
Abstract
We analysed the effect of the urban matrix, the urban space surrounding distinct habitat patches, on bird communities. In doing so we assessed the impact of urbanisation beyond the effect of habitat loss. We used a set of 54 wasteland sites of early successional stages that were scattered over the entire urban area of Berlin, Germany. Sites were similar to each other in habitat structure but differed in their surroundings, the urban matrix. Thus, our study design allows to investigate associations between birds and the urban matrix. Our measures for urbanisation are human population density and degree of sealing within 50 to 2,000 m buffer zones surrounding each wasteland site. Along the urbanisation gradients we calculated three measures of bird communities: alpha diversity, beta diversity, and trait profile of the entire bird community regarding food, life-history, and behavioural traits. Alpha diversity did not change significantly along the gradients of urbanisation. However, beta diversity increased along the urbanisation gradients with urbanisation at the local scale (50 m) but decreased at the landscape scale (200 and 2,000 m). Fourth-corner analysis of relationships between urbanisation and species traits showed trait shifts: adult survival rate increased with human population density and densities of birds that are more often reported to show innovative behaviour increased with both human population density and degree of sealing. We conclude that the influence of the urban matrix contributes to the homogenisation of the avifauna by filtering certain species traits and promoting others.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Urbanisation is rapidly changing our world’s face, reducing native habitats (Wackernagel and Rees 1996; Benfield et al. 1999; Czech et al. 2000; Sala et al. 2000) and thus the richness and composition of native bird species (Marzluff 2001; McKinney 2008). But what is urbanisation exactly, and what are the associated effects of urbanisation on birds? Urbanisation is a process of population concentration (Tisdale 1941) and is thus attended by transformation of land with near-natural habitats or agricultural areas, to land that is used for non-agricultural production, housing, or trading. These changes are known to reduce the species number of birds (reviews in Marzluff 2001; Chace and Walsh 2006; McKinney 2008). In most urban studies landscape modification and habitat availability are the most relevant factors explaining the variations in species distribution (Batten 1972; Clergeau et al. 2001; Melles et al. 2003; Donnelly and Marzluff 2006; Sandström et al. 2006; Croci et al. 2008). Urbanisation seems to cause changes in species communities by reducing appropriate habitat types; if remnants of near-natural habitats remain, also most bird species do (Haire et al. 2000). Previous studies often used pairwise urban versus non-urban comparisons or spatial transect approaches. Since urban bird communities have been shown to be independent of the bird diversity of adjacent landscapes (Clergeau et al. 2001), pairwise comparisons of urban with non-urban bird communities may not reflect the influence of urbanisation. Spatial transect approaches are not applicable to every urban area since cities are not always strictly concentric and thus do not have gradual spatial gradients (Alberti et al. 2001). Instead, many urban agglomerations are patchy, highly fragmented, and discontinuous. Additionally, most urban areas, particularly in Central and Southern Europe, are exclusively man-made and do not comprise of any appreciable remnant primary habitats. Consequently, bird species living in these landscapes either possess adaptations enabling them to cope with the conditions of urbanised areas (Croci et al. 2008) or are pre-adapted to the urban habitat features.
Whereas most studies report a decline in species diversity along the rural-to-urban gradient, some studies found highest species diversities at moderately disturbed sites (e.g., Blair 1996; Marzluff 2005; Blair and Johnson 2008). On such sites, near-natural habitats are fragmented by urbanisation but remnant primary habitat patches such as natural woodland are left. Hence, at intermediate levels of urbanisation the pristine avifauna is enriched by exogenous species that benefit from these new conditions. In urban areas without remnants of near-natural vegetation the species number depends mainly on the diversity of available habitats (e.g., Jokimäki and Suhonen 1998).
Urbanisation is also discussed to cause biotic homogenisation by replacing regionally distinct communities with cosmopolitan communities (McKinney and Lockwood 1999; Clergeau et al. 2006a). Taking the concept of Blair (1996), some species, called ‘urban avoiders’, disappear along with urbanisation while others, called ‘urban exploiters’, invade and thrive; in between there is a large group of ‘suburban adaptables’ that are able to inhabit novel man-made habitats of intermediate urbanisation levels. Blair (2004) verified this homogenisation effect. He found that the similarity of species composition between two cities was positively correlated with the degree of urbanisation. Here, the categories urban avoider and urban exploiter are used as tools to group species which does not imply any further biological or evolutionary characterisation. Since urbanisation is a very complex process, drivers of homogenisation are still not sufficiently explored and show varying effects on different species groups (McKinney and Lockwood 1999; Olden 2006). Biotic homogenisation is a temporal process. Biotic homogenisation is usually considered as a temporal process. We use here a space-for-time-substitution approach to investigate this phenomenon.
We consider a habitat as a templet sensu Southwood (1977, 1988) that acts as a filter for functional traits, e.g., foraging technique or nest location. There is evidence that this applies to birds (Petchey et al. 2007; Croci et al. 2008). Analysing traits instead of species reveals how the selection pressure, in our case the urban environment, shapes the community. The traits we selected describe food, life-history, and behaviour.
Urban habitats are reported to favour habitat generalists, omnivorous, granivorous, and cavity nesting bird species, but to decrease the abundance of ground nesting birds, multiple breeders, sedentaries, and those preferring bush-shrub habitats (Jokimäki and Suhonen 1993; Chace and Walsh 2006; Clergeau et al. 2006a; Kark et al. 2007; Blair and Johnson 2008; Devictor et al. 2008; Møller 2009). Again, most studies were not able to separate the effects of change in habitat availability and influence of the surroundings. To the best of our knowledge there is no systematic research on connections between the urban matrix and bird species traits.
Many studies are not able to distinguish between the influences of on-site characteristics, i.e., habitat quality and quantity, and the influence of the adjacent urban matrix on bird communities. We attempt to separate these two aspects by analysing similar urban habitats and determining the influence of the urban matrix on species number, species shifts, and trait shifts along the urban gradient. Since animals respond to environmental conditions on multiple scales (Jokimaki and Huhta 1996), we collected data from the patch level (fine-scale vegetation structure) to the landscape level (degree of urbanisation within 2,000 m).
To investigate the influence of the urban matrix on bird communities we analysed wasteland sites in early successional stages that were spread over the city of Berlin, Germany. We assume our study sites to have similar filtering characteristics that differ in many respects from the surrounding matrix, even from other green spaces: The wastelands feature sparse vegetation, few trees and shrubs and are thus selecting for open-land birds, ground gleaners etc. The surrounding urban matrix on the other hand may also act as a filter, either as a barrier or as part of the habitat that is used by the birds. Whereas the filtering characteristics of the study sites are similar, the degrees of urbanisation in the surroundings vary. Our setting enables us to test how the surroundings of a site influence its species and trait composition.
In a first step, we evaluate the relative influences of features of the habitat patch and the surrounding urban matrix, respectively, on species composition of the habitat patch. Second, we assess the influence of the urban matrix on species diversity and trait composition. We assume that (1) the urban matrix has an impact on species composition beside the effect of local habitat variables. We further hypothesise that with an increase of urbanisation of the matrix, that is, human population density and degree of sealing in the surroundings of habitat patches, (2) alpha diversity (i.e., area-corrected species number) decreases since there might be less urban exploiters that are added than urban avoiders that disappear, (3) beta diversity (i.e., dissimilarity of bird communities between sites) decreases since urban exploiters become more abundant, and (4) there are trait shifts from low to highly urbanised areas. Based on previous findings from the literature we hypothesise that urban exploiters, i.e., bird species that are able to cope with highly urban surroundings, (a) use artificial food sources and are thus rather omnivorous than insectivorous (Croci et al. 2008), (b) forage mainly on the ground since artificial food often can be found there, (c) show more novel ways of behaviour (Møller 2009), and (d) are rather sedentary than migratory (Kark et al. 2007; Blair and Johnson 2008; Croci et al. 2008) since urban conditions may allow them to stay in winter and occupy suitable territories before the migrants arrive. Figure 1 illustrates our study design, calculation of beta diversity, and its possible relationships to the degree of urbanisation.

Diagram of the study design (a) and possible relationships between the degree of urbanisation within the urban matrix and beta diversity between similar habitat patches (b)
Methods
Study area and site selection
All 54 study sites were located in the urban area of Berlin (52°30′2″ N 13°23′56″ E), with 3.4 million inhabitants and 892 km² it is the largest city of Germany (Amt für Statistik Berlin-Brandenburg 2010). Wastelands were located by asking at the municipality and scanning aerial photographs (Senate Department for Urban Development 2005). The study sites were spread over the whole urban area with some more sites located in the eastern part of the city and in the city centre due to historical reasons. Since Berlin emerged from several towns and villages it is until now very diverse and has several centres. The sites had various former uses, such as switching yard stations, industrial plants, or buildings that had been demolished. All sites were dominated by sparse vegetation with no or few trees and shrubs.
We selected all of the largest available wasteland sites. In addition, we chose smaller ones to cover gradients of size and degree of urbanisation. Beyond our study sites we assume this habitat type to be rare and equally distributed within the urban area of Berlin.
Bird abundance data
Birds were mapped in 2007 by visiting each of the 54 study sites four times from end of April to end of July. Each site was inspected three times in the morning and once in the late afternoon during the second activity peak (Aschoff 1966). We switched the order of visits to avoid bias caused by changing activity of birds with time of the day. Birds were easy to detect for the sparse vegetation of the sites. We did not correct for different detectability since overall detectability was very high. Moreover, habitat characteristics and species pools were similar among study sites, hence, selective bias due to differing detection probabilities is expected to be low. The field survey was conducted by the same experienced person. All birds were noted by walking slowly across the complete area at good weather conditions with no rain, no or little wind, and not above about 28 °C in the afternoon for low activity in birds at high temperatures (Kendeigh 1969). All present birds were mapped except barn-swallow Hirundo rustica, house martin Delichon urbicum, and swift Apus apus since these were permanently flying and thus less likely to be relatable to the study sites.
We calculated densities by summing up bird numbers of the four visits and dividing them by the size of each site. Densities were square-root transformed to eliminate the influence of very high densities due to small sites or large flocks. We analysed only species that occurred on at least three of the 54 sites to exclude the influence of very rare species.
Bird trait data
We define a trait as a species-specific characteristic. The selected traits are related to bird‘s physiology, behaviour, and life-history. Notably, here we include behavioural traits such as foraging or migration strategy as done, e.g., by Sol et al. (2012); thus, our definition is broader than that of other authors (e.g., Violle et al. 2007).
Five traits were selected: food type, foraging technique, adult survival, innovation rate, and migration strategy. All values are shown in Table 1. Data except adult survival and innovation rate were taken from Glutz von Blotzheim (1985). Data on adult survival were taken from BTO (2010). We employed innovation rate as a proxy for behavioural flexibility. It was calculated from numbers of novel behaviours reported in journals that are included in the Zoological Records’ web index published by Overington et al. (2009). To account for the bias in observation effort, we divided the number of reported innovations by the overall number of papers published on a given species in the Zoological Records’ web index. We assumed the innovation rate of species without entries to be zero since the literature search of Overington et al. (2009) was very exhaustive.
Birds were classified as long-distance migrants if they usually winter in Africa. For the trait analysis we excluded mallard Anas platyrhynchos and kestrel Falco tinnunculus since these were the only representatives in their trait categories for food (herbivorous and carnivorous) and for foraging technique (dabbler, air hawker). We did not account for phylogenetic relatedness since it explains less than 1 % of the variation among species for most behavioural and ecological traits (Böhning-Gaese and Oberrath 1999).
Adult survival and food were correlated by 0.65, the other traits <0.3 (Pearson correlation for quantitative data and Spearman correlation for categorical data).
Habitat data
For each study site, we mapped vegetation and surface structure and calculated proportions of each surface type per site. Additionally, the following measures were used to assess characteristics of the waste land sites: size of the study site, presence of trees adjacent to the sites, index of presence and consistency of humans, and presence of cats. Cats are modelled as present or absent. Numbers of humans and cats were mapped during bird surveys. Index of humans as approximation for direct disturbance by human intrusion was calculated as mean number of humans per visit and hectare multiplied by the degree of presence, that is, one to four visits where we were able to detect humans on the site.
Urban matrix data
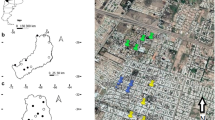
Regarding the spatial relationship between birds and their environment, a multi-level approach is often necessary for understanding species response in patchy systems (Jokimaki and Huhta 1996; Mazerolle and Villard 1999; Clergeau et al. 2006b; Thornton et al. 2011). Therefore, we sampled the urban matrix in buffer zones surrounding the habitat we focussed on. In doing so, we differentiated between three spatial scales: 50, 200, and 2,000 m (Fig. 2). We used degree of sealing and residential population density as approximation for the degree of urbanisation, since these measurements are often easily accessible and imply many structural features of an urban matrix such as housing type or vegetation cover (Alberti et al. 2001; Senate Department for Urban Development 2006). Data was taken from Environmental Atlas from the municipality of Berlin (Senate Department for Urban Development 2004, 2006). Proportion of sealed area (i.e., impervious surface) and residential population density were calculated using ArcMap 9.2. We excluded variables with a Spearman correlation coefficient ≥0.6 (absolute value) resulting in four remaining variables: degree of sealing within 50 and 2,000 m and human population density within 50 and 200 m.

Aerial image of an exemplary wasteland site (shaded) and three buffer zones (50, 200, and 2,000 m) for which population density and degree of sealing were calculated (Orthophotomosaike Berlin, © GeoBasis-DE/SenStadtUm III, 2005)
Statistical analyses
To show the independence of the features of the wasteland sites on their surroundings, we tested the overall correlation of the on-site habitat variables (area size, vegetation, disturbances) with the urbanisation variables (sealing and human population density of the surrounding urban matrix) using a Mantel statistic based on Kendall’s rank correlation tau and 3,000 permutations.
To estimate the influence of on-site variables and matrix features on species composition we first calculated a detrended correspondence analysis (DCA) to estimate the gradient length covered by the data. A length of first DCA axis <2 indicates a linear response, a length of >4 a unimodal response (Lepš and Šmilauer 2003). In our case the first DCA axis had a length of 3.4, however, all but one site covered a range of 2.1. Therefore we chose a redundancy analysis (RDA) as ordination technique that assumes the response to be linear. Species were scaled proportional to eigenvalues to assess the influence on all species equally and not only on the most variable and abundant ones (Oksanen 2011); sites remained unscaled. We did a partitioning of variance to quantify the influence of the different variable sets (habitat structure, degree of urbanisation) on bird communities. Therefore we used the function varpart from the vegan package that is based on RDA (Oksanen et al. 2010) but uses semi-partial R-squares which is the only unbiased method (Peres-Neto et al. 2006). To test the significance of the influence of the independent fractions, we used the function anova in varpart, that is, a permutation test with 999 permutations. This test shows whether a fraction of variables significantly explains variation in species composition. All predictor variables are given in Table 2.
Since our study sites varied in size, we corrected the species number for size dependency. Species number is known to be linked to area size by a power function (MacArthur and Wilson 1963). To assess the alpha diversity, we first applied a power function regression with the formula species number = b·area size z to the overall dependency of species number on area size with b = 7.4788, z = 0.3407, p < 0.0001, R2 = 0.748. Afterwards we calculated residuals from the species-area relationship as the difference of the observed species number to the predicted species number per site from this relationship as an area size independent measure of alpha diversity.
We took the Bray-Curtis dissimilarity as measure for beta diversity because it is insensitive to variation in median species number between sites and avoids the ‘double-zero problem’, i.e., if species are absent on both sites that are compared (Faith et al. 1987; Leyer and Wesche 2007). Beta diversity was calculated as running mean of Bray-Curtis dissimilarities along the urban gradients (Fig. 1). For this purpose we calculated (1) the mean of the dissimilarities of the 15 sites with lowest degree of urbanisation, (2) the mean for the second least to the 16th least urban site and so on, resulting in 40 averaged values for the 54 sites. Values of the ordinate have been calculated similarly. All analyses were done using the package vegan (Oksanen et al. 2010) implemented in the R software (R Development Core Team 2009).
To relate bird species traits to the four urban matrix variables we used the fourth-corner test because of its statistical power (Dray and Legendre 2008). This statistical method allows measuring and testing directly the links between the variations in bird traits and the urban matrix variables representing the degree of urbanisation at two different spatial scales. Similar to the RLQ analysis (Dolédec et al. 1996), the fourth-corner statistic measures the link between three tables: a table L (n × p) containing the abundances of p species at n sites, a second table R (n × m) with the measurements of m environmental variables for the n sites, and a third table Q (p × s) describing s species traits for the p species, i.e., bird traits and environmental variables are linked by the site-species matrix. By permutating sites and species it is possible to test for significant relationships between single traits of birds and environmental features, whereas the RLQ analysis provides only a p value for the overall relation between species traits and environmental variables. To test the statistical significance of the overall relationship between urbanisation and trait variables we computed a Monte-Carlo randomisation test with 99,999 permutations (Dray et al. 2002). We used the approach of Gallardo et al. (2009) by first permuting the site vectors to test null hypothesis 1 that species assemblages are not dependent upon the characteristics of the site’s surroundings where they are actually found (permutation model 2 of Dray and Legendre 2008). After that, we permuted species vectors to test null hypothesis 2 that the distributions of the species among the sites, which are related to their preferences for site conditions, does not depend on the traits of the species (permutation model 4 of Dray and Legendre 2008). Each calculation was done with 9,999 permutations. The largest p value of the two models is given here. Because of multiple testing of eight environmental variables, the alpha level had to be Bonferroni-corrected. The combined use of the two permutation models requires the square-root of this corrected alpha level to be used for the single tests (Dray and Legendre 2008; Gallardo et al. 2009). Consequently, we used p < 0.129 for alpha = 0.05 as an indication of statistical significance in Table 4. Statistical analyses were performed using the ade4 package (Dray and Dufour 2007).
A generalised linear model of square-root transformed sums of individuals with size of each site as offset term was applied to estimate shifts of bird densities along the matrix gradients with categorical traits. Numerical traits (adult survival and innovation rate) were unimodal distributed, therefore we were able to calculate trait profiles of the bird communities as median values per site weighted by abundances. Changes in trait abundances were tested using linear regression.
Results
There was no significant spatial structuring of the urban matrix according to the habitat features of the wasteland sites (Mantel statistic r = 0.017; p = 0.34).
Species composition
Urbanisation, measured by human population density and degree of sealing in the surrounding of each wasteland site, significantly influenced species composition on the wastelands; however, on-site variables like habitat structure or disturbance intensity did not (Fig. 3). According to Økland (1999) relative contributions of the fractions are of importance for interpretation, not overall explained variance.

Venn diagram of explanatory contributions of on-site and matrix variables on bird species composition based on variance partitioning. Fractions Human population density and Degree of sealing did significantly explain variation in species composition (permutation test, p = 0.005**) whereas fractions Habitat structure and Disturbance intensity did not (p = 0.28 and p = 0.95, respectively; not significant (n.s.)). The fractions, representing adjusted R2, are expressed as percentages of the total explained variance. Total explained variance is 9.6 %, this low percentage is not an indicator for low interpretational power, see (Økland 1999) for a discussion
Alpha diversity
Overall we recorded 50 bird species, 32 species occurred on at least three sites representing 4,169 individuals (including barn-swallow and house martin that were excluded from the analysis). Area-corrected species number did not change significantly with urbanisation (Table 3). For interpretation of results we also checked if there was a change in overall bird density and evenness along the gradients, but there was also no indication for a linear or other relationship. No exotic species were observed in the surveys.
Beta diversity
Bray-Curtis dissimilarities among sites as a measure for beta diversity identified a clear scale dependency of the impact of urbanisation on bird communities. While beta diversity increased with urbanisation at the 50 m scale, it decreased at the 200 and 2,000 m scale (Fig. 4).

Median Bray-Curtis dissimilarities along the urbanisation gradients. Dots are running means of 15 sites each for which the dissimilarities are calculated (see Methods section for details)
Traits shifts
There was a significant relationship between species traits and the four measurements of urbanisation (randomisation test, p < 0.0003) that validates the use of the fourth-corner analysis. The fourth-corner analysis gave eight significant relationships between urbanisation proxies and species traits. Out of the five traits, foraging technique, adult survival, and innovation rate were significantly related to characteristics of the urban matrix whereas food and migration strategy were not significantly related (Table 4).
Foraging technique was significantly related to the degree of sealing within 50 m (Table 4). Out of the three foraging strategies, only density of sit-and-wait predators changed significantly along the gradient, with increasing densities with more sealing (Table 5). The densities of sit-and-wait predators per visit increased from 0.017 individuals per hectare at 20 % sealing to 0.226 individuals per hectare at 80 % sealing, i.e., by factor 14.
Out of the three significant relationships between degree of urbanisation and adult survival (Table 4), two show a clear pattern along the urbanisation gradient with more long-living birds at sites with more densely settled surroundings (Table 6).
The innovation rate as a measure for behavioural flexibility was significantly correlated with all four matrix variables (Table 4). Mean innovation rate increased significantly with two urbanisation proxies (Table 6).
Discussion
Species composition
We showed that the influence of the urban matrix on species composition on the studied wastelands is of same dimension as the impact of habitat structure (Fig. 3). Since we did not gather data on species composition outside the wastelands, we are not able to quantitatively compare the wasteland’s avifauna to that of urban parks or the urban matrix. However, we can state that some species bred to our knowledge exclusively on wastelands, e.g., wheatear and tawny pipit, but others are common inhabitants of the urban matrix such as house sparrow or hooded crow. Overall, bird communities on the wastelands seem to be distinct from other urban bird communities.
Both human population density and degree of sealing independently explained equal proportions of variation and were the only significant predictors for species composition. Habitat structure did indeed explain a big proportion of the explained variance (51 %) but this part did not significantly explain any variation in species composition. Possibly, species composition was related to habitat structure but not in a consistent manner, i.e., habitat effects include more noise and variance whereas matrix effects are more consistent. Overlap of buffer zones might have caused some overestimation of the matrix’ influence but we believe this effect to be weak since correlation among buffer zones was low.
The intensity of disturbance by humans and cats accounts only for a small fraction of the variation and was entirely explained by the habitat structure (Fig. 3). Hence, disturbances occurred together with certain habitat features. Consequently, there seems not to be any influence of direct disturbance on birds distinguishable from habitat characteristics.
Overall, the variation-partitioning model explains 9.6 % of the total variation. This may seem quite low, but Økland (1999) showed that the common interpretation of ‘unexplained variation’ as random variation caused by unmeasured explanatory variables, is generally inappropriate. In our interpretation of the results we followed Økland’s recommendation and concentrated on relative amounts of variation explained by different sets of explanatory variables rather than focussing on the explained-to-total-inertia ratio and unexplained variation.
Alpha diversity
Within the covered gradients of urbanisation, alpha diversity did not change significantly (Table 3), indicating that there are similar numbers of species that are able to exploit the resources along the urban gradient. That contrasts to our initial hypothesis that alpha diversity is lower in highly urbanised areas and to many other studies that have found this pattern (e.g., Gavareski 1976; Beissinger and Osborne 1982; Berressem et al. 1983; Cam et al. 2000). A recent study on birds on a global scale also showed that urbanisation does not change species number considerably (Pautasso et al. 2011). If there is a uniform habitat type, as the wastelands in our study, there is apparently not much variation in species number. Hence, the species loss reported in other studies seems to be primarily related to a loss of appropriate habitats connected with urbanisation. However, our further findings on beta diversity and traits shifts suggest a species turnover along the urban gradient. Though, numbers of additional and lost species seem to be balanced, resulting in the observed pattern.
Beta diversity
Beta diversity, i.e., the similarity in species composition and abundance between sites of a specific population density/amount of sealing, did change along the urbanisation gradient. The direction was dependent on the spatial scale (Fig. 4). In contrast to our hypothesis, more urban surroundings at the local scale (50 m) caused more heterogeneous bird communities, i.e., more between-site differences (Fig. 4, left-hand) as suggested by the dashed line in Fig. 1b. In other words, bird communities become more different as the small-scale surroundings of the habitat patch become more sealed and densely populated by humans. Since the studied wastelands differ in many respects from their surroundings—no human residents, low degrees of sealing, open soils, sparse vegetation—it provides resources that are different from the adjacent urban matrix. Thus, changes in the matrix quality are likely to affect bird species composition. Bender and Fahrig (2005) showed that matrix suitability has a strong impact on occurrence of animals. Possibly, with increasing degree of urbanisation, also heterogeneity of the 50 m matrix increases, leading to more differentiated bird communities among the sites.
In contrast, at the landscape scale (200 and 2,000 m), high population densities and degrees of sealing caused more similar bird communities (Fig. 4, right-hand). In agreement with our hypothesis (solid line in Fig. 1b), the urban matrix seems to filter certain species at a spatial scale of several hundred metres to some kilometres.
Trait shifts
We observed shifts in the trait profiles (e.g., trait frequencies per area weighted by abundances) of the bird communities along the urban gradients. Three of the five selected traits were significantly related to features of the urban matrix (Table 4). These traits were only weakly correlated to each other (R2 < 0.3).
In contrast to our initial hypothesis and previous findings for the breeding period (Emlen 1974; Lancaster and Rees 1979; Beissinger and Osborne 1982; Kark et al. 2007; Blair and Johnson 2008; Croci et al. 2008) or during winter (Jokimäki and Suhonen 1998), there was no change in food types along the urban gradient. Thus, food provided by humans seems not to cause an increase in densities of omnivores on the wasteland sites. In addition, there was no decrease of insectivorous or granivorous birds, suggesting that the availability of arthropods and seeds seems not to vary much with the urban matrix. Possibly, the food supply on the wastelands is less determined by the urban matrix but mainly a feature of the wasteland site itself.
Regarding foraging technique, we did not find an increase in ground gleaners as we have expected. Rather, wastelands seem to be used by ground gleaners and tree/foliage gleaners irrespective of the urban matrix. Only the density of sit-and-wait predators increased with the degree of sealing at the 50 m scale. Presumably this is mainly caused by black redstarts Phoenicurus ochrurus that use wasteland sites especially in highly urban areas for foraging. Indeed, black redstarts inhabit all the urban area (Otto and Witt 2002) but in highly sealed areas they presumably make increased use of urban wastelands since other open habitats are more rare.
Habitats next to high human population densities selected for long-living birds (Table 6). A similar trend was revealed by pairwise comparison of closely related urban versus rural bird species (Møller 2009). A longer lifetime may better enable birds to adapt by individual learning that might be more important in urban settings compared to more stable natural habitats.
Our results support previous findings that species with behavioural flexibility have a better invasion success in novel habitats (Sol et al. 2002) and thrive in urban areas (Yaukey 1996; Møller 2009). This seems plausible as urban environments are highly variable in space and time and it should be a valuable attribute to be able to find novel ways in exploiting resources such as food or nesting sites. Our findings suggest that this kind of ‘cleverness’ is beneficial irrespective of other traits such as food type or migration strategy.
Contrary to our hypothesis and to other findings (Kark et al. 2007; Blair and Johnson 2008; Croci et al. 2008), there was no clear shift in migratory status along the urbanisation gradients. Composition of migration strategies seems to result from associations between species and habitats; as long as the habitat type is available, the urban matrix seems not to favour a certain migration strategy.
Biotic homogenisation
Our finding of decreasing beta diversity with increasing urbanisation at the 200 and 2,000 m scale indicates local biotic homogenisation. It is likely that the causes for this local and spatial homogenisation contribute to a larger-scale and temporal biotic homogenisation. Olden and Poff (2003) derived several explanatory mechanisms for biotic homogenisation from theoretical considerations and simulation models. They address the phenomenon of biotic homogenisation to invasion of non-native species and extinction of native species, respectively, as found for North America (Blair 1996, 2004; Blair and Johnson 2008). On a global scale, biotic homogenisation seems not to be driven by the invasion of exotic species (Pautasso et al. 2011). Although we did not find any non-native species in our study—a common finding for Central Europe (e.g., Clergeau et al. 2006a)—the concept of Olden and Poff is transferable. Thus, the following mechanisms could explain lower beta diversity in highly urban areas at the landscape scale: (1) local invasion of urban exploiters, that is, species that profit from a highly urbanised matrix and/or (2) local extinction of urban avoiders, that is, species that are not able to cope with strongly urban conditions.
These two processes may however not be simultaneous and could occur at different speeds. Bird extinctions could have occurred much before invasions—in fact, many habitat specialists may have been removed as soon as the matrix has been modified and never recolonized afterwards. In contrast, generalist, flexible species may have colonized recently, and probably no earlier than when the land was abandoned.
Simulation models of Olden and Poff (2003) supported empirical findings that homogenisation is greatest when similar species invade communities. Our findings suggest that long-living, innovative species invade the more urban sites while short-living and less innovative species disappear. As a result, bird communities of urbanised landscapes and thus the regional species pools become more homogenous.
Conclusions
We showed that the urban matrix causes changes in the avifauna beyond the changes caused by direct habitat modification. Human population density and degree of sealing of the urban matrix affected diversity patterns, species composition, and trait frequencies of birds. The observed shifts in species and traits are likely to contribute to the phenomenon of biotic homogenisation of birds which was found for Italian towns (Sorace and Gustin 2008), across Europe (Clergeau et al. 2006a), and worldwide (Lockwood et al. 2000). If certain traits decrease (e.g., short life-span) while others become more frequent (e.g., behavioural flexibility), urban-adapted communities will become more similar along the local urban gradient as well as at a continental or worldwide scale.
References
Alberti M, Botsford E, Cohen A (2001) Quantifying the urban gradient: linking urban planning and ecology. Avian ecology and conservation in an urbanizing world. Kluwer Academic, Norwell, pp 89–115
Amt für Statistik Berlin-Brandenburg (2010). http://www.statistik-berlin-brandenburg.de/. Accessed 5 May 2010
Aschoff J (1966) Circadian activity pattern with two peaks. Ecology 47(4):657–662
Batten LA (1972) Breeding bird species diversity in relation to increasing urbanisation. Bird Study 19(3):157–166
Beissinger S, Osborne D (1982) Effects of urbanization on avian community organization. Condor 84(1):75–83
Bender DJ, Fahrig L (2005) Matrix structure obscures the relationship between interpatch movement and patch size and isolation. Ecology 86(4):1023–1033
Benfield FK, Raimi M, Chen DDT (1999) Once there were greenfields: how urban sprawl is undermining America’s environment, economy, and social fabric. Natural Resources Defense Council, New York
Berressem KG, Berressem H, Schmidt K-H (1983) Vergleich der Brutbiologie von Höhlenbrütern in innerstädtischen und stadtfernen Biotopen. J Ornithol 124(4):431–445
Blair R (1996) Land use and avian species diversity along an urban gradient. Ecol Appl 6(2):506–519
Blair R (2004) The effects of urban sprawl on birds at multiple levels of biological organization. Ecol Soc 9(5):5
Blair R, Johnson E (2008) Suburban habitats and their role for birds in the urban-rural habitat network: points of local invasion and extinction? Landscape Ecol 23(10):1157–1169
Böhning-Gaese K, Oberrath R (1999) Phylogenetic effects on morphological, life-history, behavioural and ecological traits of birds. Evol Ecol Res 1(3):347–364
BTO (2010) BirdFacts. British Trust for Ornithology. http://blx1.bto.org/birdfacts/indexa_short.htm. Accessed 5 Oct 2010
Cam E, Nichols J, Sauer J, Hines J, Flather C (2000) Relative species richness and community completeness: birds and urbanization in the mid-Atlantic states. Ecol Appl 10(4):1196–1210
Chace J, Walsh J (2006) Urban effects on native avifauna: a review. Landsc Urban Plan 74(1):46–69
Clergeau P, Jokimäki J, Savard J-PL (2001) Are urban bird communities influenced by the bird diversity of adjacent landscapes? J Appl Ecol 38(5):1122–1134
Clergeau P, Croci S, Jokimäki J, Kaisanlahti-Jokimäki M-L, Dinetti M (2006a) Avifauna homogenisation by urbanisation: analysis at different European latitudes. Biol Conserv 127(3):336–344
Clergeau P, Jokimäki J, Snep R (2006b) Using hierarchical levels for urban ecology. Trends Ecol Evol 21(12):660–661
Croci S, Butet A, Clergeau P (2008) Does urbanization filter birds on the basis of their biological traits? Condor 110(2):223–240
Czech B, Krausman PR, Devers PK (2000) Economic associations among causes of species endangerment in the United States. Bioscience 50(7):593–601
Devictor V, Julliard R, Clavel J, Jiguet F, Lee A, Couvet D (2008) Functional biotic homogenization of bird communities in disturbed landscapes. Glob Ecol Biogeogr 17(2):252–261
Dolédec S, Chessel D, ter Braak CJF, Champely S (1996) Matching species traits to environmental variables: a new three-table ordination method. Environ Ecol Stat 3(2):143–166
Donnelly R, Marzluff J (2006) Relative importance of habitat quantity, structure, and spatial pattern to birds in urbanizing environments. Urban Ecosyst 9(2):99–117
Dray S, Dufour A (2007) The ade4 package: implementing the duality diagram for ecologists. J Stat Softw 22(4):1–20
Dray S, Legendre P (2008) Testing the species traits-environment relationships: the fourth-corner problem revisited. Ecology 89(12):3400–3412
Dray S, Pettorelli N, Chessel D (2002) Matching data sets from two different spatial samples. J Veg Sci 13(6):867–874
Emlen J (1974) An urban bird community in Tucson, Arizona: derivation, structure, regulation. Condor 76(2):184–197
Faith D, Minchin P, Belbin L (1987) Compositional dissimilarity as a robust measure of ecological distance. Plant Ecol 69(1):57–68
Gallardo B, Gascón S, García M, Comín F (2009) Testing the response of macroinvertebrate functional structure and biodiversity to flooding and confinement. J Limnol 68(2):315–326
Gavareski C (1976) Relation of park size and vegetation to urban bird populations in Seattle, Washington. Condor 78(3):375–382
Glutz von Blotzheim UN (1985) Handbuch der Vögel Mitteleuropas. AULA-Verlag, Wiesbaden
Haire S, Bock C, Cade B, Bennett B (2000) The role of landscape and habitat characteristics in limiting abundance of grassland nesting songbirds in an urban open space. Landsc Urban Plan 48(1–2):65–82
Jokimaki J, Huhta E (1996) Effects of landscape matrix and habitat structure on a bird community in northern Finland: a multi-scale approach. Ornis Fennica 73(3):97–113
Jokimäki J, Suhonen J (1993) Effects of urbanization on the breeding bird species richness in Finland: a biogeographical comparison. Ornis Fennica 70:71–77
Jokimäki J, Suhonen J (1998) Distribution and habitat selection of wintering birds in urban environments. Landsc Urban Plan 39(4):253–263
Kark S, Iwaniuk A, Schalimtzek A, Banker E (2007) Living in the city: can anyone become an ‘urban exploiter’? J Biogeogr 34(4):638–651
Kendeigh S (1969) Energy responses of birds to their thermal environments. Wilson Bull 81(4):441–449
Lancaster R, Rees W (1979) Bird communities and the structure of urban habitats. Can J Zool 57(12):2358–2368
Lepš J, Šmilauer P (2003) Multivariate analysis of ecological data using CANOCO. Cambridge University Press, Cambridge
Leyer I, Wesche K (2007) Multivariate Statistik in der Ökologie: Eine Einführung. Springer, Berlin
Lockwood J, Brooks T, Mckinney M (2000) Taxonomic homogenization of the global avifauna. Anim Conserv 3(1):27–35
MacArthur RH, Wilson EO (1963) An equilibrium theory of insular zoogeography. Evolution 17(4):373–387
Marzluff JM (2001) Worldwide urbanization and its effects on birds. In: Marzluff JM, Bowman R, Donnelly R (eds) Avian ecology and conservation in an urbanizing world. Kluwer Academic, Norwell, pp 19–47
Marzluff J (2005) Island biogeography for an urbanizing world: how extinction and colonization may determine biological diversity in human-dominated landscapes. Urban Ecosyst 8:157–177
Mazerolle M, Villard M (1999) Patch characteristics and landscape context as predictors of species presence and abundance: a review. Ecoscience 6(1):117–124
McKinney M (2008) Effects of urbanization on species richness: a review of plants and animals. Urban Ecosyst 11(2):161–176
McKinney M, Lockwood J (1999) Biotic homogenization: a few winners replacing many losers in the next mass extinction. Trends Ecol Evol 14(11):450–453
Melles S, Glenn S, Martin K (2003) Urban bird diversity and landscape complexity: species-environment associations along a multiscale habitat gradient. Conserv Ecol 7(1):5
Møller A (2009) Successful city dwellers: a comparative study of the ecological characteristics of urban birds in the Western Palearctic. Oecologia 159(4):849–858
Økland RH (1999) On the variation explained by ordination and constrained ordination axes. J Veg Sci 10(1):131–136
Oksanen J (2011) Multivariate analysis of ecological communities in r: vegan tutorial. pp. 42
Oksanen J, Kindt R, Legendre P, O’Hara B, Simpson GL, Solymos P, Stevens MHH, Wagner H (2010) Vegan: community ecology package, version 1.17-5. http://vegan.r-forge.r-project.org/. Accessed 23 Nov 2010
Olden J (2006) Biotic homogenization: a new research agenda for conservation biogeography. J Biogeogr 33(12):2027–2039
Olden J, Poff N (2003) Toward a mechanistic understanding and prediction of biotic homogenization. Am Nat 162(4):442–460
Otto W, Witt K (2002) Verbreitung und Bestand Berliner Brutvögel. Berliner Ornithologischer Bericht 12(special edition):1–256
Overington S, Morand-Ferron J, Boogert N, Lefebvre L (2009) Technical innovations drive the relationship between innovativeness and residual brain size in birds. Anim Behav 78(4):1001–1010
Pautasso M, Böhning Gaese K, Clergeau P, Cueto VR, Dinetti M, Fernández-Juricic E, Kaisanlahti-Jokimäki M-L, Jokimäki J, McKinney ML, Sodhi NS, Storch D, Tomialojc L, Weisberg PJ, Woinarski J, Fuller RA, Cantarello E (2011) Global macroecology of bird assemblages in urbanized and semi natural ecosystems. Glob Ecol Biogeogr 20:426–436
Peres-Neto PR, Legendre P, Dray S, Borcard D (2006) Variation partitioning of species data matrices: estimation and comparison of fractions. Ecology 87(10):2614–2625
Petchey OL, Evans KL, Fishburn IS, Gaston KJ (2007) Low functional diversity and no redundancy in British avian assemblages. J Anim Ecol 76(5):977–985
R Development Core Team (2009) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Sala OE, Chapin FS, Armesto JJ, Berlow E, Bloomfield J, Dirzo R, Huber-Sanwald E, Huenneke LF, Jackson RB, Kinzig A, Leemans R, Lodge DM, Mooney HA, Oesterheld M, Poff NLR, Sykes MT, Walker BH, Walker M, Wall DH (2000) Global biodiversity scenarios for the year 2100. Science 287(5459):1770–1774
Sandström UG, Angelstam P, Mikusinski G (2006) Ecological diversity of birds in relation to the structure of urban green space. Landsc Urban Plan 77(1–2):39–53
Senate Department for Urban Development (2004) Berlin digital environmental atlas, 01.02 impervious soil coverage (sealing of soil surface) (Edition 2007)
Senate Department for Urban Development (2005) DOP025-C/DVD 04.3, Orthophotomosaike Berlin - Bildflug August 2004. Berlin
Senate Department for Urban Development (2006) Berlin digital environmental atlas, 06.06 population density (Edition 2006)
Sol D, Timmermans S, Lefebvre L (2002) Behavioural flexibility and invasion success in birds. Anim Behav 63(3):495–502
Sol D, Maspons J, Vall-llosera M, Bartomeus I, García-Peña GE, Piñol J, Freckleton RP (2012) Unraveling the life history of successful invaders. Science 337(6094):580–583
Sorace A, Gustin M (2008) Homogenisation processes and local effects on avifaunal composition in Italian towns. Acta Oecologica 33(1):15–26
Southwood T (1977) Habitat, the templet for ecological strategies? J Anim Ecol 46:337–365
Southwood T (1988) Tactics, strategies and templets. Oikos 52(1):3–18
Thornton D, Branch L, Sunquist M (2011) The influence of landscape, patch, and within-patch factors on species presence and abundance: a review of focal patch studies. Landscape Ecol 26(1):7–18
Tisdale H (1941) Process of urbanization. Soc Forces 20:311
Violle C, Navas M-L, Vile D, Kazakou E, Fortunel C, Hummel I, Garnier E (2007) Let the concept of trait be functional! Oikos 116(5):882–892
Wackernagel M, Rees W (1996) Our ecological footprint: reducing human impact on the earth. New Society, Gabriola Island
Yaukey P (1996) Patterns of avian population density, habitat use, and flocking behavior in urban and rural habitats during winter. Prof Geogr 48(1):70–81
Acknowledgments
We thank L. K. Fischer and two anonymous reviewers for their comments on a previous draft of the manuscript. Funding was provided by Deutsche Forschungsgemeinschaft, Research Training Group 780/3 Urban Ecology.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Meffert, P.J., Dziock, F. The influence of urbanisation on diversity and trait composition of birds. Landscape Ecol 28, 943–957 (2013). https://doi.org/10.1007/s10980-013-9867-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-013-9867-z