
Abstract
Ecological theory predicting the impact of fire on ecological communities is typically focused on post-disturbance recovery processes or on disturbance-diversity dynamics. Yet the established relationship between vegetation structure and animal diversity could provide a foundation to predict the short-term effects of fire on biodiversity, but has rarely been explored. We tested the hypothesis that fire effects on bird assemblages would be moderated by increasing vegetation structure. We examined bird assemblages in burnt and unburnt sites at 1 and 6 years after a wildfire, and compared richness and composition responses among and within three structurally distinct vegetation types in the same landscape: heath, woodland and forest. We found that short-term changes in bird assemblage composition were largest in simple heath vegetation and smallest in complex forest vegetation. The short-term change in species richness was larger in forest than in heath. We also found that among-site assemblage variability was greater shortly after fire in heath and woodland vegetation compared with forest vegetation. Our results indicate that complexity in vegetation structure, particularly overstorey cover, can act as an important moderator of fire effects on bird assemblages. Mechanisms for this response include a greater loss of structure in vegetation characterised by a single low stratum, and a proportionally greater change in bird species composition despite a smaller absolute change in species richness. We discuss our results in the context of a new conceptual model that predicts contrasting richness and composition responses of bird assemblages following disturbance along a gradient of increasing vegetation structure. This model brings a different perspective to current theories of disturbance, and has implications for understanding and managing the effects of fire on biodiversity in heterogeneous landscapes.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Understanding the effects of fire in heterogeneous landscapes is of significant interest to ecologists and land managers (Turner 1987; Kerby et al. 2007; Faivre et al. 2011; Pastro et al. 2011; Lindenmayer et al. 2014), but accurate prediction of the impacts of fire on animal assemblages is difficult. Much of the theory underpinning fire ecology has centred on patterns of recovery, such as succession and post-fire recolonisation (Franklin et al. 2000; Kelly et al. 2011; Watson et al. 2012). Other theory has been developed to understand how spatial and temporal patterns of fire can influence biodiversity, including the patch-mosaic concept (Pickett and White 1985; Bradstock et al. 2005; Parr and Andersen 2006) and the intermediate disturbance hypothesis (Connell 1978; Westgate et al. 2012). This ecological theory has informed many areas of fire ecology, but there remains much to learn about fire impacts (Bradstock et al. 2012). For example, there is mixed support for the intermediate disturbance hypothesis (Pastro et al. 2011; Fox 2013), and some frameworks for understanding post-disturbance succession lack specificity (Keeley et al. 2006; Lindenmayer et al. 2008b; Smith et al. 2012). Further research is required to develop a better understanding of how fire affects different animal assemblages.
One potential avenue for further research is the integration of vegetation structure into studies on fauna responses to fire. Fire can have dramatic effects on vegetation structure (Fuhlendorf et al. 2006; Santana et al. 2012), and therefore its associated fauna. Vegetation structure is also an important driver of animal species diversity in terrestrial ecosystems worldwide (Bohning-Gaese 1997; Tews et al. 2004). For example, empirical studies have shown that bird species richness increases with the vertical height of vegetation (e.g. MacArthur and MacArthur 1961; Lindenmayer et al. 2008a; Kutt and Martin 2010). Each additional stratum adds substrates for foraging, nesting, and shelter, thus increasing niche availability, facilitating species coexistence, and driving higher diversity. Previous studies that have linked vegetation structure to animal responses to fire, however, have focused on its role in moderating species recovery (Monamy and Fox 2000; Lindenmayer et al. 2008a), and not as a tool to predict short-term impacts.
The heterogeneity of landscapes is strongly influenced by the occurrence of different vegetation types (Forman 1995), and can facilitate or retard the effects of disturbance (Turner 1987). When fire occurs across multiple vegetation types, the differences in structure may be critical to how faunal assemblages are affected (Monamy and Fox 2000; Lindenmayer et al. 2008a; Kutt and Martin 2010). However, most studies have examined the effects of fire on assemblages in a single vegetation type, such as grassland (Coppedge et al. 2008; Collins and Calabrese 2012) or forest (Schimmel and Granstrom 1996; Clavero et al. 2011). These studies can provide valuable insight into how changes in vegetation after fire can alter faunal assemblages (Brotons et al. 2004; Faivre et al. 2011; Pastro et al. 2011; Santana et al. 2012), but provide limited information about the relative impact of fire on animals in different and distinct vegetation types within a landscape.
There are contrasting expectations for how vegetation structure might influence the responses of animals to fire. First, many fire-prone landscapes consist of structurally simple vegetation, such as the savannah grasslands of tropical latitudes that rebound rapidly from fire (Murphy and Bowman 2012). This suggests that the impact of fire might be small in structurally simple vegetation. However, studies have also demonstrated show that complex vegetation prior to a disturbance has greater potential to retain structure following a disturbance (Franklin et al. 2002), and that complex vegetation can be more resilient to disturbance (Lavorel 1999). This suggests that the impact of fire might be limited in more complex vegetation. To the best of our knowledge, there have been no previous studies that have examined these contrasting expectations in a single fire-prone and structurally heterogeneous landscape.
In this investigation, we test the hypothesis that the impact of fire on bird assemblages would be smaller in structurally complex vegetation compared with structurally simple vegetation. Our study landscape allowed for an explicit comparison of bird assemblage responses to fire among distinct vegetation types, including heath, open woodland and tall forest vegetation. This extends the well-established phenomenon that bird assemblages differ in composition among vegetation types, and instead examines how this general pattern might moderate fire impacts. Our test of whether vegetation structure moderates fire effects on species assemblages places an emphasis on the need for explicit consideration of habitat structural attributes to predict where a fire might have the greatest impact. We use our results to develop a conceptual model for exploring the role of vegetation structure in moderating disturbance effects on animal assemblages in heterogeneous landscapes.
Methods
Study area and design
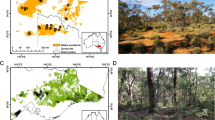
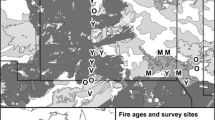
We conducted this study in Jervis Bay Territory, approximately 200 km south of Sydney in south-east Australia (150.70° East, −35.15° South). The Jervis Bay Territory includes Booderee National Park, which covers approximately 7,500 ha (Fig. 1), and is comprised of several vegetation communities (Lindenmayer et al. 2008a), with forest, woodland and heath being the three most widespread vegetation types (Fig. 1). In September 2003, permanent study sites were established across the National Park as part of a landscape-scale monitoring program (Lindenmayer et al. 2008a), with the number of sites in each of the different vegetation types generally proportional to the amount of cover of that vegetation. Each site was 100 m long, marked with a central 100-m transect line, and had permanent markers placed at 0, 20, 40, 60, 80 and 100 m.

A location map a showing the distribution of our survey sites and area burned by the 2003 wildfire in Jervis Bay Territory, south-east Australia. Study sites were established in three distinct vegetation types including a forest, b woodland, and c heath. These vegetation types create a heterogeneous landscape and a wide gradient in structural complexity
In December 2003, an unplanned wildfire burnt approximately 50 % of Booderee National Park, but occurred unevenly across the different vegetation communities (Fig. 1). This provided an opportunity to conduct a ‘natural experiment’ to test the effects of recent fire on animal assemblages across multiple vegetation types. Active and widespread suppression of fire does not occur in Booderee National Park. Therefore, the experimental ‘treatment’ applied to our sites by the 2003 wildfire is compared against a set of ‘unburnt’ sites with a complex fire history. For example, since 1972 the number of fires in any given part of the National Park has varied from none to five (Lindenmayer et al. 2008a), and three of the unburnt sites used in our analyses were burnt once in the 10 years prior to the major 2003 wildfire. Our comparison of burnt with unburnt sites in different vegetation types therefore averages across the fire histories of individual sites, and focuses on the short-term responses of birds to vegetation structural changes as a result of the 2003 fire.
In this study, we selected 56 sites from the three dominant vegetation types: heath (15 burnt sites, 2 unburnt), woodland (12 burnt sites, 7 unburnt) and forest (11 burnt sites, 9 unburnt). Together, these vegetation types cover approximately 70 % of the total vegetative cover of Jervis Bay Territory (Fig. 1, Table S1).
Data collection
We completed bird surveys in September 2004 and 2009, approximately 1 and 6 years after the 2003 wildfire. September is the breeding season for the majority of bird species in Booderee National Park, and when most summer migrants have arrived. For each survey year, we performed two repeat point counts of birds at the 20 and 80 m markers in each site, resulting in four point counts in each site per survey year. We recorded all birds seen or heard within a 50 m radius of the marker during a 5-min period, excluding birds flying overhead. We conducted surveys between dawn and mid-morning. Each site (i.e. both markers) was surveyed on a different day by a different observer to reduce day effects on detection and to overcome potential observer heterogeneity problems (Cunningham et al. 1999). Our survey protocol followed standards that are widely reported in the ecology literature (de Lima et al. 2013; Driscoll and Lindenmayer 2010; Ikin et al. 2013), and helps to correct for false negative errors (i.e. failure to detect a species that is present at the site (Tyre et al. 2003). We pooled surveys at each plot within a site to give one set of observations per site per year.
In December 2004 and 2009, we measured six vegetation structural attributes in two 20 × 20 m plots located between the 20–40 and 60–80 m points in every site. The measures were: estimated percentage cover of overstorey, midstorey and understorey vegetation, as well as percentage cover of grass, leaf litter, and bare ground. We defined overstorey as vegetation over 10 m in height, midstorey between 2 and 10 m, and understorey as less than 2 m in height. Each stratum was assessed independently of the other, meaning that a site could have, for example, both 80 % understorey cover as well as 80 % overstorey cover. We took the average of the measures from the two plots to give a single measure for each vegetation attribute at each site, and used these in our subsequent analyses.
Data analysis
Gradients in vegetation structure
We quantified vegetation attributes at each site to compare structure across the three vegetation types. We used principal components analysis (PCA) to summarise the major gradients in vegetation structure using a co-variance matrix of percentage cover of the six vegetation attributes. We used the first two components of the PCA as response variables in a linear mixed model to compare the changes in vegetation structure within and among vegetation types in GenStat 14 (VSNI 2013). We fitted fire (burnt, unburnt), vegetation type (forest, woodland, heath), and time since fire (1 year, 6 years) as fixed factors, and site as a random factor to account for repeated measures of each site.
Bird species richness
We used the software EstimateS 9.1 (Colwell 2013) to estimate ‘true’ species richness of birds from our study sites and assess the thoroughness of our bird surveys. We calculated estimates using the abundance-based coverage (ACE) estimator and the Chao 1 estimator, and compared the estimated number of species with the observed number of species. These two estimators consider the number of rare species in a sample (counts from a site) and were the most appropriate for our data (Magurran and McGill 2011). We calculated separate species richness estimates for each vegetation type in 2004 and 2009.
We tested for differences in bird species richness among burnt and unburnt sites in each vegetation type using a generalised linear mixed model with a Poisson error distribution (estimated dispersion = 0.91) and log-link function in GenStat 14 (VSNI 2013). We fitted burnt status, vegetation type, and time since fire as fixed factors. We also fitted site as a random factor to account for temporal autocorrelation due to repeated measures of each site. The significance of effects was determined using Wald tests. We tested for spatial autocorrelation in model residuals using the ‘correlog’ function in the ‘ncf’ package in R (Zuur et al. 2009), but found no evidence of this for species richness among sites in close proximity.
Bird assemblage composition
We examined differences in bird assemblage composition among burnt and unburnt sites in each vegetation type using three different multivariate approaches. For all tests, bird abundance data were square root transformed to reduce the influence of abundant species. We used a Mantel test to examine the correlation between pairwise site dissimilarity in bird assemblage composition (Bray–Curtis), and pairwise site dissimilarity in vegetation structure (Euclidean) across all sites. We repeated the test for each survey year to determine whether the correlation was different at 1 year after the wildfire compared with 6 years after wildfire. Significance was determined using 9,999 permutations of the data in PC-Ord 5 (McCune and Grace 2002).
We used canonical analysis of principal coordinates (Anderson and Willis 2003) to examine how bird assemblage composition responded to fire within each vegetation type. We constrained the ordination with regard to the two gradients in vegetation complexity identified in the PCA above. We plotted mean scores in an ordination diagram, and grouped sites according to their vegetation type, burnt status, and time since fire. This allowed us to compare the interactive effects of vegetation type and fire on changes in bird assemblage composition. We examined the strength of the correlation between the two vegetation structure gradients and the first two constrained ordination axes, and identified bird species that contributed strongly to the multivariate patterns (r ≥ ±0.3). We used 9,999 permutations of the data to test for the significance of correlations between the two vegetation gradients and the first two constrained ordination axes.
We used Permutational Analysis of Multivariate Dispersion (Anderson et al. 2006) to test for differences in among-site variability in assemblages from each vegetation type between 1 and 6 years after the wildfire. This test calculates the average distance to the centroid of a group of samples projected in multivariate space, with a greater distance to centroid indicating greater among-sample variability (Anderson et al. 2006). Statistical significance was determined from 9,999 permutations of the data.
Results
Gradients in vegetation structure
Our principal components analysis produced two new axes that accounted for a combined 73 % of the variation in vegetation structure (Table S2). The first axis (PC1) had high loadings for leaf litter and overstorey cover, representing a gradient of increasing overstorey density, i.e. increasing vertical structure (Fig. S1). The second axis (PC2) had high positive loadings for understorey cover and negative loadings for bare ground, representing a gradient of increasing understorey density, i.e. increasing horizontal structure (Fig. S1). The contrasting vertical and horizontal structure of the three vegetation types can be seen clearly in Fig. 1. We found a significant interactive effect of fire, vegetation type and time on site vertical structure (PC1) scores (Wald2 = 11.61, p = 0.005, Table S3). Vertical structure of unburnt heath vegetation, but not burnt heath vegetation, increased from 1 to 6 years after the wildfire (Fig. S2). For horizontal structure (PC2), we found no interactive effect of fire, vegetation type and time (Wald2 = 1.97, p = 0.380), but there was a significant interaction between fire and time (Wald1 = 51.40 p < 0.001). There was a large decrease in horizontal structure for all vegetation types 1 year after the fire, with the largest being evident for heath vegetation. We found no difference in horizontal structure between burnt and unburnt sites after 6 years (Fig. S2).
Bird species richness
We recorded 4,181 birds from 68 species during our surveys in 2004 and 2009 (Table S4). Our surveys were thorough, ranging from 68 to 97 % in our 2004 surveys, and 88–97 % in our 2009 surveys (Table S5). We detected no interactive effects of fire, vegetation type and year on bird species richness (Wald2 = 1.93, p = 0.387). However, we found a significant main effect of vegetation type (Wald2 = 35.97, p < 0.001, see Table S6), with species richness of birds lowest in heath and highest in forest (Fig. 2). Although no significant interaction was detected, the difference in richness between burnt and unburnt vegetation was greater in woodland and forest than in heath at 1 year after the fire. These apparent differences were absent 6 years later.

Mean (±SE) species richness of bird assemblages in burnt and unburnt sites in Heath, Woodland and Forest vegetation at 1 (2004) and 6 years (2009) after wildfire
Bird assemblage composition
We found a significant positive correlation between pairwise bird assemblage dissimilarity and vegetation structural dissimilarity, and that the magnitude of the correlation increased from 1 year after the fire (r = 0.218, p = 0.0003) to 6 years after the fire (r = 0.304, p < 0.0001). This indicated that sites with similar vegetation structure shared similar bird assemblages, but that this relationship was weaker after recent fire.
The first two axes of the principal coordinate analysis explained 20.1 and 11.5 % of the variance in bird assemblage composition, respectively (Fig. 3). Ordination of constrained site scores showed that bird assemblages were clearly differentiated between vegetation types along a gradient of increasing vertical structure, represented by axis one (Fig. 3a, p < 0.001). Bird assemblages were also differentiated within each vegetation type along a gradient of increasing horizontal structure, represented by axis two (Fig. 3a, p < 0.001). Sites burnt 1 year after the wildfire always had bird assemblages that changed in composition in the direction of decreasing horizontal structure. Further, the magnitude of the difference in bird composition between burnt and unburnt sites was greatest in heath vegetation, followed by woodland, and smallest in forest vegetation.

Ordination of site scores derived from canonical correlation analysis of bird assemblages in three vegetation types at 1 year and 6 years after fire. a Mean (±SE) scores of burnt (B) and unburnt (UB) sites at 1 and 6 years AFTER wildfire. b Correlation biplot of bird species (grey dots) and vegetation structural gradients (Understorey and Overstorey) with canonical axes. Bird species contributing strongly to patterns are indicated by the dark grey dots (see Table S4 for scientific names)
Several bird species were strongly correlated with the canonical ordination axes (Fig. 3b). Two heath specialists, the southern emu-wren (Stipiturus malachurus) and eastern bristlebird (Dasyornis brachypterus), were each strongly correlated with low vertical structure (i.e. heath vegetation), but also were correlated with high horizontal structure (i.e. dense understorey). In contrast, two forest specialists, the grey fantail (Rhipidura albiscapa) and eastern spinebill (Acanthorhynchus tenuirostris), were correlated with high horizontal and vertical structure (i.e. forest with dense understorey). The crimson rosella (Platycercus elegans) was strongly correlated with low horizontal structure and high vertical structure (i.e. forest with open understorey).
We found a significant overall difference in spatial variability of bird assemblages among vegetation types at 1 and 6 years after the wildfire (F = 4.34, p < 0.001). Post-hoc comparisons indicated that variability among sites within each vegetation type was significantly higher 1 year after fire compared with 6 years for heath and woodland, but not forest vegetation (Fig. 4). All vegetation types had similar levels of among-site variability 6 years after fire.

Differences in among-site dissimilarity of bird assemblage across burnt and unburnt sites between Heath, Woodland and Forest at 1 (2004) and 6 years (2009) after wildfire. Pairwise differences are significant at *p < 0.05
Discussion
Heterogeneous landscapes provide distinct challenges to understanding the effects of disturbance on biodiversity (Turner 1987; He and Mladenoff 1999; Fuhlendorf et al. 2006; Lindenmayer et al. 2014). Our study of the effects of fire in a landscape with a mix of different vegetation communities revealed an interacting role of vertical and horizontal vegetation structure in moderating bird assemblage responses to fire. Our results supported our hypothesis that increased vegetation structure would reduce the effects of wildfire on bird assemblages, although we found this only for composition changes and not richness changes. Below we discuss the links between vegetation structure and bird assemblage responses, and then develop our findings into a conceptual model to provide a framework to link vegetation structure with fire disturbance effects on animal assemblages.
The moderating influence of vegetation structure
We have shown how vertical and horizontal structure is associated with bird assemblage patterns among different vegetation types. Effects of vegetation structure on bird communities are commonly reported (e.g. MacArthur and MacArthur 1961; Bohning-Gaese 1997; Davis et al. 2000; Montague-Drake et al. 2009), but the separate effects of vertical and horizontal structure are less well understood. For example, we found higher species richness of birds in forest vegetation compared with woodland or heath vegetation. This was due to greater overstorey cover, which was a strong driver of the occurrence of the grey fantail and eastern spinebill, for example. It is hypothesised that greater overstorey cover generates greater resource availability in the form of foliage, branch and trunk foraging substrates, as well as shelter from predators for a range of species (Recher et al. 1985; Montague-Drake et al. 2009). Although vertical structure was important for determining differences in bird assemblages between vegetation types, horizontal structure in the lower stratum was associated with changes in bird assemblage composition within vegetation types. The variation in horizontal structure was due to changes in understorey shrub cover, and was a strong driver of the occurrence of heath specialists such as the eastern bristlebird and southern emu wren (Lindenmayer et al. 2008a).
Notably, we found a loss of horizontal structure in burnt areas of all vegetation types, and this was matched by a corresponding change in bird assemblage composition along this structural gradient. This suggests that a change in the density of shrubs and foliage in this stratum is a key mechanism linking the effects of fire to changes in bird assemblage composition across multiple vegetation types. This can lead to negative impacts on foraging resources and shelter from potential predators for species that use this vegetation layer (Davis et al. 2000; Brotons et al. 2004; Lindenmayer et al. 2009). Further, the magnitude of this change increased when vertical structure was low or absent. This was highlighted by the greater change in assemblage composition in heath compared with forest vegetation. This pinpoints the critical role of vertical structure in moderating the effects of fire in our study system, even after the loss of understorey vegetation.
Our study provides a novel example of the moderating effects of vegetation structure in a terrestrial ecosystem. Previous research has identified vegetation structure as an important determinant of the recovery of faunal assemblages after fire, including for reptiles (Lindenmayer et al. 2008b) and mammals (Monamy and Fox 2000). However, these examples have focused on the role of vegetation in moderating the recovery trajectories, and not the initial impacts. Our finding that structurally simple vegetation may be more vulnerable to perturbations than complex vegetation with multiple strata, and experience a more profound short-term change in its associated fauna, suggests that habitat structural attributes may play a key role in moderating the short-term impacts of fire.
A conceptual model to explore fire effects on biodiversity in heterogeneous landscapes
Our findings support the hypothesis that increasing vegetation structure moderates the effects of fire on bird assemblages. However, we found that this result was the case only for bird composition changes and not for species richness. This suggests a complex interplay between fire, vegetation structure and bird assemblages, with fire affecting this relationship in different ways for species richness and species composition. From this result, we describe a conceptual model (Fig. 5) that summarises our results into a generalised prediction of the short-term impact of fire on bird diversity across a vegetation gradient of increasing structural complexity. We split bird diversity into its species richness (alpha-diversity) and compositional (beta-diversity) components to highlight the contrasting patterns found in our study. Our model predicts that the short-term change in species richness after fire will be higher in complex vegetation relative to simpler vegetation (compare richness of burnt versus unburnt sites 2004 forest with 2004 heath in Fig. 2). Our model also predicts that fire will cause a greater change in species composition in simple vegetation relative to complex vegetation (compare among-site dissimilarity of sites from 2004 heath with 2004 forest in Fig. 4). The contrasting response of species richness and composition to fire is in part due to the underlying relationship between species richness and overstorey cover (see Fig. 2). Complex vegetation characterised by multiple strata contains a greater variety of feeding resources, foraging substrates, and nesting and perching sites, thus enabling greater specialisation among species and driving higher richness in forest vegetation (Recher 1969; Tews et al. 2004). Moreover, this gradient in richness means that structurally simple vegetation, with lower species richness, is susceptible to proportionally greater changes in bird species composition, even though absolute changes in richness can be smaller. The different effects of fire on alpha and beta components of animal diversity has not been explored in detail (Farnsworth et al. 2014), and represents a potential new area for investigation.

A conceptual model depicting the expected short-term change in bird diversity after fire across a gradient of increasing vegetation structure. Species richness (solid line) can be expected to show the greatest change in structurally complex vegetation and the smallest change in structurally simple vegetation. In contrast, species composition (dotted line) can be expected to show the greatest change in simple vegetation and smallest change in complex vegetation
Our conceptual model has clear links with the recent synthesis of disturbance effects on ecosystems by Peters et al. (2011). These authors describe a framework that outlines three key aspects important to understanding the disturbance cycle, including (i) initial system properties, (ii) disturbance mechanisms and (iii) post-disturbance legacies. Our model addresses the first two of these aspects. Knowledge of the spatial distribution of structurally distinct vegetation types provides valuable information about initial ecosystem properties, and enables better prediction of fire impact on bird communities. Further, the role of vegetation complexity in driving species richness gradients across landscapes provides some information about potential mechanisms of fire effects. Incorporating information about fire severity and frequency in each vegetation type could be an important next step in understanding post-disturbance legacies (Whelan 1995; Franklin et al. 2000; Peters et al. 2011). This would complement other conceptual work on periodic disturbances (Tanentzap et al. 2013; Thom et al. 2013), and lead towards better integration of prediction with knowledge of disturbance regimes.
We have outlined an example of the relative impacts of fire on animal communities across structurally distinct vegetation types within the same landscape. This provides critical context to our findings, with our conceptual model likely to be most applicable to heterogeneous landscapes comprising a mix of distinct vegetation types, such as is typical in many Mediterranean-climate regions around the world (Cowling et al. 1996; Lavorel 1999). However, a key strength of our conceptual model is that it builds on the well-established relationship between animal species diversity and habitat structure and complexity. Many empirical studies have documented the effects of habitat complexity and heterogeneity on various groups of organisms and in different biomes (e.g. Heck and Wetstone 1977; August 1983; Hansen 2000; Lassau and Hochuli 2004; Tews et al. 2004). This rich literature could be used to develop further mechanistic hypotheses about the disturbance response of different taxa in other kinds of ecosystems. For example, the composition and complexity of plant communities has been linked to the diversity and biomass of arthropod communities through resource and habitat provision (Borer et al. 2012). Alternatively, vegetation structure may underpin reptile responses by providing basking opportunities and foraging sites (Pike et al. 2011). Of course, different specific predictions might be tailored for different taxa, as the relative importance of different measures of structure, complexity or heterogeneity will vary.
Implications
The moderating effect of vegetation structure has significant implications for fire management and biodiversity conservation. Altered disturbance regimes can lead to simplification of landscapes and the loss of variation in vegetation structure and composition (Vandvik et al. 2005; Grossmann and Mladenoff 2007). From the perspective of our study, such simplification may also lead to a loss of variation in faunal responses to disturbance among different vegetation types, and may diminish the resilience of heterogeneous landscapes to major ecological disturbances such as fire. In such landscapes, meta-population and meta-community dynamics among vegetation patches are critical to their functioning, and contribute to landscape-scale biodiversity patterns (Atauri and de Lucio 2001; Biswas and Wagner 2012; Tscharntke et al. 2012). Our finding that habitat heterogeneity is linked to the differential responses of birds to fire within a landscape, provides further evidence that local-scale processes underpin the maintenance of biodiversity within landscapes (Brotons et al. 2004; Barton et al. 2013). It also supports theory that landscape heterogeneity can moderate the effects of disturbance more generally (Turner 1987, 2010).
Much of the current thinking about managing the effects of fire on biodiversity is dominated by the patch-mosaic concept where a diversity of fire histories is considered important for generating ecosystem heterogeneity (Parr and Andersen 2006; Kelly et al. 2012). Our finding that vegetation structure can moderate the impacts of fire on bird assemblages goes beyond the domain of the patch-mosaic concept and suggests that fire management needs to incorporate vegetation attributes in addition to fire history attributes. This is because the short-term impacts of fire on fauna will vary depending on vegetation structure, thus driving heterogeneity in faunal assemblages independent of spatial patterns of fire history. This will enable the prediction of the relative impact of fire in heterogeneous landscapes with variable vegetation. Where fire occurs across multiple vegetation types, this requires careful consideration of the potential for more pronounced effects on structurally simple vegetation, and associated changes in faunal assemblages. This perspective is somewhat counter-intuitive, as fire management practices may need to be concerned about both the structurally complex and species-rich places in a landscape, as well as the less complex or less diverse parts of a landscape. Our model provides a predictive framework to investigate in more detail the effects of fire and other disturbances on faunal assemblages, and the moderating role of vegetation structure in heterogeneous landscapes.
References
Anderson MJ, Willis TJ (2003) Canonical analysis of principal coordinates: a useful method of constrained ordination for ecology. Ecology 84(2):511–525
Anderson MJ, Ellingsen KE, McArdle BH (2006) Multivariate dispersion as a measure of beta diversity. Ecol Lett 9(6):683–693
Atauri JA, de Lucio JV (2001) The role of landscape structure in species richness distribution of birds, amphibians, reptiles and lepidopterans in Mediterranean landscapes. Landscape Ecol 16(2):147–159
August PV (1983) The role of habitat complexity and heterogeneity in structuring tropical mammal communities. Ecology 64(6):1495–1507
Barton PS, Cunningham SA, Manning AD, Gibb H, Lindenmayer DB, Didham RK (2013) The spatial scaling of beta diversity. Glob Ecol Biogeogr 22(6):639–647
Biswas SR, Wagner HH (2012) Landscape contrast: a solution to hidden assumptions in the metacommunity concept? Landscape Ecol 27(5):621–631
Bohning-Gaese K (1997) Determinants of avian species richness at different spatial scales. J Biogeogr 24(1):49–60
Borer ET, Seabloom EW, Tilman D (2012) Plant diversity controls arthropod biomass and temporal stability. Ecol Lett 15(12):1457–1464
Bradstock RA, Bedward M, Gill AM, Cohn JS (2005) Which mosaic? A landscape ecological approach for evaluating interactions between fire regimes, habitat and animals. Wildl Res 32(5):409–423
Bradstock RA, Gill AM, Williams RJ (eds) (2012) Flammable Australia: fire regimes, biodiversity and ecosystems in a changing world. CSIRO Publishing, Collingwood
Brotons L, Herrando S, Martin JL (2004) Bird assemblages in forest fragments within Mediterranean mosaics created by wild fires. Landscape Ecol 19(6):663–675
Clavero M, Brotons L, Herrando S (2011) Bird community specialization, bird conservation and disturbance: the role of wildfires. J Anim Ecol 80(1):128–136
Collins SL, Calabrese LB (2012) Effects of fire, grazing and topographic variation on vegetation structure in tallgrass prairie. J Veg Sci 23(3):563–575
Colwell RK (2013) EstimateS 9.1: statistical estimation of species richness and shared species from samples. http://purl.oclc.org/estimates
Connell JH (1978) Diversity in tropical rain forests and coral reefs—high diversity of trees and corals is maintained only in a non-equilibrium state. Science 199(4335):1302–1310
Coppedge BR, Fuhlendorf SD, Harrell WC, Engle DM (2008) Avian community response to vegetation and structural features in grasslands managed with fire and grazing. Biol Conserv 141(5):1196–1203
Cowling RM, Rundel PW, Lamont BB, Arroyo MK, Arianoutsou M (1996) Plant diversity in mediterranean-climate regions. Trends Ecol Evol 11:326–362
Cunningham RB, Lindenmayer DB, Nix HA, Lindenmayer BD (1999) Quantifying observer heterogeneity in bird counts. Austral Ecol 24:270–277
Davis MA, Peterson DW, Reich PB et al (2000) Restoring savanna using fire: impact on the breeding bird community. Restor Ecol 8(1):30–40
de Lima RF, Dallimer M, Atkinson PW, Barlow J (2013) Biodiversity and land-use change: understanding the complex responses of an endemic-rich bird assemblage. Divers Distrib 19(4):411–422
Driscoll DA, Lindenmayer DB (2010) Assembly rules are rare in SE Australian bird communities, but sometimes apply in fragmented agricultural landscapes. Ecography 33(5):854–865
Faivre N, Roche P, Boer MM, McCaw L, Grierson PF (2011) Characterization of landscape pyrodiversity in Mediterranean environments: contrasts and similarities between south-western Australia and south-eastern France. Landscape Ecol 26(4):557–571
Farnsworth LM, Nimmo DG, Kelly LT, Clarke MF, Bennett AF (2014) Does pyrodiversity beget alpha, beta or gamma diversity? A case study using reptiles from semi-arid Australia. Divers Distrib. doi:10.1111/j.1466-8238.2011.00747.x
Forman RTT (1995) Land mosaics: the ecology of landscapes and regions. Cambridge University Press, Cambridge
Fox JW (2013) The intermediate disturbance hypothesis should be abandoned. Trends Ecol Evol 28(2):86–92
Franklin JF, Lindenmayer DB, MacMahon JA et al (2000) Threads of continuity: ecosystem disturbances, biological legacies and ecosystem recovery. Conserv Biol Pract 1:8–16
Franklin JF, Spies TA, Van Pelt R et al (2002) Disturbances and structural development of natural forest ecosystems with silvicultural implications, using Douglas-fir forests as an example. For Ecol Manage 155(1–3):399–423
Fuhlendorf SD, Harrell WC, Engle DM, Hamilton RG, Davis CA, Leslie DM (2006) Should heterogeneity be the basis for conservation? Grassland bird response to fire and grazing. Ecol Appl 16(5):1706–1716
Grossmann EB, Mladenoff DJ (2007) Open woodland and savanna decline in a mixed-disturbance landscape (1938–1998) in the Northwest Wisconsin (USA) Sand Plain. Landscape Ecol 22:43–55
Hansen RA (2000) Effects of habitat complexity and composition on a diverse litter microarthropod assemblage. Ecology 81(4):1120–1132
He HS, Mladenoff DJ (1999) Spatially explicit and stochastic simulation of forest-landscape fire disturbance and succession. Ecology 80(1):81–99
Heck KL, Wetstone GS (1977) Habitat complexity and invertebrate species richness and abundance in tropical seagrass meadows. J Biogeogr 4(2):135–142
Ikin K, Beaty RM, Lindenmayer DB, Knight E, Fischer J, Manning AD (2013) Pocket parks in a compact city: how do birds respond to increasing residential density? Landscape Ecol 28(1):45–56
Keeley JE, Fotheringham CJ, Baer-Keeley M (2006) Demographic patterns of postfire regeneration in mediterranean climate shrublands of California. Ecol Monogr 76(2):235–255
Kelly LT, Nimmo DG, Spence-Bailey LM et al (2011) Influence of fire history on small mammal distributions: insights from a 100-year post-fire chronosequence. Divers Distrib 17(3):462–473
Kelly LT, Nimmo DG, Spence-Bailey LM et al (2012) Managing fire mosaics for small mammal conservation: a landscape perspective. J Appl Ecol 49(2):412–421
Kerby JD, Fuhlendorf SD, Engle DM (2007) Landscape heterogeneity and fire behavior: scale-dependent feedback between fire and grazing processes. Landscape Ecol 22(4):507–516
Kutt AS, Martin TG (2010) Bird foraging height predicts bird species response to woody vegetation change. Biodivers Conserv 19(8):2247–2262
Lassau SA, Hochuli DF (2004) Effects of habitat complexity on ant assemblages. Ecography 27(2):157–164
Lavorel S (1999) Ecological diversity and resilience of Mediterranean vegetation to disturbance. Divers Distrib 5:3–13
Lindenmayer DB, Wood JT, Cunningham RB et al (2008a) Testing hypotheses associated with bird responses to wildfire. Ecol Appl 18(8):1967–1983
Lindenmayer DB, Wood JT, MacGregor C et al (2008b) How predictable are reptile responses to wildfire? Oikos 117(7):1086–1097
Lindenmayer DB, MacGregor C, Wood JT et al (2009) What factors influence rapid post-fire site re-occupancy? A case study of the endangered Eastern Bristlebird in eastern Australia. Int J Wildland Fire 18(1):84–95
Lindenmayer DB, Blanchard W, McBurney L et al (2014) Complex responses of birds to landscape-level fire extent, fire severity and environmental drivers. Divers Distrib. doi:10.1111/ddi.12172
MacArthur R, MacArthur JW (1961) On bird species-diversity. Ecology 42(3):594–598
Magurran AE, McGill BJ (eds) (2011) Biological diversity: frontiers in measurement and assessment. Oxford University Press Inc., New York
McCune B, Grace JB (2002) Analysis of ecological communities. MjM Software Design, Gleneden Beach
Monamy V, Fox BJ (2000) Small mammal succession is determined by vegetation density rather than time elapsed since disturbance. Austral Ecol 25(6):580–587
Montague-Drake RM, Lindenmayer DB, Cunningham RB (2009) Factors affecting site occupancy by woodland bird species of conservation concern. Biol Conserv 142(12):2896–2903
Murphy BP, Bowman D (2012) What controls the distribution of tropical forest and savanna? Ecol Lett 15(7):748–758
Parr CL, Andersen AN (2006) Patch mosaic burning for biodiversity conservation: a critique of the pyrodiversity paradigm. Conserv Biol 20(6):1610–1619
Pastro LA, Dickman CR, Letnic M (2011) Burning for biodiversity or burning biodiversity? Prescribed burn versus wildfire impacts on plants, lizards, and mammals. Ecol Appl 21(8):3238–3253
Peters DPC, Lugo AE, Chapin FS et al (2011) Cross-system comparisons elucidate disturbance complexities and generalities. Ecosphere 2(7):1–26
Pickett STA, White PS (eds) (1985) The ecology of natural disturbance and patch dynamics. Academic Press, New York
Pike DA, Webb JK, Shine R (2011) Removing forest canopy cover restores a reptile assemblage. Ecol Appl 21(1):274–280
Recher HF (1969) Bird species diversity and habitat diversity in Australia and North America. Am Nat 103(929):75–80
Recher HF, Holmes RT, Schulz M, Shields J, Kavanagh R (1985) Foraging patterns of breeding birds in eucalypt forest and woodland of southeastern Australia. Austral Ecol 10:399–419
Santana J, Porto M, Gordinho L, Reino L, Beja P (2012) Long-term responses of Mediterranean birds to forest fuel management. J Appl Ecol 49(3):632–643
Schimmel J, Granstrom A (1996) Fire severity and vegetation response in the boreal Swedish forest. Ecology 77(5):1436–1450
Smith AL, Bull CM, Driscoll DA (2012) Post-fire succession affects abundance and survival but not detectability in a knob-tailed gecko. Biol Conserv 145(1):139–147
Tanentzap AJ, Lee WG, Schulz KAC (2013) Niches drive peaked and positive relationships between diversity and disturbance in natural ecosystems. Ecosphere 4(11):133
Tews J, Brose U, Grimm V et al (2004) Animal species diversity driven by habitat heterogeneity/diversity: the importance of keystone structures. J Biogeogr 31(1):79–92
Thom D, Seidl R, Steyrer G, Krehan H, Formayer H (2013) Slow and fast drivers of the natural disturbance regime in Central European forest ecosystems. For Ecol Manage 307:293–302
Tscharntke T, Tylianakis JM, Rand TA et al (2012) Landscape moderation of biodiversity patterns and processes—eight hypotheses. Biol Rev 87(3):661–685
Turner MG (ed) (1987) Landscape heterogeneity and disturbance. Springer-Verlag, New York
Turner MG (2010) Disturbance and landscape dynamics in a changing world. Ecology 91(10):2833–2849
Tyre AJ, Tenhumberg B, Field SA, Niejalke D, Parris K, Possingham HP (2003) Improving precision and reducing bias in biological surveys: estimating false-negative error rates. Ecol Appl 13(6):1790–1801
Vandvik V, Heegaard E, Maren IE, Aarrestad PA (2005) Managing heterogeneity: the importance of grazing and environmental variation on post-fire succession in heathlands. J Appl Ecol 42(1):139–149
VSNI (2013) GenStat for windows, 14th edn. VSNI Ltd., Hemel Hempstead
Watson SJ, Taylor RS, Nimmo DG et al (2012) Effects of time since fire on birds: how informative are generalized fire response curves for conservation management? Ecol Appl 22(2):685–696
Westgate MJ, Driscoll DA, Lindenmayer DB (2012) Can the intermediate disturbance hypothesis and information on species traits predict anuran responses to fire? Oikos 121(10):1516–1524
Whelan RJ (1995) The ecology of fire. Cambridge University Press, Cambridge
Zuur AF, Ieno EN, Walker NJ, Saveliev AA, Smith GM (2009) Mixed effects models and extensions in ecology with R. Springer, New York
Acknowledgments
We thank staff from Parks Australia and the Wreck Bay Aboriginal Community for their strong support of our work. We thank Claire Foster for comments on an early draft of the manuscript. Funding for this study came from Parks Australia, the Department of Defence, and the Australian Research Council. All work reported in this study complies with the current laws of Australia.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Barton, P.S., Ikin, K., Smith, A.L. et al. Vegetation structure moderates the effect of fire on bird assemblages in a heterogeneous landscape. Landscape Ecol 29, 703–714 (2014). https://doi.org/10.1007/s10980-014-0017-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-014-0017-z