
Abstract
The desire to improve urban sustainability is motivating many city planners to adopt growth strategies that increase residential density, leading to substantial changes to urban landscapes. What effect this change will have on biodiversity remains unclear, but it is expected that the role of public greenspace in providing wildlife habitat will become critical. We explored the role of urban “pocket parks” as habitat for birds, and how this role changed with increasing residential density in the surrounding neighbourhood. We found that parks in neighbourhoods with high levels of public greenspace (corresponding to less residential land) supported more bird species and individuals overall, and more woodland-dependent species, insectivores and hollow-nesters. Total greenspace area was more important (included in the best ranked models for all bird responses) than the configuration (number, average size and connectivity) of greenspace patches. The majority of species were common suburban birds, indicating that species we assume are tolerant to urban areas will be negatively affected by increasing residential density. Parks form part of an interconnected network of urban open space. For parks to continue to support a diverse native bird community, the network must be viewed, managed, and maintained in its entirety. We suggest three key management actions to improve the bird diversity values of urban greenspaces in compact cities: (1) Increase urban greenspace cover in residential neighbourhoods. (2) Increase vegetation structure in greenspace. (3) Encourage homeowners to plant trees and shrubs.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Every year the number of people living in urban areas grows by approximately 1.96 %, compared with 0.11 % in rural areas (United Nations 2011). This worldwide rapid urban growth has major impacts on the social, economic and environmental sustainability of cities (Andersson 2006). The desire to improve urban sustainability is motivating many city planners to adopt growth strategies that increase residential density, termed ‘residential densification’ (Pauleit et al. 2005; Tratalos et al. 2007; Musacchio 2009). This approach meets many urban sustainability goals, including reduced urban sprawl, a smaller land-take for each person, reduced transport costs, and improved water and energy efficiency (Tratalos et al. 2007). Not surprisingly, however, residential densification leads to substantial changes in the urban form (the physical design and layout of the city). Cities once characterised by large residential blocks and extensive greenspaces (for example, gardens, road verges, laneways and parks) are moving towards “compact city” plans of small residential blocks and high levels of building, road, and pavement cover. This residential densification affects established urban areas, through the sub-division of blocks and infill of greenspace, as well as new urban areas through the deliberate planning of high density housing developments (Pauleit et al. 2005).
Although the concept of sustainability incorporates environmental considerations (Andersson 2006), it is unclear what effect residential densification and associated changes to the urban form will have on urban biodiversity (Tratalos et al. 2007). It is expected that as the total amount of greenspace decreases, the role of remaining greenspace, particularly parks, in providing wildlife habitat will become critical. However, the capacity of parks to support wildlife has been found to be strongly affected by local landscape context. Lim and Sodhi (2004), for example, found that the amount of built-up environment within 250 m of parks in Singapore had significant negative effects on the richness and abundance of several bird guilds, including insectivores and tree-cavity nesters. The isolation of a park from other greenspaces (Oliver et al. 2011), and the age (Loss et al. 2009) and degree of urban intensity surrounding a park (Ortega-Alvarez and MacGregor-Fors 2009; Suarez-Rubio and Thomlinson 2009; Huste and Boulinier 2011), are also expected to have negative effects. Residential densification, therefore, may simultaneously cause parks to become refugia or habitat “islets” (Rey Benayas et al. 2008) within the urban matrix, whilst lessening their capacity to support functional wildlife populations. It is very important, therefore, to assess how different urban forms in the neighbourhoods surrounding parks affect wildlife within them.
Landscape ecology, with its focus on the spatial patterns caused by ecological processes (Wiens 2009), is an effective framework from which to understand differences between alternative urban forms. This approach characterises landscapes by two spatial components: composition and configuration (Fahrig et al. 2011). Composition refers to the amount and diversity of different land use (e.g. residential, greenspace) or land cover (e.g. buildings, trees) patches in the landscape (Botequilha Leito et al. 2006). In comparison, configuration refers to how these patches are spatially distributed or arranged (e.g. average residential block size, connectivity between greenspaces). These two aspects of landscape structure can have independent or interactive effects on the ecological processes within urban parks (Botequilha Leito et al. 2006). The relationship between landscape composition and configuration also can be viewed from the perspective of habitat loss and habitat fragmentation, and there is growing interest regarding their relative importance to biodiversity (e.g. Fahrig 2003; Lindenmayer and Fischer 2006; Laurance 2008; Thornton et al. 2011). To fully understand how changes to the urban form affect the capacity of parks to support wildlife, therefore, it is vital to disentangle the different effects of land use and land cover composition and configuration.
The aim of this study was to understand the role of “pocket parks” as habitat for birds, and how this role changed with different urban forms in the surrounding neighbourhood. Pocket parks were the smallest type of parks (≤2 ha) within our study area in Canberra, southeast Australia (ACT Government 2006), but their high representation in the urban landscape (within 0.4 km of every residential dwelling) make these areas important. Residential densification is expected to change the composition of urban land use and land cover, by increasing residential land use and decreasing public greenspace, and public and private tree cover within the urban landscape (Pauleit et al. 2005). It is also expected to change the configuration of urban land use by increasing residential block numbers and decreasing residential block sizes, greenspace patch sizes and numbers, and greenspace connectivity. We investigated the total species richness and abundance of all birds in the pocket parks, as well as the richness of woodland dependent species, insectivorous species, and tree-cavity/hollow-nesting species. We predicted that residential densification would have negative effects on urban bird diversity, so that pocket parks in neighbourhoods with an urban form characterised by high residential density would support fewer bird species and individuals overall, and fewer woodland, insectivorous and hollow-nesting species.
Methods
Study area and experimental design
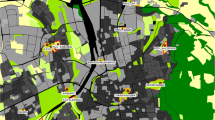
Our study was located in southeastern Australia, in Canberra, Australian Capital Territory (ACT). Canberra is the capital city of Australia, and with a population of 362 000 people and covering 814 km2, it is the country’s largest inland city (ABS 2010). Urban design standards require that neighbourhood “pocket parks” are located within 400 m of every dwelling (ACT Government 2006). These are small (0.25–2 ha) residential parks that include playground facilities and are typically used for recreation. For this study, we identified pocket parks that were between 0.5 and 2 ha, >500 m from other pocket parks, >250 m from protected nature reserves, contained eucalypt (Eucalyptus spp.) trees, and located in suburbs where the median residential block size was between 200 and 1100 m2. We located the geographic centroid of each pocket park and placed a 50 m radius (0.8 ha) site at this point. For pocket parks that were between 0.5 ha and 0.8 ha, or were of an irregular shape, we located the site so that the majority of the site area was within the park boundary. However, we excluded sites that had <60 % of site area within the park boundary. From an original total of 337 pocket parks, this selection process yielded sites in 109 parks in 64 suburbs (Fig. 1a).

Study area in Canberra, southeast Australia. a Study sites in 109 parks (black points) in 64 suburbs (grey polygons). b Depiction of 250 m neighbourhood buffer (white circle) around the centre of the park (white point within white polygon). c Land use composition and configuration categories. d Land cover composition within the neighbourhood buffer
Park and neighbourhood data
Park composition
In our study, we were primarily interested in the effects of local landscape context on park bird diversity. However, several studies have demonstrated the importance of vegetation composition for birds, including woody plant cover (e.g. Shwartz et al. 2008; Fontana et al. 2011) and large tree density (e.g. Stagoll et al. 2012). Thus, to quantify park composition, we measured the tree cover within the park boundary of each site. We recorded the total number of trees per ha (of all species) and the total number of large eucalypts per ha. We defined a large eucalypt as having a diameter at breast height (DBH; 1.3 m above ground level) ≥50 cm (or the equivalent for multi-stemmed trees; following Fischer et al. 2009). This diameter is the minimum trunk measurement for trees to be protected within our study area (Tree Protection Act 2005).
Land use composition and configuration
First, to classify land use composition in the local landscape surrounding our pocket parks, we used ArcMap 9.3 (ESRI, Redlands, CA, USA) to create two spatial layers of public greenspace and private residential land. The greenspace layer combined ACT land designations “urban open space” (i.e. parks, road verges and laneways) and “reserved land” (Territory and Municipal Services, ACT Government, 2010). The residential land layer represented ACT cadastral blocks (ACT Planning and Land Authority, 2008) clipped to the 2006 Canberra residential area, excluding public greenspace. These spatial layers represented the most recent spatial data available. We created 250 m buffers (representing the surrounding “neighbourhood”) around the centre of each site (following Lim and Sodhi 2004; Fig. 1b). This buffer distance reflects maximum gap-crossing abilities of passerine birds in forested and agricultural landscapes in Australia (Robertson and Radford 2009) and urban landscapes in Canada (Tremblay and St. Clair 2011). Within each neighbourhood, we calculated the percent area of greenspace and residential land use in each pocket park neighbourhood. Second, we classified the configuration of greenspace and residential land within each neighbourhood (Fig. 1c). We calculated the number and average size of greenspace patches and residential blocks, and the proximity of each park to the nearest greenspace patch larger than 20 ha. These metrics are readily interpretable and reflect predictions of how increasing residential density will affect residential and greenspace land use configuration.
Land cover composition
To categorize land cover composition within the pocket park neighbourhoods, we developed a land cover classification using Object Based Image Analysis (OBIA) of high resolution satellite imagery (Quickbird imagery pansharpened to 60 cm resolution) and airborne laser scanning data collected in 2005 and 2004, respectively. This technique overcomes several problems associated with urban pixel-based remote sensing approaches, including errors caused by shadows and high spatial heterogeneity (Myint et al. 2011). There are two basic steps involved in OBIA: image segmentation and classification. First, we segmented groups of pixels into image objects, which are relatively homogenous areas that represent real world entities of interest, such as houses or trees. Second, we classified image objects using discrimination rules that utilize image object attributes, their relationships to surrounding objects, and/or their place within image-object hierarchies. We used the Definiens Developer 7 software to implement this approach (Definiens 2008). Using our classification, we categorised land cover within each pocket park neighbourhood into trees and shrubs, herbaceous vegetation and grasses, bare ground, paved surfaces, buildings, and water (Fig. 1d). We assessed the accuracy of the classification by comparing the land cover classification with actual land cover evident on independent high resolution aerial photography at 250 randomly generated points. The classification had an overall accuracy of 93.9 % with a kappa statistic of 0.92, which indicates almost perfect agreement between the actual and classified land cover. We then quantified the percent tree and shrub cover within each park neighbourhood to represent land cover composition. Tree and shrub cover was negatively correlated with herbaceous vegetation and grass (r s = −0.74), bare ground (r s = −0.71), paved surfaces (r s = −0.72), and building cover (r s = −0.48).
Bird response data
We conducted bird surveys during spring (October–November) 2010. In southeastern Australia, this period is the best time to ensure that birds, including summer migrants, are associated with sites (and not just passing through), because birds establishing breeding territories exhibit strong site fidelity (Montague-Drake et al. 2009). One observer (K.I.) surveyed each site twice for 10 min, using 50 m radius point counts (Sutherland et al. 2004). All birds seen or heard within the site were counted, but we excluded birds flying overhead and those within the 50 m radius but outside the park boundary. We surveyed during the first 4 h after dawn (generally between 5:45 AM and 9:45 AM), and avoided rainy or windy days. To minimise bias and ensure the maximum detection of birds, we randomised the order that sites were visited each morning, repeat surveys were undertaken on a separate day, and the order of these repeat surveys was also randomised. Due to the open vegetation and high visibility within our sites, we felt confident that our methods allowed us to detect most bird species present at parks.
We calculated five bird response metrics: (1) total species richness of each park (totalled over the two surveys); (2) abundance of all species (averaged over the two surveys); (3) woodland species richness (following the Birds Australia guild classification for woodland birds developed by Silcocks et al. 2005); (4) insectivore richness (including species that forage on the ground, in shrubs and trees, and from the air); and (5) hollow-nester richness (species that nest in tree cavities—in Australia, all species within this group are secondary-cavity nesters). Our woodland species, insectivore and hollow-nester groups were not mutually exclusive, and individual species could be assigned to more than one group.
Data analysis
Model preparation
We first log-transformed several variables to normalise positively skewed distributions (number of large eucalypts per ha, percent greenspace land use, greenspace patch size and number), and then standardised all variables by subtracting the mean and dividing by the standard deviation to obtain standardised regression coefficients. We checked for possible pairwise correlations (see Table S1 in the Supporting Materials), and found that only percent greenspace and residential land uses were highly correlated (r s = −0.82). For this reason, we decided to not include percent area of residential land in further analysis. We also checked for possible correlations with the age of the neighbourhood derived from gazettal date. We found that percent tree and shrub cover was positively correlated with park age, so we did not include park age in further analysis. We then grouped the explanatory variables by their park or neighbourhood category: park composition, land use composition, land use configuration, and land cover composition (Table 1a), following Rhodes et al. (2009).
Model selection
We followed an information-theoretic approach to model selection (Burnham and Anderson 2002). The aim of this approach was to compare multiple alternative hypotheses (models) to find the one that best explained patterns in the data (in contrast to testing a null hypothesis against a single alternative hypothesis). We constructed a candidate set of 18 models (Table 1b). These models included different combinations of the park and landscape variable groups, but were underlined by two assumptions. First, we assumed that park composition would always be important (given that previous studies have consistently found local vegetation composition to be important for birds (e.g. Shwartz et al. 2008; Fontana et al. 2011; Stagoll et al. 2012). We thus included this variable group (i.e. the two variables “# trees/ha” and “# large eucalypts/ha”) in all 18 models. Second, we assumed that land use and land cover would be equally important, and thus explored all combinations of these variable groups in the models. Two models in the candidate set also included interactions among land use configuration and land cover (to investigate if the effect of greenspace patch size or residential block size differed with different levels of tree and shrub cover).
We fit generalised linear models (McCullagh and Nelder 1989) for the five bird metrics (total species richness, abundance, woodland species richness, insectivore richness and hollow-nester richness). For the richness models, we used a log-link function and a Poisson error distribution. For the abundance models, we log-transformed abundance and used an identity link function with a Gaussian error distribution.
We first fitted the full model containing all of the explanatory variables and interactions. After the analysis, we looked at residuals plots to confirm that model assumptions were met, and removed outliers from subsequent analyses. We confirmed there was no spatial autocorrelation by using correlograms to assess the level of correlation between sites at a range of distances (Bjornstad and Falck 2001; Rhodes et al. 2009). We checked both the raw data and the Pearson residuals of the model containing all of the explanatory variables for each bird metric. In all instances, there was no spatial auto-correlation, indicating that sites were independent of each other.
We used the R package “AICmodavg” to rank the 18 models for each bird metric, based on each model’s Akaike’s Information Criterion corrected for small-sample bias (AICc), following Burnham and Anderson (2002). Models with the lowest AICc, and thus highest Akaike weight (interpreted as the relative likelihood of the model being the best), were considered to have the best fit with the data. Models that had an AICc difference (∆i) ≤ 2 from the best model were considered to be “best-ranked”. For each bird metric, we calculated the 95 % confidence set, which includes the models that have a summed Akaike weight (in order, from highest till lowest) of ≥0.95. We interpreted the Akaike weight of each model as the approximate probability that the model is actually the best, given the data and the set of candidate models. Lastly, we assessed the nature of the responses, i.e. the direction and magnitude of effect sizes, and calculated percent deviance explained (residual deviance/null deviance) for the best-ranking models.
Results
We recorded 44 species of birds, with a mean total species richness of 7.8 (±2.6 standard deviation) and a mean abundance of 11.5 (±6.2) individual birds per site (see Table S2 in the Supporting Materials for our species list). Woodland species comprised 57 % of all species (3.5 ± 1.5 species per site), insectivores comprised 23 % of species (3.8 ± 1.8) and hollow-nesters comprised 34 % of species (2.7 ± 1.7).
Responses to park and neighbourhood composition and configuration
The best-ranked models for all bird metrics included neighbourhood variables, and explained between 77 and 88 % of deviance (Table 2). Greenspace land use composition was included in the best-ranked models (AIC c difference ≤2) for abundance, and for total species, woodland species, insectivore, and hollow-nester richness. Land cover composition was included in the best-ranked models for abundance and insectivore richness. Residential land use configuration was included in the best-ranked models for total species and woodland species richness. Park composition on its own was only included in the best-ranked models for hollow-nester richness. There was some support for greenspace configuration (included in the 95 % confidence sets for all bird metrics, see Table S3 in the Supporting Materials), but there was no support for models that included interactions between land use and land cover.
Nature of responses
We found that all bird metrics were positively related to percent greenspace land use (Table 2, Fig. 2). Greenspace was negatively correlated with residential land use, indicating that the bird metrics would also be negatively correlated. Woodland species richness was positively related to tree and shrub land cover. Total species and woodland species richness was negatively related to the number of neighbourhood residential blocks, and species richness was negatively related to average residential block size. Total species and insectivore richness was negatively related to the number of park trees, but abundance and woodland species and hollow-nester richness was positively related to the number of large trees.

Predicted relationships between percent greenspace land use and a total species richness, b abundance, c woodland species richness, d insectivore richness, and e hollow-nester richness. Predictions are taken from the best-fitting (lowest AIC c ) model for each response variable; model coefficients are given in Table 2. Dashed and solid lines represent minimum (0.0) and maximum (28.9) numbers of large eucalypts per ha, respectively. Grey shadings represent 95 % confidence intervals
Discussion
As cities around the world grow in population (United Nations 2009), urban planners are adopting residential densification strategies to improve urban sustainability (Pauleit et al. 2005; Tratalos et al. 2007; Musacchio 2009). Our study used landscape ecology principles to explore what effects this may have on urban bird diversity, specifically bird richness and abundance in urban “pocket parks”. We found that the capacity of these small greenspaces to support birds, including woodland, insectivorous and hollow- (tree-cavity) nesting species, was affected by the composition and configuration of the urban form in the surrounding neighbourhood. In general, as residential intensity increased, pocket parks had fewer bird species and individuals, supporting our prediction that residential densification may have negative effects on urban bird diversity.
Bird responses to the composition and configuration of the urban form
Parks in neighbourhoods with a high amount of greenspace (and therefore less residential land) supported more bird species and individuals overall, and more woodland species, insectivores and hollow-nesters. This pattern indicates that parks form part of the greenspace network, but are not distinct points of refuge that can offset increasing residential densification. Based on inclusion in the best-ranked models, the configuration (number, average size and connectivity) of greenspace patches was much less important than land use composition (percent greenspace cover). Greenspace configuration reflects habitat fragmentation, and the negative effects of this process on biodiversity have been well-established (e.g. Fahrig 2003; Lindenmayer and Fischer 2006; Laurance 2008; Thornton et al. 2011). However, given the small scale of each neighbourhood and the high mobility of birds, individual species may not perceive small greenspace patches as fragmented, and may be able to move among different greenspaces (Hostetler and Holling 2000). Our findings thus support the growing body of evidence that habitat loss is the prevailing factor driving species declines in human-disturbed landscapes (Fahrig 2003).
The effect of neighbourhood greenspace cover did not change with the proportion of tree and shrub cover in each greenspace patch. This indicates that land use (greenspace vs. residential) may be more important than land cover (treed and shrubby vs. other). However, woodland species richness was higher in parks with neighbourhoods with high tree and shrub cover, demonstrating that neighbourhood vegetation is important for many birds. Residential densification is predicted to decrease vegetation structural complexity, and thus the availability of habitat resources will also decrease. This decline will be to the detriment of woodland species, many of which have shown decreasing population trends over the past 10 years (Bounds et al. 2010), including the nationally threatened Superb Parrot (Polytelis swainsonii) (Environment Protection and Biodiversity Conservation Act 1999; see Table S2 in the Supporting Materials).
Parks in neighbourhoods with high numbers of residential blocks supported fewer bird species overall and fewer woodland species. This finding supports our prediction that the capacity of urban parks to provide habitat for urban wildlife will diminish with increasing residential density. Parks had lower total species richness, however, when average block size in the neighbourhood was larger. This finding was unexpected, given that residential densification leads to smaller block sizes, but may be explained by the large building-to-block ratio of many residential areas. Large blocks may also have large houses, driveways and paved areas, and thus have low habitat resources for birds. Additionally, our spatial layer of cadastral blocks may not have distinguished multi-residential from single-residential blocks, which may have confounded the results and masked some of the negative effects of residential densification due to small block sizes.
How will the bird community change with residential densification?
We found that overall bird richness and abundance, and woodland, insectivore and hollow-nester richness had consistently negative responses to increasing residential density. Thus, changes to the urban form due to residential densification are likely to have similar, detrimental effects on the entire bird community. However, the majority of species we recorded were common suburban birds (Fennell 2009), indicating that sensitive urban-avoiding species (Blair 1996) have already been lost from suburban areas. Greater urban densification would therefore affect currently common species, suggesting that even species that we assume are urban-adapted or tolerant will be negatively affected in the future. This predicted decline of common species is of serious concern, and mirrors recent awareness in Europe that previously abundant farmland species, e.g. the Common Starling (Sturnus vulgaris), are declining due to changes in agricultural practices (Donald et al. 2006). The possibility of similar declines in Australian agricultural areas is also beginning to be recognised (Hanspach et al. 2011; but see Lindenmayer and Cunningham 2011). Less attention has been placed on the declines of common species in urban areas, both in Australia and internationally. We suggest that this is an important conservation issue, and should be a future research priority in urban areas. Longitudinal data, in particular, are especially needed (Lindenmayer and Cunningham 2011).
What will be the role of urban parks in the future?
In neighbourhoods with urban forms characterised by high residential density, pocket parks supported fewer bird species and individuals. It is unlikely, therefore, that these parks will become refugia or habitat “islets” (Rey Benayas et al. 2008) with increasing residential densification. These parks will not support the same number of birds that they currently support, although they may increasingly support relatively more birds compared with residential areas. However, parks form part of an interconnected greenspace network. For parks to continue to support a diverse native bird community, this network must be viewed, managed and maintained in its entirety. Our findings suggest that increasing the total amount of greenspace area (regardless of its configuration) is the single most important planning and management action that can be undertaken to minimise the negative effects of residential densification on urban bird diversity in small urban parks.
Improved urban sustainability is not synonymous with improved biodiversity value, and it will become increasingly important to recognise this if the current trend of residential densification continues. However, the concept of sustainability does encompass environmental considerations (Andersson 2006), and residential densification can lead to many positive environmental outcomes. These include improved water and energy efficiency, leading to lowered greenhouse gas emissions, and reduced urban sprawl, which minimises vegetation clearing for development (Tratalos et al. 2007). The challenge for conservation scientists and managers, therefore, becomes not one of reducing residential densification but one of balancing the trade-offs between different environmental priorities.
Management recommendations
Based on the findings from our study, we suggest three key management actions to improve the bird diversity values of urban greenspaces that are not in conflict with residential densification and compact cities:
-
1.
Increase greenspace cover in residential neighbourhoods. These greenspaces can range in size and management intensity from pedestrian parkland to laneways, informal-use sporting fields, managed town parks, road verges or squares. Over small scales (i.e. <0.2 km2), connectivity and configuration are less important than accumulative greenspace area. Moreover, not only will increasing greenspace cover improve bird diversity values within individual greenspace patches, but recent research demonstrates that greenspaces also provide substantial values to the humans that use them. For example, Luck et al. (2011) found that the proportion of neighbourhood vegetation cover was positively correlated with the personal wellbeing of residents in 36 Australian neighbourhoods.
-
2.
Increase vegetation structure in greenspaces. Parks with high numbers of large (DBH ≥ 50 cm) eucalypts supported more individual birds and more woodland and hollow-nesting species. Recent findings show that as the size of these eucalypts increase, their positive effect on bird diversity also increases (Stagoll et al. 2012). Furthermore, several studies demonstrate that maintaining and improving vegetation structure within urban parks can achieve tangible biodiversity outcomes (Shwartz et al. 2008; Fontana et al. 2011).
-
3.
Encourage homeowners to plant trees and shrubs. Increases in tree cover within the neighbourhood will strengthen the capacity of parks to support woodland-dependent species, of which many in our study area are regionally or nationally declining (see Table S2 in the Supporting Materials). Goddard et al. (2010) suggest that the best results are achieved when homeowners work co-operatively with each other, but also when planners structure new developments so that gardens are clustered together. In this way, the individual constraints imposed by small residential blocks can begin to be overcome.
References
ABS 2010. Australian Demographic Statistics, 3101.0, Dec 2010. Australian Bureau of Statistics
Andersson E (2006) Urban landscapes and sustainable cities. Ecol Soc 11:34
Bjornstad ON, Falck W (2001) Nonparametric spatial covariance functions: estimation and testing. Environ Ecol Stat 8:53–70
Blair RB (1996) Land use and avian species diversity along an urban gradient. Ecol Appl 6:506–519
Botequilha Leito A, Miller J, Ahern J, McGarigal K (2006) Measuring landscapes: a planner’s handbook. Island Press, Washington
Bounds J, Taws N, Cunningham RB (2010) A statistical analysis of trends in occupancy rates of woodland birds in the ACT, December 1998 to December 2008: the ten-year data analysis. Canberra Ornithologists Group
Burnham KP, Anderson DR (2002) Model selection and multimodel inference. Springer, New York
Definiens 2008. Definiens Developer 7.0, User Guide
Donald PF, Sanderson FJ, Burfield IJ, van Bommel FPJ (2006) Further evidence of continent-wide impacts of agricultural intensification on European farmland birds, 1990–2000. Agric Ecosyst Environ 116:189–196
1999. Environment Protection and Biodiversity Conservation Act. Cth. 2005. Tree Protection Act. ACT
Fahrig L (2003) Effects of habitat fragmentation on biodiversity. Annu Rev Ecol Syst 34:487–515
Fahrig L, Baudry J, Brotons L, Burel FG, Crist TO, Fuller RJ, Sirami C, Siriwardena GM, Martin J-L (2011) Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecol Lett 14:101–112
Fennell P (ed) (2009) Birds of Canberra gardens. Canberra Ornithologists Group, Canberra
Fischer J, Stott J, Zerger A, Warren G, Sherren K, Forrester RI (2009) Reversing a tree regeneration crisis in an endangered ecoregion. Proc Natl Acad Sci USA Biol Sci 106:10386–10391
Fontana S, Sattler T, Bontadina F, Moretti M (2011) How to manage the urban green to improve bird diversity and community structure. Landsc Urban Plan 101:278–285
Goddard MA, Dougill AJ, Benton TG (2010) Scaling up from gardens: biodiversity conservation in urban environments. Trends Ecol Evol 25:90–98
ACT Government 2006. Design standards for urban infrastructure. 14. Urban Open Space. Territory and Municipal Services, ACT
Hanspach J, Fischer J, Stott J, Stagoll K (2011) Conservation management of eastern Australian farmland birds in relation to landscape gradients. J Appl Ecol 48:523–531
Hostetler M, Holling CS (2000) Detecting the scales at which birds respond to structure in urban landscapes. Urban Ecosyst 4:25–54
Huste A, Boulinier T (2011) Determinants of bird community composition on patches in the suburbs of Paris, France. Biol Conserv 144:243–252
Laurance WF (2008) Theory meets reality: how habitat fragmentation research has transcended island biogeographic theory. Biol Conserv 141:1731–1744
Lim HC, Sodhi NS (2004) Responses of avian guilds to urbanisation in a tropical city. Landsc Urban Plan 66:199–215
Lindenmayer DB, Cunningham RB (2011) Longitudinal patterns in bird reporting rates in a threatened ecosystem: is change regionally consistent? Biol Conserv 144:430–440
Lindenmayer DB, Fischer J (2006) Habitat fragmentation and landscape change. An ecological and conservation synthesis. CSIRO Publishing, Collingwood
Loss SR, Ruiz MO, Brawn JD (2009) Relationships between avian diversity, neighborhood age, income, and environmental characteristics of an urban landscape. Biol Conserv 142:2578–2585
Luck GW, Davidson P, Boxall D, Smallbone L (2011) Relations between urban bird and plant communities and human well-being and connection to nature. Conserv Biol 25:816–826
McCullagh P, Nelder JA (1989) Generalized linear models. Chapman & Hall/CRC, Boca Raton
Montague-Drake R, Lindenmayer DB, Cunningham R (2009) Factors effecting site occupancy by woodland bird species of conservation concern. Biol Conserv 142:2896–2903
Musacchio LR (2009) The scientific basis for the design of landscape sustainability: a conceptual framework for translational landscape research and practice of designed landscapes and the six Es of landscape sustainability. Landscape Ecol 24:993–1013
Myint SW, Gober P, Brazel A, Grossman-Clarke S, Weng Q (2011) Per-pixel vs. object-based classification or urban land cover extraction using high spatial resolution imagery. Remote Sens Environ 115:1145–1161
Oliver AJ, Hong-Wa C, Devonshire J, Olea KR, Rivas GF, Gahl MK (2011) Avifauna richness enhanced in large, isolated urban parks. Landsc Urban Plan 102:215–225
Ortega-Alvarez R, MacGregor-Fors I (2009) Living in the big city: effects of urban land-use on bird community structure, diversity and composition. Landsc Urban Plan 90:189–195
Pauleit S, Ennos R, Golding Y (2005) Modeling the environmental impacts of urban land use and land cover change—a study in Merseyside, UK. Landsc Urban Plan 71:295–310
Rey Benayas JM, Bullock JM, Newton AC (2008) Creating woodland islets to reconcile ecological restoration, conservation and agricultural land use. Front Ecol Environ 6:329–426
Rhodes J, McAlpine C, Zuur A, Smith G, Ieno E (2009) GLMM applied on the spatial distribution of koalas in a fragmented landscape. In: Zuur A (ed) Mixed effects models and extensions in ecology with R. Springer, Berlin
Robertson OJ, Radford JQ (2009) Gap-crossing decisions of forest birds in a fragmented landscape. Austral Ecol 34:435–446
Shwartz A, Shirley S, Kark S (2008) How do habitat variability and management regime shape the spatial heterogeneity of birds within a large Mediterranean urban park? Landsc Urban Plan 84:219–229
Silcocks A, Tzaros C, Weston M, Olsen P (2005) An interim guild classification for woodland and grassland birds in Australia. Birds Australia Supplementary Report to State of the Environment Report 2006, Carlton
Stagoll K, Lindenmayer DB, Knight E, Fischer J, Manning AD (2012) Large trees are keystone structures in urban parks. Conserv Lett 5:115–122
Suarez-Rubio M, Thomlinson JR (2009) Landscape and patch-level factors influence bird communities in an urbanized tropical island. Biol Conserv 142:1311–1321
Sutherland WJ, Newton I, Green RE (2004) Bird ecology and conservation: a handbook of techniques. Oxford University Press, Oxford
Thornton DH, Branch LC, Sunquist ME (2011) The relative influence of habitat loss and fragmentation: do tropical mammals meet the temperate paradigm? Ecol Appl 21:2324–2333
Tratalos J, Fuller RA, Warren PH, Davies RG, Gaston KJ (2007) Urban form, biodiversity potential and ecosystem services. Landsc Urban Plan 83:308–317
Tremblay MA, St. Clair CC (2011) Permeability of a heterogeneous urban landscape to the movements of forest songbirds. J Appl Ecol 48:679–688
United Nations 2009 (2009) Revision of World Urbanization Prospects. In: United Nations Development Programme
United Nations 2011 (2011) Revision of world urbanization prospects. In: United Nations Department of Economic and Social Affairs P. D.
Wiens J (2009) Landscape ecology as a foundation for sustainable conservation. Landscape Ecol 24:1053–1065
Acknowledgments
Thanks to staff from ACT Conservation, Planning and Research, Territory and Municipal Services; Land Development Agency, ACT Planning and Land Authority, and J. Hibberd, P. Barton and P. Lentini for useful discussions; and M. Westgate and I. Stirnemann for providing helpful comments on earlier versions of this paper. K.I. was the recipient of a postgraduate independent research scholarship, jointly funded by the Fenner School of Environment and Society (The Australian National University) and Conservation, Planning and Research (ACT Government). Ethics approval was obtained before conducting this work (F.ES.08.10).
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Ikin, K., Beaty, R.M., Lindenmayer, D.B. et al. Pocket parks in a compact city: how do birds respond to increasing residential density?. Landscape Ecol 28, 45–56 (2013). https://doi.org/10.1007/s10980-012-9811-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-012-9811-7