
Abstract
Protected areas are established to conserve biodiversity and facilitate resilience to threatening processes. Yet protected areas are not isolated environmental compounds. Many threats breach their borders, including transportation infrastructure. Despite an abundance of roads in many protected areas, the impact of roads on biota within these protected areas is usually unaccounted for in threat mitigation efforts. As landscapes become further developed and the importance of protected areas increases, knowledge of how roads impact on the persistence of species at large scales and whether protected areas provide relief from this process is vital. We took a two-staged approach to analysing landscape-scale habitat use and road-kill impacts of the common wombat (Vombatus ursinus), a large, widely distributed herbivore, within New South Wales (NSW), Australia. Firstly, we modelled their state-wide distribution from atlas records and evaluated the relationship between habitat suitability and wombat road fatalities at that scale. Secondly, we used local-scale fatality data to derive an annual estimate of wombats killed within an optimal habitat area. We then combined these two approaches to derive a measure of total wombats killed on roads within the protected area network. Our results showed that common wombats have a broad distribution (290,981 km2), one quarter (24.9 %) of their distribution lies within protected areas, and the percentage of optimal habitat contained within protected areas is 35.6 %, far greater than the COP10 guidelines of 17 %. Problematically, optimal habitat within protected areas was not a barrier to the effects of road-kill, as we estimated that the total annual count of wombat road-kill in optimal habitat within protected areas could be as high as 13.6 % of the total NSW population. These findings suggest that although protected areas are important spatial refuges for biodiversity, greater effort should be made to evaluate how reserves confer resilience from the impacts of roads across geographic ranges.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Roads are strongly correlated with both economic growth and natural resource degradation (Wilkie et al. 2000), while their effects on biota can extend outwards from the road edge for hundreds of metres (Forman and Alexander 1998; Bissonette and Adair 2008). Road networks are expanding globally, pressing the need to assess the conservation implications of their impact on biodiversity and existing conservation efforts. To date, most research on road impacts has focussed on localised impacts over small spatial areas (Clevenger and Waltho 2000; Carr and Fahrig 2001; Ramp et al. 2005; Klöcker et al. 2006; Roger and Ramp 2009), but assessment over large geographic regions is critical because road impacts operate along a continuum of scales that includes biogeographic, landscape, and patch-level effects (Trombulak and Frissell 2000; Forman et al. 2003; Grilo et al. 2011). Some notable exceptions do exist where landscape-scale studies on the effects of roads on wildlife have been evaluated (see Kramer-Schadt et al. 2004; Hobday and Minstrell 2008; Eigenbrod et al. 2009 for details). Landscape-scale studies are important in highlighting fragilities at large scales (see van der Ree et al. 2011) that are not apparent because some localised populations appear to be subsisting in road-impacted environments (Roger et al. 2011). Dependencies among adjacent populations across landscapes can destabilise metapopulations when one subpopulation becomes threatened, ultimately leading to a decline in overall species persistence (Gaston and Fuller 2008). In particular, species previously considered (or still considered) common, that have large geographic ranges or are able to disperse (seasonally or permanently), are frequently affected by breakdowns in exchange among populations at landscape scales (Epps et al. 2005).
Landscape-scale considerations of the impacts of roads on biodiversity is of direct relevance to global conservation efforts, for which the primary mechanism is the setting aside of protected areas (Regan et al. 2008). Due to the variety and severity of threats facing wildlife, protected areas are instrumental in conferring resilience to threatening processes (McDonnell et al. 2002). Protected areas are at the forefront of many regional and global conservation strategies, such as the tenth annual meeting of the Conference of the Parties (COP 10) on biosafety protocol. Protected areas are not impenetrable and many threatening processes breach their borders (Deguise and Kerr 2006). It is therefore crucial to be able to quantify how effective protected areas are protecting the species within them (Pressey et al. 2000; Crofts 2004; Wilson et al. 2007; Soutullo et al. 2008). The impact of roads in protected areas is often overlooked by conservation programmes (Ament et al. 2008), despite many protected areas having surprisingly high densities of roads within them that have been directly linked to population declines (e.g. Ramp and Ben-Ami 2006; Ament et al. 2008). Most protected areas fulfil the dual roles of protecting resource values as well as providing visitor enjoyment, but these roles are often difficult to balance as visitation can impact natural systems (Ament et al. 2008). Globally, road-kill remains a pervasive threat for large numbers of species both outside of and within protected areas (Clevenger et al. 2003; Fahrig and Rytwinski 2009).
The effect of roads on wildlife is often ignored because road fatalities have been considered unlikely to affect persistence of common species; which in most cases constitute the majority of road-kill (Forman and Alexander 1998). Lack of information on threats to common species is not new. Conservation investment routinely targets already threatened species and the areas where they are still found (McKinney and Lockwood 1999; Warren et al. 2001; Devictor et al. 2007), yet threatening processes also impact on common species (Gaston and Fuller 2008; Roger et al. 2011). Common species are defined as those species that are both abundant and widespread (Gaston and Fuller 2007). There is growing evidence that large numbers of species that currently meet these criteria are undergoing substantial decline (Gaston and Fuller 2007, 2008); however, responses of common species to land-use change remains largely unexplored (for exceptions see Epps et al. 2005). Given the functional role many common species have in facilitating ecosystem processes (Gaston 2008; Gaston and Fuller 2008), maintaining viable and functional populations of common species is a vital component of biodiversity conservation efforts (Lennon et al. 2004; Lyons et al. 2005; Pearman and Weber 2007).
Our objective was to assess road fatalities rates within protected areas for a common marsupial species, the common wombat (Vombatus ursinus); a typical example of a species that is impacted by roads at small scales (Roger et al. 2011) but for which the implication of road fatalities over large scales has not previously been examined. We focussed on assessing the impact of road fatalities in optimal habitat of protected areas due to the importance of these areas for species persistence. We addressed this landscape-scale question by applying a two-step approach. First we modelled their state-wide distribution from atlas records. We then used this information to evaluate the relationship between habitat suitability and annual wombat road fatalities across their geographic range. Secondly, we used local fatality data to derive an annual estimate of wombats killed within an optimal habitat area. We then combined these two approaches to derive an estimate of the annual total of wombats killed on roads within the protected area network.
Methods
Study species
The common wombat is a large burrowing marsupial and is thought to be both widespread and abundant throughout temperate south-eastern Australia (McIlroy 1995) (Fig. 1), however, informative data describing population distributions across its range is currently lacking (Roger et al. 2007). Despite this, their distribution appears to have contracted southwards since European settlement expansion circa 1860′s (McIlroy 1995; Buchan and Goldney 1998). Unlike many native species, common wombats benefit from the clearing of native bushland as it increases foraging habitat (Evans 2008). Their broad niche suggests they are a relatively robust and adaptable species, reflected by their use of agricultural and other modified landscapes (Roger et al. 2007; Roger and Ramp 2009). Adaptation to modified landscapes brings considerable cost, however, as they are frequently killed on roads because they exhibit little road avoidance or aversive behaviour (Roger and Ramp 2009).

Sighting locations of common wombats across their range throughout continental south-eastern Australia from 1990 to 2009. The boxed area adjacent to the ACT represents the location of the local scale fatality data. The abbreviations are those used for the states and territory of eastern Australia: ACT (Australian Capital Territory); NSW (New South Wales); VIC (Victoria); SA (South Australia); and QLD (Queensland)
Study area
Our analysis incorporated both broad-scale and fine-scale analyses. To define the geographic range of the common wombat we set the landscape extent of available habitat to be within New South Wales (NSW) and the Australian Capital Territory (ACT), an area that encompasses approximately half of the species’ total distribution (Fig. 1). Given the broad distribution of the species it was not possible to obtain similar quality information on habitat use across the species entire range. NSW is Australia’s most populous state and is located on the east coast of the continent with an area of 810,000 km2. The ACT is an enclave within NSW with a total land area of 2,400 km2. There are 752 protected areas that are greater than 10 km2 within the NSW and ACT, with a total area of 86,164 km2 for all protected areas. For the purpose of analysis we treated both the ACT and NSW as one modelling domain. To obtain an estimate of the annual total of wombats killed within optimal habitat inside protected areas we used previously published information from a 26-km segment of the Snowy Mountains Highway in Kosciuszko National Park (35º19′S, 148°14′E) (Roger and Ramp 2009).
Modelling common wombat distribution
We used common wombat presence data from records held within the NSW Atlas dataset obtained from the NSW Department of Environment Climate Change and Water (DECCW 2009). Records included data from multiple mammal surveys, collected between 1990 and 2009 by government staff, researchers, naturalists, environmental consultants, land management officers, and the public. To minimise spatial errors, we excluded all records before 1990 and where spatial uncertainty was greater than 500 m. Species occurrence data has been routinely used in species habitat modelling (Guisan and Thuiller 2005, Robertson et al. 2010) as it provides an estimate of species’ distributions across large scales where data are often scarce. We avoided the use of randomly selected pseudo-absence points (see Zarnetske et al. 2007) by locating wombat pseudo-absences from non-wombat sightings within the atlas database. We did this by examining the survey methods used to detect wombats and generated pseudo-absences where terrestrial mammals other than wombats were recorded using survey methods expected to identify the presence of wombats. Although these locations remain pseudo-absences, their selection has advantages over randomly selected points as they are derived from the same dataset as the presence data. To minimise type II errors, we excluded pseudo-absences within 320 m (equivalent to average wombat home range) of a known presence (Roger et al. 2007).
Predictive variables
We collated landscape-scale environmental and climatic variables for the study area using previously published studies of habitat selection by common wombats to guide variable selection (Skerratt et al. 2004; Roger et al. 2007; Evans 2008). Variables included descriptors of geography, vegetation, and climate (See online Appendix A). A Digital Elevation Model (DEM) was obtained from the Shuttle Radar Topography Mission (SRTM) with a spatial resolution of 3 arc seconds or approximately 90 m (Farr et al. 2007). Slope and aspect were derived from the DEM using ArcGIS 9.2 (ESRI 2007). Indices for topographic wetness (an estimate of the accumulation of overland water flow across catchments), slope steepness (Moore et al. 1991), and roughness (Allmaras et al. 1966) were generated to describe the surface properties of the DEM. We used the Enhanced Vegetation Index (EVI) satellite data obtained from the Terra Moderate Resolution Imaging Spectroradiometer (MODIS) sensor (Justice et al. 1998). Both EVI mean and variance were calculated for a 10-year period (2000–2009) at a resolution of 250 m. Digital information for water bodies (floodplains, lakes, reservoirs, and lagoons) was obtained from DECCW, derived from a combination of classification of spectral classes of Landsat MSS and TM imagery, along with ancillary wetland information (Kingsford et al. 2004). A digital image of major rivers was also obtained from DECCW, allowing for distance to water bodies and main rivers to be calculated as predictive variables. Climatic variables across the study area were obtained using the correlative modelling tool BIOCLIM 5.1 (Nix 1986). Twenty-seven climatic parameters were interpolated from recorded climatic data and elevation (Nix 1986; Houlder et al. 2000) (See online Appendix A).
Model development
We avoided collinear variables in any given model by reducing the number of variables prior to the selection of a final model. As we did not wish to subjectively reduce variables, we followed a data-driven pathway to reduce variables prior to model selection (Pinheiro and Bates 2000; Hastie et al. 2001; Thomson et al. 2010). We initially grouped the predictive variables into six collinear categories (geographic, vegetation, temperature, precipitation, moisture, and water). Using a logistic Generalized Additive Model (GAM) within the R statistical environment (package ‘gam’, R Development Core Team, 2005), we examined the goodness-of-fit values for each variable using the pseudo R 2. Following this, we selected a single representative variable from each of the six collinear groups to be used in the model selection process.
We carried out a further model selection process using all 64 unique combinations obtained from the six identified predictor variables (Table 1). To validate the models we ran a bootstrapping procedure using the .632 estimator rule (Hastie et al. 2001), which is suitable when distributions are unknown, and can outperform cross-validation (Efron 1983; Efron and Tibshirani 1997). This approach provides a predictive performance estimate of a model without the expense of collecting a completely new model testing set (Wintle et al. 2005).
We evaluated model performance by calculating the average area under the receiver operating curve (AUC) across all bootstrapped replicates and used this to evaluate the extent to which each model successfully estimated positive and negative observations (Fielding and Bell 1997; Hirzel et al. 2006). A best model set was selected by identifying all models with an AUC value within one standard error from the model with the highest AUC value. The one standard error rule is often used to find a more parsimonious model than the top model selected in the model selection process (Hastie et al. 2001). Selection of a final model from the best model set was made using a trade-off between models in the best model set that had the fewest numbers of predictor variables and the largest AUC value. Model selection was repeated using Akaike’s Information Criterion (AIC) to cross-check the model selection process. Hierarchical Partitioning was used to calculate the independent contribution of each variable across all model combinations (Mac Nally 2000). Fitted values of wombat habitat suitability were then predicted across the entire study area at a resolution of 90 m.
The distribution of wombat habitat suitability values was then used to obtain an estimate of the common wombat distribution across the study area. A threshold for occupancy was identified by applying the Jenks’ natural breaks method, which determines the best arrangement of values into classes by iteratively comparing sums of the squared difference between observed values within each class and class means (Brewer and Pickle 2002). The geographic range of the wombat within the study area was subsequently defined by suitability values above 0.16.
Linking suitability to road fatalities
To assess fatality rates of common wombats on roads throughout their geographic range we estimated the distribution of wombat fatalities on roads in NSW and the ACT. We obtained information on the distribution of roads within NSW and the ACT from DECCW (See online Appendix A). The road layer contained 2,632 segments of road throughout NSW, where segments were defined as sections of road between intersections. There were five categories of roads included in the road layer we used for our analysis: dual carriageway, principal road, secondary road, minor road and track, however, we excluded tracks from the analysis. The total length of road included in the analysis was 49,254 km. We grouped dual carriageways and principal roads into ‘highways’, secondary roads we labelled ‘major roads’ and minor roads were ‘minor roads’. All roads used in the analysis were sealed. To identify collision locations we used the Traffic Accident Database System of NSW (TADS), a database that includes statistics on road traffic accidents in NSW (See online Appendix A). Collision data between wildlife and vehicles are only included in TADS when reported to NSW Police because of human injury or extensive vehicle damage (Ramp and Roger 2008). There are very few data detailing the frequency of wombat vehicle collisions and the number of associated fatalities across their range, and although TADS considerably underestimates wombat fatalities (most collisions only result in injury to the animal and therefore go unreported), no other state-wide data exist. There were 150 wombat-related accidents recorded in the TADS database for the ten year period between 1996 and 2005. To provide context for this underestimate, Roger and Ramp (2009) reported 209 wombat road fatalities between the period of 1998 and 2005 from a single 40 km stretch of road.
Our approach therefore was to utilise information from TADS to infer the spatial distribution of collision likelihood, rather than using it to infer actual numbers killed annually within the study region. To estimate wombat road fatalities per kilometre of road within NSW we sampled all wombat fatality records contained in the TADS database using ArcGIS. Road segments with no reported collisions were assigned zero. To account for variability in road use (we did not have access to traffic volume data for all roads), the ratio between wombat-related vehicle collisions and all other wildlife-related vehicle collisions recorded in TADS was calculated and standardised by length of road segment.
To assess the relationship between habitat suitability and the mean probability of a wombat fatality, suitability values were averaged for each road segment and weighted by the length of the segment using Hawth’s Analysis Tools add-on for ArcGIS (Beyer 2004). We used the Jenks’ natural breaks classification to stratify wombat suitability probabilities into four categories: unsuitable (≤0.16), medium (≤0.45), high (≤0.78), and optimal (≤1). This enabled us to compare probabilities of wombat fatalities from the TADS database within habitat suitability categories across different road categories. We examined the relationship between habitat suitability groups and the mean probability of a wombat fatality using SPSS (SPSS Inc., 2006). Differences between habitat suitability groups in relation to road class were examined using two-way analysis of variance (ANOVA). Significant differences between means were compared using Tukey’s Least Significant Difference (LSD) methods.
Fatality rates within protected areas
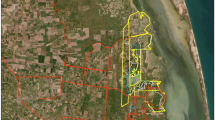
To derive a measure of annual wombat road fatalities per kilometre of road within optimal habitat in protected areas we used an additional source of road fatality data (see Roger and Ramp 2009). Fine-scale fatality information across the entire study area and for different levels of habitat suitability, within and outside protected areas, would be optimal but these data do not currently exist. For the purpose of this study, however, identifying the susceptibility of wombats to fatalities within optimal habitat in protected areas is sufficient. Fatalities of common wombats were recorded on a 26-km segment of the Snowy Mountains Highway within Kosciuszko National Park over a five year period (Fig. 2). Wombat fatalities were recorded using a hand-held global positioning system (GPS) device on average three days per week between 2002 and 2006. Carcases were removed from the roadside after recording to avoid double counting. The road segment was travelled 560 times, recording 117 wombat fatalities. Assuming wombat fatality frequencies were temporally correlated, we calculated monthly frequencies by dividing the recorded number of wombat fatalities with the number of trips each month. The actual number of fatalities per month was estimated by multiplying the monthly ratios with the number of days each month and averaged over the five year period. This resulted in an average fatality rate per month, which we summed to obtain the total rate per year. We then standardised this rate for each kilometre of road by dividing the total rate per year by the total number of kilometres driven each trip.

Fatality data was collected from the Snowy Mountains Highway in southern NSW. The sampled road segment as well as protected area boundaries are displayed
We then calculated the total kilometres of road length within protected areas that fell within the optimal suitability category using ArcGIS. Since our fine-scale wombat fatality data came from a road within an optimal habitat area (≥0.78), we could only reliably estimate wombat fatalities for optimal suitability areas within NSW in protected areas. As a final measure of range-wide road fatality impacts we multiplied the rate of wombats killed per year from the Snowy Mountains Highway by the total length of roads in protected areas that fall within optimal wombat habitat. We recognise that we had to make several assumptions in order to derive this calculation, and as a result the calculation only serves to provide a rough estimate of the numbers of wombat killed in optimal habitat protected areas. We assumed non-stationarity in the relationship between road presence and road-kill, while also assuming equal distribution of wombats across optimal habitat areas.
Results
Wombat habitat suitability model
There was good agreement on the final model among the two methods of model selection: the top AIC model was within 1 SE of the top AUC model. To maximise parsimony we chose the top AUC model which selected mean EVI, mean annual temperature, and mean moisture index of the warmest quarter. The final model explained 70.6 % of the deviance (AUC 0.802) (Table 2).
Mean annual temperature was negatively correlated with wombat habitat suitability, with suitability linearly declining in the warmer regions of north-eastern NSW (Fig. 3). Suitability declined steeply after a mean moisture of 0.4 was reached (mean moisture is scaled from 0 to 1). Suitability was significantly, but weakly, associated with mean EVI (Fig. 3). The inclusion of EVI, a measure of greenness (similar to the normalised vegetation index used in Roger et al. (2007)), indicated that although wombats make use of agricultural land for grazing, their distribution is constrained to wooded areas and/or cleared areas in proximity to remnant vegetation.

The partial residual plot shows the relationship between a given independent variable and the response variable given that other independent variables are also in the model. The x-axis represents the range of values for each environmental variable, a mean annual temperature (°C), b mean moisture index of warmest quarter ((1 − e soilb*store/maxstore)/(1 − e soilb), and c mean EVI. The y-axis displays the smoothed environmental variable
The habitat suitability model identified areas of optimal habitat mostly within the mountainous regions of the Great Dividing Range and in some coastal temperate regions (Fig. 4). The common wombat distribution appears to be bounded by a large climatic envelope that limits them to the mesic and semi-arid environments of south-eastern Australia, concurring well with expert opinion on common wombat distribution (Triggs 1988).

Habitat suitability values (probabilities) across NSW and ACT. Major protected areas networks within NSW and the ACT are also shown
Linking suitability to road fatality
Habitat suitability was positively correlated with fatality likelihood (Fig. 5, F 0.030, 2.448 = 10.453, P < 0.001). Results of Tukey’s Least Significant Test revealed large differences between the lowest suitability grouping (≤0.16) and the highest (≤1) as expected. The probability of a wombat fatality also varied among road categories and suitability groupings (Fig. 6). Significant differences between habitat suitability groups in relation to road class were observed (F 0.052, 2.426 = 18.515, P < 0.001). Significant variation once again occurred between the lowest suitability grouping and the highest.

Mean probability and standard error of a wombat fatality within protected areas plotted against stratified suitability groupings

Mean probability and standard error of a wombat fatality within protected areas plotted against road category and suitability groupings. Highways were omitted from the optimal suitability grouping due to their absence in protected areas
Distribution and fatality rates in protected areas
Common wombats were predicted to have a geographic range of 290,981 km2 (areas with habitat suitability above 0.16), distributed throughout eastern NSW and the ACT (Fig. 4) and for which 24.9 % is currently protected as national park or conservation reserve. The component of the total range considered optimal habitat (above 0.78) was calculated as 44,035 km2, 35.6 % of which is contained within protected areas.
Using the fine-scale information from the Snowy Mountains Highway we estimated that an average of 8.9 wombats were killed each month (with an annual average of 92.3), equating to 3.53 wombats per km of road per year. Given that there are 804 km of similar roads in optimal habitat within protected areas in the study area, we estimated that a total of 2,841 wombats may be being killed annually in these areas. Previous research in the same optimal habitat area has estimated a density of 1.3 wombats per km2 (Roger et al. 2007). Extrapolating this value by the total area of optimal habitat in protected areas (15,676 km2) equates to a population of 20,901 wombats within optimal habitat protected areas. Based on these figures, it is plausible that the total number of wombats killed annually within optimal habitat in protected areas is around 13.6 % of the total population.
Discussion
Empirical examples are needed to support theories developed primarily via simulation (e.g. Roger et al. 2011). Research has focussed on developing models of wildlife fatality hotspots (Ramp et al. 2005; Roger and Ramp 2009), the efficacy of mitigation (Clevenger and Waltho 2005), barrier effects on genetic drift and population viability (Gerlach and Musolf 2000), landscape planning (Jaarsma and Willems 2002), and the effects of road type on population persistence (Jaeger et al. 2005). However, this research is limited in scope, and cannot legitimately comment on how road development impacts on biota over larger spatial scales. Our research is one of the first to begin to quantify landscape extent impacts of roads over this large scale, but some notable exceptions do exist (see Hobday and Minstrell 2008; Fahrig and Rytwinski 2009 for details). Thus, although considerable uncertainty exists (due primarily to data limitations), we believe our two-step approach provides an important basis to begin to quantify how road fatalities impact on biodiversity. Roads will likely increase in significance as a form of disturbance over the coming decades making it all the more crucial.
The wide geographic range (211,107 km2) of common wombats across a range of elevations throughout eastern NSW confirms previous studies describing common wombat extent (Buchan and Goldney 1998; Catling et al. 2000; Roger et al. 2007; Borchard et al. 2008; Matthews et al. 2010). However, contrary to the ecological/biological mechanisms that have been proposed as good predictors of wombat distribution at local scales of analysis (Catling et al. 2000, 2002; Roger et al. 2007), regulation of wombat distributions across their geographic range is most strongly correlated with climatic controls (Guisan and Thuiller 2005). The selection of mean annual temperature suggests that across the species’ geographic range it is not extreme temperatures but mean temperatures that drive its distribution. Common wombats are also influenced by vegetation and the inclusion of mean EVI reflects wombat preference for good foraging habitat near cover (Evans 2008). McIlroy (1973) and Buchan and Goldney (1998) considered forest cover important for providing protection from predators and weather conditions. Unfortunately for common wombats, many roadside environments present these attributes by offering cleared land for grazing in close proximity to wooded habitat (Roger et al. 2007), excacerbating the problem of fatalities by attracting wombats to these locations. Given that the geographic range of common wombats has contracted southwards since European settlement (McIlroy 1973), it would be interesting to explore if this southern contraction is a result of changing climatic conditions, human changes in land-use, introduced threats, or a combination of all three.
In this study we assessed the relative abundance of common wombats within protected areas across the study area as well as the percentage of optimal habitat contained within the protected areas network, estimated using a habitat suitability model. We found that one quarter (24.9 %) of common wombat estimated geographic range lies within protected areas, while the percentage of optimal habitat represented within the protected areas network was 35.6 %. Our results suggest that protected areas constitute an important spatial refuge for common wombats and at first glance this seems to bode well for the continued persistence of the species.
Unsurprisingly, we also showed that the probability of a wombat road fatality increases with increasing habitat suitability (Fig. 5). This finding makes sense given that suitable habitat is correlated with higher densities of species, and this in turn can result in increased road fatality rates if animal density is linearly correlated with fatality likelihood (Forman and Alexander 1998). Nevertheless, it was important to demonstrate the link between habitat suitability and the probability of wombat road fatalities which to our knowledge has not been previously demonstrated. In related work, Grilo et al. (2011) observed a higher frequency of road fatalities on roads traversing continuously forested habitat. The authors highlighted that road networks in well-connected landscapes appear to be a serious threat to long-term population stability and viability. Although not specific to protected areas, their finding provides further evidence that road fatalities in areas considered important for species conservation are of concern for a wide range of species.
The relationship between road category and suitability grouping allowed us to demonstrate that the probability of a wombat fatality within highly suitable habitat remains high despite road category (Fig. 6). This is important for management which may not have considered major and minor roads as significant locales of wombat fatalities. The relationship between road category and road fatality is not linear, with various hypotheses presented to predict the effects of traffic on road-kill probability (see Seiler 2004; Jaeger et al. 2005 for details). How important road category is in terms of contributing to the frequency of road fatality seems highly dependent on species, with road avoidance behaviour likely playing a large role in determining susceptibility (Jaeger et al. 2005). By broadening the scope of study, research can begin to quantify landscape extent impacts of roads on populations and how patterns of habitat use and selection change with road-based fatality rates. It is vital that we develop an understanding of the motivations behind animal presence and movement to fully comprehend how roads interact with susceptible species. If species are highly susceptible to the impacts of roads then both rare and abundant species are potentially at great threat especially if their reproductive rates or recruitment rates are low.
A common assumption of protected area networks is that they act as sources for species across their geographic ranges, particularly if they constitute substantial components of the remaining or better quality habitat (Gaston 2008). We found strong support for this assumption with wombats favouring protected areas, but the number of fatalities occurring within these areas is problematic. Indeed, we previously reported that annual road fatalities within a 30 km2 protected area appeared to match the total population estimate for this area (Roger et al. 2007), a finding that implies that dispersal to this location was the only explanation for their continued existence there. This raises the question of whether protected areas that are infiltrated by roads may themselves contain localised population sinks, and effort should be expended in evaluating how protected areas confer resilience from the impacts of roads. Unfortunately, to test this theory for common wombats we currently lack information on how many are killed outside protected areas. We cannot assume that the relationship between density and fatality rates is linear, and hence a comparison of fatality rates for different habitat suitability and population densities across their geographic range would be a valuable contribution to the research. Likewise information on traffic volume (which we are lacking) has been shown to be important in assessing road impacts (Seiler 2004; Jaeger et al. 2005).
In a review of the ecological effects of roads, Forman and Alexander (1998) considered road fatalities unlikely to affect persistence of common species because birth rates were presumed to exceed road fatality rates for many species. As a result, species level conservation in road-impacted environments has remained focused on species already threatened with regional extinction in the near future (Forman et al. 2003). However, like the common wombat, a number of studies have recently documented population level depletions of common species as a result of road impacts at local scales (Jones 2000; Ramp and Ben-Ami 2006; Fahrig and Rytwinski 2009; Roger et al. 2011). There is a pressing need to quantify how different forms of land-use impact on biodiversity and how ultimately common species will persist as processes that underpin their decline intensify. How the threat of roads within protected areas impacts on species persistence should be of vital interest to conservation practitioners around the world.
References
Allmaras RR, Burwell RE, Larson WE, Holt RF (1966) Total porosity and random roughness of the interrow zone as influenced by tillage. Conservation research report, USDA, pp 1–14
Ament R, Clevenger AP, Yu O, Hardy A (2008) An assessment of road impacts on wildlife populations in US National Parks. Environ Manag 42:480–496
Beyer HL (2004) Hawth's Analysis Tools for ArcGIS. Available from http://www.spatialecology.com/htools
Bissonette JA, Adair W (2008) Restoring habitat permeability to roaded landscapes with isometrically-scaled wildlife crossings. Biol Conserv 141:482–488
Borchard P, McIlroy J, McArthur C (2008) Links between riparian characteristics and the abundance of common wombat (Vombatus ursinus) burrows in an agricultural landscape. Wildl Res 35:760–767
Brewer CA, Pickle L (2002) Evaluation of methods for classifying epidemiological data on choropleth maps in series. Ann Assoc Am Geogr 92:662–681
Buchan A, Goldney DC (1998) The common wombat Vombatus ursinus in a fragmented landscape. In: Wells RT, Pridmore PA (eds) Wombats. Surrey Beatty & Sons, Chipping Norton, New South Wales, pp 251–261
Carr LW, Fahrig L (2001) Effect of road traffic on two amphibian species of differing vagility. Conserv Biol 15:1071–1078
Catling PC, Burt RJ, Forrester RI (2000) Models of the distribution and abundance of ground-dwelling mammals in the eucalypt forests of north-eastern New South Wales in relation to habitat variables. Wildl Res 27:639–654
Catling PC, Burt RJ, Forrester RI (2002) Models of the distribution and abundance of ground-dwelling mammals in the eucalypt forests of north-eastern New South Wales in relation to environmental variables. Wildl Res 29:313–322
Clevenger AP, Waltho N (2000) Factors influencing the effectiveness of wildlife underpasses in Banff National Park, Alberta, Canada. Conserv Biol 14:47–56
Clevenger AP, Waltho N (2005) Performance indices to identify attributes of highway crossing structures facilitating movement of large mammals. Biol Conserv 121:453–464
Clevenger AP, Chruszcz B, Gunson KE (2003) Spatial patterns and factors influencing small vertebrate fauna road-kill aggregations. Biol Conserv 109:15–26
Crofts R (2004) Linking protected areas to the wider world: a review of approaches. J Environ Pol Plann 6:143–156
DECCW (2009) The atlas of NSW wildlife. The NSW Department of Environment, Climate Change and Water, Sydney, Australia
Deguise IE, Kerr JT (2006) Protected areas and prospects for endangered species conservation in Canada. Conserv Biol 20(1):48–55
Devictor V, Godet L, Julliard R, Couvet D, Jiguet F (2007) Can common species benefit from protected areas? Biol Conserv 139:29–36
Efron B (1983) Estimating the error rate of the prediction rule: improvement on cross-validation. J Am Statistical Association 78:316–331
Efron B, Tibshirani R (1997) Improvements on cross-validation: the.632+ bootstrap method. J Am Statistical Association 92:548–560
Eigenbrod F, Hecnar SJ, Fahrig L (2009) Quantifying the road-effect zone: threshold effects of a motorway on anuran populations in Ontario. Canada. Ecol Soc 14:24
Epps CW, Palsboll PJ, Wehausen JD, Roderick GK, Ramey II RR, McCullough DR (2005) Highways block gene flow and cause a rapid decline in genetic diversity of desert bighorn sheep. Ecol Lett 8:1029–1038
ESRI (2007) ArcGIS version 9.2. Environmental Systems Research Institute, Redlands, California, USA
Evans MC (2008) Home range, burrow-use and activity patterns in common wombats (Vombatus ursinus). Wildl Res 35:455–462
Fahrig L, Rytwinski T (2009) Effects of roads on animal abundance: an empirical review and synthesis. Ecol Soc 14:21
Farr TG, Rosen PA, Caro E, Crippen R, Duren R, Hensley S, Kobrick M, Paller M, Rodriguez E, Roth L, Seal D, Shaffer S, Shimada J, Umland J, Werner M, Oskin M, Burbank D, Alsdorf D (2007) The shuttle radar topography mission. Rev Geophys 45:33
Fielding AH, Bell JF (1997) A review of methods of the assessment f prediction errors in conservation presence/absence models. Environ Conserv 24:38–49
Forman RTT, Alexander LE (1998) Roads and their major ecological effects. Annu Rev Ecol Syst 29:207–231
Forman RTT, Sperling D, Bissonette JA et al (2003) Road ecology: science and solutions. Island Press, Washington
Gaston KJ (2008) Bliodiversity and extinction: the importance of being common. Prog Phys Geog 32:73–79
Gaston KJ, Fuller RA (2007) Biodiversity and extinction: losing the common and the widespread. Prog Phys Geog 31:213–225
Gaston KJ, Fuller RA (2008) Commonness, population depletion and conservation biology. Trends Ecol Evol 23:14–19
Gerlach G, Musolf K (2000) Fragmentation of landscape as a cause for genetic subdivision in bank voles. Conserv Biol 14:1066–1074
Grilo C, Ascensao F, Bissonette JA, Santos-Reis M (2011) Do well-connected landscapes promote road-related mortality? Eur J Wildl Res 57:707–716
Guisan A, Thuiller W (2005) Predicting species distribution: offering more than simple habitat models. Ecol Lett 8:993–1009
Hastie T, Tibshirani R, Friedman JH (2001) The elements of statistical learning: data mining inference and prediction. Springer, New York
Hirzel AH, Le Lay G, Helfer V, Randin C, Guisan A (2006) Evaluating the ability of habitat suitability models to predict species presences. Ecol Model 199:142–152
Hobday AJ, Minstrell ML (2008) Distribution and abundance of roadkill on Tasmanian highways: human management options. Wildl Res 35:712–726
Houlder DJ, Hutchinson MF, Nix HA, McMahon JP (2000) ANUCLIM User Guide, Version 5.1. Centre for Resource and Environmental Studies, Australian National University, Canberra
Jaarsma CF, Willems GPA (2002) Reducing habitat fragmentation by minor rural roads through traffic calming. Landsc Urban Plan 58:125–135
Jaeger JAG, Bowman J, Brennan J, Fahrig L, Bert D, Bouchard J, Charbonneau N, Frank K, Gruber B, von Toschanowitz KT (2005) Predicting when animal populations are at risk from roads: an interactive model of road avoidance behavior. Ecol Model 185:329–348
Jones ME (2000) Road upgrade, road mortality and remedial measures: impacts on a population of eastern quolls and Tasmanian devils. Wildl Res 27:289–296
Justice CO, Vermote E, Townshend JRG, Defries R, Roy DP, Hall DK, Salomonson VV, Privette JL, Riggs G, Strahler A, Lucht W, Myneni RB, Knyazikhin Y, Running SW, Nemani RR, Wan ZM, Huete AR, Van Leeuwen W, Wolfe RE, Giglio L, Muller JP, Lewis P, Barnsley MJ (1998) The Moderate Resolution Imaging Spectroradiometer (MODIS): land remote sensing for global change research. Ieee T Geosci Remote 36:1228–1249
Kingsford RT, Brandis K, Thomas RF, Crighton P, Knowles E, Gale E (2004) Classifying landform at broad spatial scales: the distribution and conservation of wetlands in New South Wales, Australia. Mar Freshw Res 55:17–31
Klöcker U, Croft DB, Ramp D (2006) Frequency and causes of kangaroo-vehicle collisions on an Australian outback highway. Wildl Res 33:5–15
Kramer-Schadt S, Revilla E, Wiegand T, Breitenmoser U (2004) Fragmented landscapes, road mortality and patch connectivity: modelling influences on the dispersal of Eurasian lynx. J Appl Ecol 41:711–723
Lennon JJ, Koleff P, Greenwood JJD, Gaston KJ (2004) Contribution of rarity and commonness to patterns of species richness. Ecol Lett 7:81–87
Lyons KG, Brigham CA, Traut BH et al (2005) Rare species and ecosystem functioning. Conserv Biol 19:1019–1024
Mac Nally R (2000) Regression and model-building in conservation biology, biogeography and ecology: the distinction between and reconciliation of ‘predictive’ and ‘explanatory’ models. Biodiversity Conserv 9:655–671
Matthews A, Spooner PG, Lunney D, Green K, Klomp NI (2010) The influence of snow cover, vegetation and topography on the upper range limit of common wombats (Vombatus ursinus) in the subalpine zone, Australia. Divers Distrib 16:277–287
McDonnell MD, Possingham HP, Ball IR, Cousins EA (2002) Mathematical methods for spatially cohesive reserve design. Environ Model Assess 7:107–114
McIlroy JC (1973) Aspects of the ecology of the Common wombat Vombatus ursinus (Shaw, 1800). Dissertation, Australian National University
McIlroy JC (1995) Common Wombat. In: Strahan R (ed) The mammals of Australia. Reed Books, Chatswood, New South Wales, pp 204–205
McKinney ML, Lockwood JL (1999) Biotic homogenization: a few winners replacing many losers in the next mass extinction. Trends Ecol Evol 14:450–453
Moore ID, Grayson RB, Ladson AR (1991) Digital terrain modeling—a review of hydrological, geomorphological, and biological applications. Hydrol Process 5:3–30
Nix H (1986) A biogeographic analysis of Australian Elapid snakes. In: Longmore R (ed) Snakes: Atlas of elapid snakes of Australia. Bureau of Flora and Fauna, Canberra, pp 4–10
Pearman PB, Weber D (2007) Common species determine richness patterns in biodiversity indicator taxa. Biol Conserv 138:109–119
Pinheiro JC, Bates DM (2000) Mixed effects models in S and S-PLUS. Springer, New York
Pressey RL, Hager TC, Ryan KM, Schwarz J, Wall S, Ferrier S, Creaser PM (2000) Using abiotic data for conservation assessments over extensive regions: quantitative methods applied across New South Wales, Australia. Biol Conserv 96:55–82
R Development Core Team (2005) R: a language and environment for statistical computing. Vienna, Austria
Ramp D, Ben-Ami D (2006) The effect of road-based fatalities on the viability of a peri-urban swamp wallaby population. J Wildl Manag 70:1615–1624
Ramp D, Roger E (2008) Frequency of animal-vehicle collisions in NSW. In: Lunney D, Munn A, Meikle W (eds) Too close for comfort. Royal Zoological Society of New South Wales, Mosman, Australia, pp 118–126
Ramp D, Caldwell J, Edwards KA, Warton D, Croft DB (2005) Modelling of wildlife fatality hotspots along the snowy mountain highway in New South Wales, Australia. Biol Conserv 126:474–490
Regan HM, Hierl LA, Franklin J, Deutschman DH, Schmalbach HL, Winchell CS, Johnson BS (2008) Species prioritization for monitoring and management in regional multiple species conservation plans. Divers Distrib 14:462–471
Robertson MP, Cumming GS, Erasmus BFN (2010) Getting the most out of atlas data. Divers Distrib 16:363–375
Roger E, Ramp D (2009) Incorporating habitat use in models of fauna fatalities on roads. Divers Distrib 15:222–231
Roger E, Laffan SW, Ramp D (2007) Habitat selection by the common wombat (Vombatus ursinus) in disturbed environments: implications for the conservation of a ‘common’ species. Biol Conserv 137:437–449
Roger E, Laffan SW, Ramp D (2011) Road impacts a tipping point for wildlife populations in threatened landscapes. Popul Ecol 53:215–227
Seiler A (2004) Trends and spatial patterns in ungulate-vehicle collision in Sweden. Wildlife Biol 10:301–313
Skerratt LF, Skerratt JHL, Banks S, Martin R, Handasyde K (2004) Aspects of the ecology of common wombats (Vombatus ursinus) at high density on pastoral land in Victoria. Aust J Zool 52:303–330
Soutullo A, Castro MD, Urios V (2008) Linking political and scientifically derived targets for global biodiversity conservation: implications for the expansion of the global network of protected areas. Divers Distrib 14:604–613
Thomson FJ, Moles AT, Auld TD, Ramp D, Ren S, Kingsford RT (2010) Chasing the unknown: predicting seed dispersal mechanisms from plant traits. J Ecol 98:1310–1318
Triggs B (1988) The wombat: common wombats in Australia. Revised edition. New South Wales University Press, Kensington, New South Wales
Trombulak SC, Frissell CA (2000) Review of ecological effects of roads on terrestrial and aquatic communities. Conserv Biol 14:18–30
van der Ree R, Jaeger JAG, van der Grift EA, Clevenger AP (2011) Effects of roads and traffic on wildlife populations and landscape funtion: road ecology is moving toward larger scales. Ecol Soc 16:48
Warren MS, Hill JK, Thomas JA, Asher J, Fox R, Huntley B, Roy DB, Telfer MG, Jeffcoate S, Harding P, Jeffcoate G, Willis SG, Greatorex-Davies JN, Moss D, Thomas CD (2001) Rapid responses of British butterflies to opposing forces of climate and habitat change. Nature 414:65–69
Wilkie D, Shaw E, Rotberg F, Morelli G, Auzel P (2000) Roads, development, and conservation in the Congo basin. Conserv Biol 14:1614–1622
Wilson KA, Underwood EC, Shaw MR, Possingham HP (2007) Conserving biodiversity efficiently: what to do, where, and when. PLoS Biol 5:223
Wintle BA, Elith J, Potts JM (2005) Fauna habitat modelling and mapping: a review and case study in the Lower Hunter Central Coast region of NSW. Austral Ecol 30:729–748
Zarnetske PL, Edwards TC, Moisen GG (2007) Habitat classification modeling with incomplete data: pushing the habitat envelope. Ecol Appl 17:1714–1726
Acknowledgments
We are grateful to New South Wales Department of Environment, Climate Change and Water for the provision of Atlas data and the New South Wales Roads and Traffic Authority for provision of TADS. We thank David Warton, Shiquan Ren and David Eldridge for statistical advice and Evan Webster and Shawn Laffan for technical support.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Roger, E., Bino, G. & Ramp, D. Linking habitat suitability and road mortalities across geographic ranges. Landscape Ecol 27, 1167–1181 (2012). https://doi.org/10.1007/s10980-012-9769-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-012-9769-5