responded differently to land use intensity. While species richness of the FOREST group decreased with increasing land use intensity, FARM species richness increased (Table 1). This characteristic had to be considered in the model, and therefore the relationship between bird species richness and landscape characteristics for each habitat group was modelled separately.
Variance inflation factors (VIFs) revealed no relevant multicollinearity between the explanatory variables (Table 2) on any scale (100, 500, 1000, 5000 m). VIFs were smaller than 4.2 for variables on all scales. For the 100 scale model VIFs were even smaller than 2.5 for all variables. The statistical analysis of all estimated models (Poisson model, negative-binomial model, zero-inflated Poisson model, and zero-inflated negative-binomial model) revealed that the mixed-effects hurdle model performed best on all scales and for all species groups. However, we found a remarkably high consistency amongst models regarding which variables had a statistically significant impact and the size of the coefficient estimates (results are available from the authors on request).
The hurdle model (Table 3) showed a moderate but consistent relationship between avian species richness and the index D2N. Models on all scales and for all species subsets except the subset FARM revealed a negative impact of D2N on species richness. The species group FARM showed a negative dependence between D2N and species richness for the count model part of the hurdle model, but a positive dependence for the binary (presence/absence) logit models on all scales. On the 500 m scale the influence of D2N on FARM species richness was not significant (Table 3). This apparent contradiction can be explained by the fact that FARM species rarely occurred in forested habitats, but mostly in an agriculturally used and thus open landscape (with an average D2N > 0.2). Nevertheless, further land use intensification and therefore an increase in D2N did not cause an increase in FARM species richness.
Species richness decreased with increasing altitude. This relationship was consistent for all species subsets. Influence of LCD on species richness was small and mostly not significant. An increase in PD was associated with an increased richness of FOREST and OTHER species and a decreased richness of FARM species. Increased forest cover was associated with increased FOREST species richness, but decreased FARM species richness. Here the species group OTHER reacted ambiguously.
The results achieved with the models demonstrate that D2N is an important explanatory variable for bird richness. Especially the richness of the FOREST species group was strongly influenced by land use intensity. An increase in D2N by one standard deviation changed the expected species richness by a factor of 0.65 (in the 100 m radius). This effect is the partial effect of D2N as the effects of all other variables (altitude, patch density, land cover diversity and ratio of forest coverage) were partialed out (Table 4).
Discussion
Evaluating the influence of land use and cover on biodiversity on the landscape scale is a very challenging task. This is not only due to the inherent complexity of interactions in time and on different spatial scales, but also to the fact that data about biodiversity are and always will be limited and incomplete. This is also true for the study at hand. The use of data from only one year could be seen as such a drawback as annual changes in population dynamics might have influenced the results. We analysed bird data from 2008 to ensure temporal congruence with the LULC data set that incorporates detailed and large-scale agricultural land use data from the same year. The bird observation data from 2008 generally correspond with data from the preceding 10-year period from 1998 to 2008 (Teufelbauer 2010). As we focused on the influence of landscape characteristics on common bird diversity and not on rare or specialised species and considering the geographically wide distribution and large number of observation points, we are convinced that possible seasonal population effects did not exert a systematic influence on the results. The focus on common bird species, which obviously account for just a fraction of overall bird diversity, and the restriction to species richness instead of abundance-related indicators can also be seen as limitations. These restrictions were deliberately chosen to reduce the effects of stochastic events as well as the influence of various observers (Nichols et al. 2000).
Species richness is very often used as a measure of community and regional diversity, not only as a basis for many ecological models but also in the context of conservation efforts and studies evaluating changes within ecosystems (Gotelli and Colwell 2001). A drawback to using species richness as a measure of biodiversity is that it may not detect changes in the species composition of a community (Koch et al. 2011). We attempted to account for this not only by analysing species richness of various individual avian species groups, but also by estimating mean D2N values for each species (Appendix 1 in supplementary material). The fact that all species from the group FARM occurred at points with an average D2N value above 0.2 and all species from the group FOREST occurred at points with an average D2N below 0.16 confirms the soundness of these habitat groups selected by the European Bird Census Council (European Bird Census Council 2012) and emphasizes the importance of utilising multifaceted indicators in the context of biodiversity monitoring and decision-making support.
The results of the hurdle model show that increased land use intensity as measured by D2N is generally accompanied by a decrease in bird species richness. This is in accordance with Rüdisser et al. (2012), who claimed that D2N can be used as a surrogate for land use-related anthropogenic influence on biodiversity. As we analysed the whole range of land use intensity types (from natural habitats to highly urbanised areas), it was especially important to account for bird species groups with distinctive habitat preferences. If analysis had been based on total avian species richness, models would have had a poor predictive power because different habitat preferences of FOREST and FARM species might have masked the effect of land use characteristics. This might have happened to Schindler et al. (2013), who reported poor performance of landscape metrics as indicators for avian species richness. As expected, FOREST species richness increased with an increase in the amount of forested area, but also benefited from low land use intensity. FARM species are open habitat species, which is why it is not surprising that an increasing amount of forested area was accompanied by a decrease in FARM species richness. The mean D2N values of observation points where the FARM species occurred (Appendix 1 in supplementary material) demonstrate that FARM species rarely occurred at observation points with very low D2N values. Very low D2N values in the study region were mainly achieved in naturally forested areas. Nevertheless, the count model part of the hurdle model showed that increased land use intensity (D2N)—considering the amount of forest as a covariable—caused a decrease in FARM species richness. As FARM species depend on open habitats, they profit from extensive land use in areas where forests are the natural land cover type, but not from further land use intensification. Here, use of the hurdle model assists ecological understanding in a case where the reason for the presence of a species group might differ from the underlying reason affecting species diversity (c.f. Cunningham and Lindenmayer 2005).
Our results support the assumptions made by Desrochers et al. (2011), who investigated how the conversion of natural area to human-dominated land cover affects avian species richness in Ontario, Canada. They found that conversion of up to 44 % of natural habitats to human-dominated land cover can result in increased avian richness, because many open habitat species can benefit from such changes while only few forest-obligate species are lost. Total species richness was consistent with the sum of the species-area curves for natural habitat species and human-dominated habitat species (Desrochers et al. 2011). This argumentation is in line with Salek et al. (2010), who observed increased bird richness along forest edges and forest roads as compared to forest interior. As the forests in the study region in Central Europe were dominated by spruce plantations and increasing diversity of tree species also positively affected bird richness, Salek et al. (2010) assumed that margins of low-traffic roads might provide additional habitat resources in structurally poor forests. Interestingly, our analysis also shows a positive relationship between patch density and forest species, but a negative relationship with farmland species. This is in accordance with Filippi-Codaccioni et al. (2010), who hypothesised that habitat diversity caused by vertical features enhancement could negatively affect farmland specialist birds, because some of them need relatively large open landscapes and can be harmfully affected by habitat heterogeneity caused by trees and bushes.
Performance of landscape metrics was best at a close range around observation points. This is in accordance with Schindler et al. (2013), who investigated the relations between 52 landscape metrics and species richness of different taxa (woody plants, orchids, orthopterans, amphibians, reptiles, and small terrestrial birds) and showed that species richness of birds was better indicated by landscape metrics at a close range of 20–50 ha. Besides the fact that model performance was best at the 100 and 500 m radius, our results indicate that the relation between the investigated landscape characteristics and bird richness reacted relatively robustly to changes in the analysis scale.
Conclusion
Effective future protection of biodiversity will not only require an honest and tremendous political will, but also accurate methods to give conservationists and policy makers a sound basis for making efficient and informed management decisions. Even within science biodiversity is often reduced to species diversity (Haber 2008; Walz and Syrbe 2013), but the description of biodiversity with all its dimensions and facets definitely cannot be reduced to a few indicators. Different protection aims need different approaches and indicators. In our work we focused on the relation between land use characteristics and diversity of common breeding birds, because bird indicators are being increasingly used as a measure of environmental health and for evaluation and monitoring purposes. The comparatively broad data availability on breeding birds makes bird indicators very valuable tools (Gregory and van Strien 2010). Nevertheless, these indicators have to be interpreted with care. As we demonstrate, results can be influenced by the species set chosen and possible effects can be obscured if a species set includes species with oppositional reaction to environmental changes. Furthermore, the superior mobility of birds in general and the pronounced ability of some species to adapt their habitat needs to changing environmental conditions might weaken the sensitivity of breeding bird indicators to detect general biodiversity changes. Additional data and indicators based on other taxa are urgently needed to better understand, measure, and monitor biodiversity changes caused by anthropogenic land use.
References
Akaike H (1981) Likelihood of a model and information criteria. J Econom 16:3–14
Atauri JA, de Lucio JV (2001) The role of landscape structure in species richness distribution of birds, amphibians, reptiles and lepidopterans in Mediterranean landscapes. Landscape Ecol 16:147–159
Bar-Massada A, Wood EM, Pidgeon AM, Radeloff VC (2012) Complex effects of scale on the relationships of landscape pattern versus avian species richness and community structure in a woodland savanna mosaic. Ecography 35:393–411
Batary P, Baldi A, Kleijn D, Tscharntke T (2011) Landscape-moderated biodiversity effects of agri-environmental management: a meta-analysis. Proc R Soc B 278:1894–1902
Baudron F, Giller KE (2014) Agriculture and nature: trouble and strife? Biol Conserv 170:232–245
Benton TG, Vickery JA, Wilson JD (2003) Farmland biodiversity: is habitat heterogeneity the key? Trends Ecol Evol 18:182–188
Bibby C, Burgess N, Hill D, Mustoe S (2000) Bird census techniques. Academic Press, London
Böhning-Gaese K (1997) Determinants of avian species richness at different spatial scales. J Biogeogr 24:49–60
Bolker BM (2008) Ecological models and data in R. Princeton University Press, Princeton
Butler SJ, Brooks D, Feber RE, Storkey J, Vickery JA, Norris K (2009) A cross-taxonomic index for quantifying the health of farmland biodiversity. J Appl Ecol 46(6):1154–1162
Concepción ED, Díaz M, Baquero R (2008) Effects of landscape complexity on the ecological effectiveness of agri-environment schemes. Landscape Ecol 23:135–148
Concepción ED, Díaz M, Kleijn D, Báldi A, Batáry P, Clough Y, Gabriel D, Herzog F, Holzschuh A, Knop E, Marshall EJP, Tscharntke T, Verhulst J (2012) Interactive effects of landscape context constrain the effectiveness of local agri-environmental management. J Appl Ecol 49:695–705
Culman SW, Young-Mathews A, Hollander AD, Ferris H, Sanchez-Moreno S, O’Geen AT, Jackson LE (2010) Biodiversity is associated with indicators of soil ecosystem functions over a landscape gradient of agricultural intensification. Landscape Ecol 25:1333–1348
Cunningham RB, Lindenmayer DB (2005) Modeling count data of rare species: some statistical issues. Ecology 86:1135–1142
Desrochers RE, Kerr JT, Currie DJ (2011) How, and how much, natural cover loss increases species richness. Glob Ecol Biogeogr 20:857–867
Donald PF, Green RE, Heath MF (2001) Agricultural intensification and the collapse of Europe’s farmland bird populations. Proc R Soc B 268:25–29
Donald PF, Sanderson FJ, Burfield IJ, van Bommel FPJ (2006) Further evidence of continent-wide impacts of agricultural intensification on European farmland birds, 1990–2000. Agric Ecosyst Environ 116:189–196
Dullinger S, Essl F, Rabitsch W, Erb K, Gingrich S, Haberl H, Hülber K, Jarošík V, Krausmann F, Kühn I, Pergl J, Pyšek P, Hulme PE (2013) Europe’s other debt crisis caused by the long legacy of future extinctions. PNAS 110:7342–7347
European Environment Agency (EEA) (2007) CLC2006 technical guidelines. Technical report, Luxembourg
European Environment Agency (EEA) (2009) Progress towards the European 2010 biodiversity target. EEA Technical report 4. Office for Official Publication of the European Communities, Luxembourg
European Bird Census Council (2012) Report on the Pan-European common bird monitoring scheme. June 2012. http://www.ebcc.info. Accessed July 2014
Farina A (1997) Landscape structure and breeding bird distribution in a sub-Mediterranean agro-ecosystem. Landscape Ecol 12:365–378
Filippi-Codaccioni O, Devictor V, Bas Y, Julliard R (2010) Toward more concern for specialisation and less for species diversity in conserving farmland biodiversity. Biol Conserv 143:1493–1500
Frühauf J, Teufelbauer N (2006) Evaluierung des Einflusses von ÖPUL-Maßnahmen auf Vögel des Kulturlandes anhand von repräsentativen Monitoring-Daten: Zustand und Entwicklung: Studie von BirdLife Österreich für die ÖPUL-Halbzeit-Evaluierung (update) im Auftrag des BMLFUW, Wien
Frühauf J, Teufelbauer N (2008) Preparation of the Austrian Farmland bird index. Pilot study. Report on behalf of the Austrian Federal Ministry of Agriculture, Forestry, Environment and Water Management, Wien
Gonthier DJ, Ennis KK, Farinas S, Hsieh HY, Iverson AL, Batáry P, Rudolphi J, Tscharntke T, Cardinale BJ, Perfecto I (2014) Biodiversity conservation in agriculture requires a multi-scale approach. Proc R Soc B 281:20141358
Gotelli NJ, Colwell RK (2001) Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecol Lett 4:379–391
Gottschalk TK, Diekoetter T, Ekschmitt K, Weinmann B, Kuhlmann F, Purtauf T, Dauber J, Wolters V (2007) Impact of agricultural subsidies on biodiversity at the landscape level. Landscape Ecol 22:643–656
Gregory RD, van Strien A (2010) Wild bird indicators: using composite population trends of birds as measures of environmental health. Ornithol Sci 9:3–22
Gregory RD, van Strien A, Vorisek P, Meyling AWG, Noble DG, Foppen RPB, Gibbons DW (2005) Developing indicators for European birds. Phil Trans R Soc B 360:269–288
Haber W (2008) Biological diversity a concept going astray? GAIA—Ecol Perspect Sci Soc 17(Supplement 1):91–96
Huggett AJ (2005) The concept and utility of ‘ecological thresholds’ in biodiversity conservation. Biol Conserv 124:301–310
Kirchner M, Schmidt J, Kindermann G, Kulmer V, Mitter H, Prettenthaler F, Rüdisser J, Schauppenlehner T, Schönhart M, Strauss F, Tappeiner U, Tasser E, Schmid E (2015) Ecosystem services and economic development in Austrian agricultural landscapes—the impact of policy and climate change scenarios on trade-offs and synergies. Ecol Econ 109:161–174
Kleijn D, Rundlöf M, Scheper J, Smith HG, Tscharntke T (2011) Does conservation on farmland contribute to halting the biodiversity decline? Trends Ecol Evol 26:474–481
Koch AJ, Drever MC, Martin K (2011) The efficacy of common species as indicators: avian responses to disturbance in British Columbia, Canada. Biodivers Conserv 20:3555–3575
Krebs JR, Wilson JD, Bradbury RB, Siriwardena GM (1999) The second silent spring? Nature 400(6745):611–612
Loss SR, Ruiz MO, Brawn JD (2009) Relationships between avian diversity, neighborhood age, income, and environmental characteristics of an urban landscape. Biol Conserv 142:2578–2585
Martin TG, Wintle BA, Rhodes JR, Kuhnert PM, Field SA, Low-Choy SJ, Tyre AJ, Possingham HP (2005) Zero tolerance ecology: improving ecological inference by modelling the source of zero observations. Ecol Lett 8:1235–1246
McFadden D (1973) Conditional Logit analysis of qualitative choice behavior. In: Zarembka P (ed) Frontiers in econometrics. Academic Press, New York, pp 105–142
McGarigal K, Marks B (1995) FRAGSTATS: spatial pattern analysis program for quantifying landscape structure: U.S. Forest Service General Technical Report, Portland, OR
Nichols JD, Hines JE, Sauer JR, Fallon FW, Fallon JE, Heglund PJ (2000) A double-observer approach for estimating detection probability and abundance from point counts. Auk 117:393–408
Olson DM, Dinerstein E, Wikramanayake ED, Burgess ND, Powell GV, Underwood EC, D’Amico JA, Itoua I, Strand HE, Morrison JC, Loucks CJ, Allnutt TF, Ricketts TH, Kura Y, Lamoreux JF, Wettengel WW, Hedao P, Kassem KR (2001) Terrestrial ecoregions of the worlds: a new map of life on Earth. Bioscience 51:933–938
Pimentel D, Stachow U, Takacs DA, Brubaker HW, Dumas AR, Meaney JJ, O’Neil JAS, Onsi DE, Corzilius DB (1992) Conserving biological diversity in agricultural/forestry systems. Bioscience 42:354–362
Radford JQ, Bennett AF, Cheers GJ (2005) Landscape-level thresholds of habitat cover for woodland-dependent birds. Biol Conserv 124:317–337
Rüdisser J, Tasser E (2011) Landbedeckung Österreichs—Datenintegration und Modellierung. In: Strobl J, Blaschke T, Griesebner G (eds) Angewandte Geoinformatik 2011: Beiträge zum 23. AGIT-Symposium Salzburg, pp 579–588
Rüdisser J, Tasser E, Tappeiner U (2012) Distance to nature—a new biodiversity relevant environmental indicator set at the landscape level. Ecol Indic 15:208–216
Sala OE, Chapin FS, Armesto JJ, Berlow E, Bloomfield J, Dirzo R, Huber-Sanwald E, Huenneke LF, Jackson RB, Kinzig A, Leemans R, Lodge DM, Mooney HA, Oesterheld M, Poff NL, Sykes MT, Walker BH, Walker M, Wall DH (2000) Biodiversity—global biodiversity scenarios for the year 2100. Science 287:1770–1774
Salek M, Svobodova J, Zasadil P (2010) Edge effect of low-traffic forest roads on bird communities in secondary production forests in central Europe. Landscape Ecol 25:1113–1124
Schindler S, von Wehrden H, Poirazidis K, Wrbka T, Kati V (2013) Multiscale performance of landscape metrics as indicators of species richness of plants, insects and vertebrates. Linking landscape structure and biodiversity. Ecol Indic 31:41–48
Schwarz G (1978) Estimating the dimension of a model. Ann Stat 6:461–464
Skaug H, Fournier D, Nielsen A, Magnusson A, Bolker B (2013) Generalized linear mixed models using AD model builder: R package version 0.7.5
Tasser E, Sternbach E, Tappeiner U (2008) Biodiversity indicators for sustainability monitoring at municipality level: an example of implementation in an alpine region. Ecol Indic 8:204–223
Teufelbauer N (2010) Der Farmland Bird Index für Österreich—erste Ergebnisse zur Bestandsentwicklung häufiger Vogelarten des Kulturlandes: the Farmland Bird Index for Austria—first results of the changes in populations of common birds of farmed land. Egretta 51:35–50
Tscharntke T, Klein AM, Kruess A, Steffan-Dewenter I, Thies C (2005) Landscape perspectives on agricultural intensification and biodiversity—ecosystem service management. Ecol Lett 8:857–874
Tscharntke T, Tylianakis JM, Rand TA, Didham RK, Fahrig L, Batáry P, Bengtsson J, Clough Y, Crist TO, Dormann CF, Ewers RM, Fründ J, Holt RD, Holzschuh A, Klein AM, Kleijn D, Kremen C, Landis DA, Laurance W, Lindenmayer D, Scherber C, Sodhi N, Steffan-Dewenter I, Thies C, Van der Putten WH, Westphal C (2012) Landscape moderation of biodiversity patterns and processes—eight hypotheses. Biol Rev 87:661–685
UNESCO (United Nations Educational S, and Cultural Organization) (1996) Operational guidelines for the implementation of the world heritage convention. UNESCO, Paris. http://whc.unesco.org/archive/opguide05-annex3-en.pdf
Vackár D, ten Brink B, Loh J, Baillie JEM, Reyers B (2012) Review of multispecies indices for monitoring human impacts on biodiversity. Ecol Indic 17:58–67
Voříšek P, Klvaňová A, Wotton S, Gregory RD (eds) (2008) A best practice guide for wild bird monitoring schemes. Czech Republic, Třeboň
Walpole M, Almond REA, Besancon C, Butchart SHM, Campbell-Lendrum D, Carr GM, Collen B, Collette L, Davidson NC, Dulloo E, Fazel AM, Galloway JN, Gill M, Goverse T, Hockings M, Leaman DJ, Morgan DHW, Revenga C, Rickwood CJ, Schutyser F, Simons S, Stattersfield AJ, Tyrrell TD, Vie J, Zimsky M (2009) Tracking progress toward the 2010 biodiversity target and beyond. Science 325:1503–1504
Walz U, Syrbe R (2013) Linking landscape structure and biodiversity. Ecol Indic 31:1–5
Wehrden HV, Abson D, Beckmann M, Cord A, Klotz S, Seppelt R (2014) Realigning the land-sharing/land-sparing debate to match conservation needs: considering diversity scales and land-use history. Landscape Ecol 29:941–948
Whittingham MJ, Krebs JR, Swetnam RD, Vickery JA, Wilson JD, Freckleton RP (2007) Should conservation strategies consider spatial generality? Farmland birds show regional not national patterns of habitat association. Ecol Lett 10:25–35
Wilson JD, Evans AD, Grice PV (2010) Bird conservation and agriculture: a pivotal moment? Ibis 152:176–179
Wretenberg J, Part T, Berg A (2010) Changes in local species richness of farmland birds in relation to land-use changes and landscape structure. Biol Conserv 143:375–381
Wright HL, Lake IR, Dolman PM (2012) Agriculture—a key element for conservation in the developing world. Conserv Lett 5:11–19
Wu J (2010) Landscape of culture and culture of landscape: does landscape ecology need culture? Landscape Ecol 25:1147–1150
Zimmermann P, Tasser E, Leitinger G, Tappeiner U (2010) Effects of land-use and land-cover pattern on landscape-scale biodiversity in the European Alps. Agric Ecosyst Environ 139:13–22
Zuur FA, Ienon E, Walker N, Savelier AA, Smith GM (2009) Mixed effects models and extensions in ecology with R. Springer, New York
Acknowledgments
This article was funded by the Austrian Climate Research Fund project ‘CAFEE-Climate change in agriculture and forestry: an integrated assessment of mitigation and adaptation measures in Austria’ as well as the collaborative research programme proVISION of the Austrian Federal Ministry of Science under Research Contract 100394. Ulrike Tappeiner is a member of the research area ‘Alpine Space—Man and Environment’ at the University of Innsbruck. Special thanks go to all the volunteer observers of the Austrian Common Bird Monitoring Scheme and Mag. Ingeborg Fiala of the Austrian Federal Ministry of Agriculture, Forestry, Environment and Water Management (BMLFUW), who prompted this analysis. Finally, we wish to thank the editor Teresa Pinto-Correia and an anonymous reviewer for their very helpful comments.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Rüdisser, J., Walde, J., Tasser, E. et al. Biodiversity in cultural landscapes: influence of land use intensity on bird assemblages. Landscape Ecol 30, 1851–1863 (2015). https://doi.org/10.1007/s10980-015-0215-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-015-0215-3