
Abstract
Assessment of locations where wildlife species cross highways is a key question in mitigating future wildlife-vehicle mortality. Examination of the spatial structure, complexities, and patterns of vegetation or other land-use types (i.e., cropland, urban areas) near roadways allows scientists to identify any thresholds that influence where animals are likely to die or successfully cross the roadway. We used a historic 1982 to 2017 dataset of ocelot (Leopardus pardalis pardalis) mortality locations and approximate road crossing locations of telemetered ocelots in the Lower Rio Grande Valley in Texas to examine the spatial structure of woody vegetation within a hypothesized road effect zone. We determined if there were differences in the spatial structure of woody cover within a 1050 m buffer of each successful crossing and roadkill location using PERMANOVA and principal component analyses. We used a similarity percentages analysis to determine the relative contribution of each aspect of spatial structure on differences in successful crossing and roadkill locations. We found statistically significant differences in spatial attributes of patches at the locations of successful crossing versus roadkill locations of ocelots at the 150 m spatial extent (pseudo-F1,41 = 4.85, P(perm) = 0.008, permutations = 9949). Largest patch index contributed most to the differences between successful crossing and roadkill locations (15.94%), followed by mean patch area (15.44%), percent woody cover (15.18%), aggregation indices (14.53%), Euclidean nearest neighbor (13.47%), edge (13.08%) and patch densities (12.36%). Roadkill locations were clustered in locations with lower-quality woody cover within 300 m of the highway. This suggests areas immediately surrounding roads need to contain woody patches that are larger and closer together to reduce the barrier-effects of roads. Such information is important for informing highway planners about where to encourage crossings or to build wildlife crossing structures to promote movement across the highway.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Continental transportation corridors (i.e., roads) are a profound fixture on the world landscape, accounting for > 35 million km as of 2020, and a projected > 70% increase over the next 30 years (Dulac 2013; Meijer et al. 2018). Roads have had a profound negative impact on ecosystems, ecological processes, and wildlife species globally (Jackson and Fahrig 2011; van der Ree et al. 2015). In advanced countries like the United States where roads already affect more than 20% of the landmass (Forman and Alexander 1998), expansions of road infrastructure, especially in areas with known human impacts, can further exacerbate deleterious impacts on flora and fauna (Ament et al. 2008; Taylor and Goldingay 2010; Qin et al. 2022). Near large metropolitan areas, a high human population growth rate often coincides with the rapid land conversion of adjacent wildlife habitat which leads to an increase in road networks in once-rural landscapes; this, in turn, leads to road planning and habitat preservation challenges (Forman et al. 2003; Shanley and Pyare 2011). Construction and expansion of new and existing roadways result in increased traffic volume leading to increased direct road effects such as a barrier to movement and reduced landscape connectivity (Grilo et al. 2021; Veals et al 2022b), increased vehicle-induced mortality (Blackburn et al. 2021b), and stress-induced physiological changes in species risk (Ditmer et al. 2018).
Ecological effects of roadways occur within a road effect zone (REZ), which is defined as the asymmetrical area around a road over which significant ecological impacts are measured (Bennett 1991, Forman and Deblinger 2000). The REZ is often used by transportation planners and biologists to estimate the area of impact for species and determine where to mitigate these effects (Andrasi et al. 2021). The effects of REZs may vary by habitat type, road type, species, and sex (Kasworm and Manley 1990; Semlitsch et al. 2007; Shanley and Pyare 2011; Andrasi et al. 2021). For instance, in the Netherlands, relative bird diversity was greatest at 1,000 m from roadways in pasturelands. however, density near roads in forested areas was greatest around 500 m from roadways (Forman and Deblinger 2000). In carnivores, the European wildcat (Felis sylvestris) showed decreased habitat use and avoidance in areas within 200 m of roadways (Klar et al. 2008). Blackburn et al. (2021a) assessed the scale of effect of ocelot (Leopardus pardalis) road mortality locations and random sites and found that land cover beyond 1050 m only had significant effects with patches that were closer together. Knowledge of the extent of the REZ and the spatial arrangement of land cover types within the REZ has powerful implications for understanding where species successfully traverse roads and where they succumb to road mortalities.
Wildlife vehicle collisions are the leading and most detrimental impact of roads on wildlife populations (Grilo et al. 2021). In some areas, vehicle-caused mortality is the leading source of mortality for wildlife, which can pose serious threats to endangered or threatened carnivores. For instance, Grilo et al. (2021) analyzed roadkill rates of 392 different mammal species worldwide and found that roadkill rates of brown hyena (Hyaena brunna) in southern Africa and maned wolf (Chrysocyon brachyurus) in Brazil are projected to lead to increased extinction risk. Road mortalities enhance the negative effects of decreased gene flow, limited connectivity, recruitment, and dispersal of wildlife species (van der Ree et al. 2015). Wildlife-vehicle mortalities have often been linked to areas near vegetative cover (Clevenger et al. 2003), spatial aggregations of habitat patches (Main and Allen 2002), and landscape structure of vegetative cover (Blackburn et al. 2021a). Understanding differences in spatial arrangements of locations where wildlife species successfully traverse roads compared to wildlife-vehicle observations is important to mitigate future mortality (Neumann et al. 2012) and help inform placement of wildlife crossing structures.
In the southern Texas-northern México borderlands, the Lower Rio Grande Valley eco-region contains a rapidly developing bi-national metropolitan area with an increasing urban footprint and transportation network infrastructure. The region’s extensive and expanding agricultural fields, which radiate from the urban core of Brownsville, McAllen, and Harlingen, produce the third-greatest citrus yields in the United States (Leslie 2016; Lombardi et al. 2020). The combination of these factors led to the loss and fragmentation of woody cover communities in the region in the 20th century, with 91% loss through the 1990s (Tremblay et al. 2005). More recent studies indicate while woody cover across the Lower Rio Grande Valley has increased since 1987, it was due to a sharp increase in small patches < 0.4 ha and loss of large patches > 100 ha (Lombardi et al. 2020). The loss of large patches of native woody vegetation, coupled with expanding road infrastructure has had a negative effect on wildlife. Among affected species is the US-federally endangered ocelot (Leopardus pardalis pardalis), a medium-sized native felid.
Ocelots in Texas are separated into two isolated populations; the Ranch Ocelot Population (likely 60 + individuals; Lombardi et al. 2022) occurs on large private ranchlands in Kenedy and Willacy counties. The smaller Refuge Ocelot Population (16–20 individuals) occurs in coastal areas of the Lower Rio Grande Valley, specifically in and around Laguna Atascosa National Wildlife Refuge (LANWR; S. Lehnen, USFWS, pers. com). Loss of available woody cover in the region (Veals et al. 2022a, b) coupled with extensive road infrastructure and traffic volumes has led to high levels of ocelot-road mortality to the point where vehicle collisions are the primary cause of ocelot mortality within this population (Haines et al. 2005; Blackburn et al. 2021b). Consequently, research on how ocelot populations in this region interface with low to high traffic paved roads is critical to future recovery and road mitigation efforts (Haines et al. 2005; Schmidt et al. 2020; Blackburn et al. 2021a, b).
Over the last 20 years, evidence has grown of the importance of incorporating landscape structure and landscape metrics in understanding of effects of rapid urbanization on woody cover (Lombardi et al. 2020), ocelot habitat use (Jackson et al. 2005; Lombardi et al. 2021), and road ecology in southern Texas (Blackburn et al 2021a, 2022; Schmidt et al. 2020; Veals et al. 2022a, b). Ocelots in large private ranchlands devoid of highways use large, low-density patches of live oak (Quercus virginiana) forest and thornshrub with lower shape indices and high aggregations (Lombardi et al. 2021). However, ocelots in the Refuge Population select for large, isolated patches of Tamaulipan thornshrub communities with higher landscape shape indices (Jackson et al. 2005) and are more likely to use areas farther from paved roads (Veals et al. 2022a, b). In an evaluation of road mortality data, Blackburn et al. (2021a) used landscape metrics to suggest ocelot road mortality sites had a more woody cover than random sites along highways. Schmidt et al. (2020) also indicated ocelot mortality risk was highest with smaller core areas (ha) and greater edge contrast (%) of woody scrub habitats. Historically, the placement of wildlife crossing structures for ocelots has focused on areas with ocelot road mortalities and proximate woody cover (Blackburn et al. 2022). However, no study has investigated the spatial patterns of woody cover within a hypothesized REZ differ between where ocelots successfully crossed and where individuals were roadkilled. If woody cover metrics differ significantly between crossing sites and roadkill locations, it would have powerful implications for the placement of wildlife crossing structures on paved roadways and habitat restoration efforts in nearby areas.
We analyzed the spatial structure of woody cover at monitored ocelot highway crossing locations from a 19-year (1982–2001) telemetry database (n = 52) and locations of confirmed roadkilled ocelots (n = 26) from 1982 to 2017. We compared the landscape structure of woody cover within a 1,050 m REZ of 1) individual telemetered ocelot successful crossing locations and road mortality locations of ocelots that died of vehicle collisions (all ocelot analysis); 2) successful ocelot crossing locations of telemetered ocelots that were roadkilled to their respective roadkill locations (paired ocelot analysis); 3) successful telemetered ocelot crossing locations for ocelots with an unknown fate compared to successful telemetered ocelot crossing locations for ocelots who were roadkilled between 1982 and 2017 (successful crossing analysis).
Methods
Study area
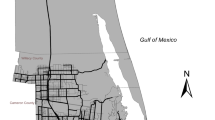
The study focused on a fragmented urban-agriculture-wildland mosaic landscape in coastal Cameron County in the Lower Rio Grande Valley and Laguna Madre Barrier Island and Coastal Marsh eco-regions in southern Texas (Griffith et al. 2007). This area has a semi-arid subtropical climate (10–36 °C) with highly variable rainfall (313–529 mm) and episodic drought periods (Norwine and Kuruvilla 2007). The focal area of interest contained a mosaic of low to medium-density urban development, large private ranchlands managed for cattle, native game species (e.g., white-tailed deer (Odocoileus virginiana), and waterfowl, row-crop agriculture, and protected natural habitat (Fig. 1).

Map of the urban-agricultural mosaic study areas including the location of Laguna Atascosa National Wildlife Refuge (LANWR), paved roads with low–high traffic volume, dirt and gravel roadways in eastern Willacy and Cameron counties in southern Texas
The focal area is in and around the US Fish and Wildlife Service’s (USFWS) Laguna Atascosa National Wildlife Refuge (LANWR; 26° 13’ N, 97°, 22’ W) and proximate areas in Cameron and Willacy Counties, Texas (Haines et al. 2005; Blackburn et al. 2021a; Lehnen et al. 2021; Veals et al 2022b). Paved roads included county roads (CR), state highways (SH), farm-to-market roads (FM), and major secondary roads on Laguna Atascosa National Wildlife Refuge (i.e., Bayside Loop [active 1982–2013]). Traffic volume in these areas has been consistent or increased since the end of ocelot monitoring in 2001 with annual average daily traffic (AADT) volumes ranging from 176 to > 9000 cars/day (Table 1). High traffic volume roads (> 7000 cars/day) are connected to seasonal tourism on the nearby South Padre Island beachside community (Aaron Economic Consulting 2017).
Laguna Atascosa National Wildlife Refuge is found in the northeastern half of the study area and is composed of low-traffic volume paved roadways (40 kph) that served as scenic drives. Vegetation communities on the refuge included patches of native thornshrub communities in the southern and northern edges of LANWR, scattered hypersaline estuarine lakes, tidal flats, and restored and native coastal prairie (Veals et al. 2022a, b). Private ranchlands in Willacy County and scattered throughout eastern Cameron County contained rangeland for cattle and fragmented patches of thornshrub communities. Associated thornshrub vegetation communities used by ocelots in this area included lime prickly ash (Zanthoxylum fagara), spiny hackberry (Celtis pallida), crucifixion thorn (Castela emoryi), blackbrush acacia (Vachellia rigidula), and ebony (Ebenopsis ebano; Lombardi et al. 2021).
Methodology
Ocelot telemetry data collection
We complied a historic (very high frequency [VHF]) telemetry database of ocelots (n = 52, 29 males, 23 females; Blackburn et al. 2021a; Veals et al. 2022a, b) monitored from individuals collared on LANWR and immediate surrounding areas, Cameron County, Texas from 1982 to 2001. Telemetry location data with VHF varied across time, but at least one diurnal location was recorded per day, and locations were recorded up to two weeks/per month using traditional ground-based triangulation with more than one observer sporadically throughout the year (Veals et al. 2022a, b). During these 19 years, duration of VHF monitoring generally ranged from ½ to 2 years, with multiple individuals monitored > 2 times over multiple years.
Ocelot road mortality data
We obtained 26 roadkill locations from 1982 to 2017 from a comprehensive database of confirmed ocelot-vehicle mortalities in the study area of both radio-collared and non-collared individuals (Blackburn et al. 2021a; Schmidt et al. 2020). Of these 26 roadkill locations found throughout the region, only five were of radio-collared individuals monitored from the 1982 to 2001 monitoring period. These road mortality observations were each discovered incidentally over the 35 years by state game wardens, academic and federal biologists, and private citizens; all mortalities were verified by trained biologists, and GPS locations and habitat descriptions were collected. Unlike more standardized road mortality surveys, the nonsystematic collection method of these ocelot mortality locations over 35 years may represent a potential bias (Blackburn et al. 2021a) and may not be representative of every ocelot road mortality during this period. Due to the small size of this local population (less than 120 individuals in a given year); however, we are confident that the data used in this study likely represent the majority of ocelot mortalities in this region.
Spatial analyses
To understand the role of the spatial structure of woody cover at crossing locations of paved roads we employed a four-pronged approach: classification of paved roadways in the study area, identification of successful road crossing locations of telemetered ocelots through simulated movement paths, roadkill locations on paved roads (Blackburn et al. 2021a), and development of temporally distinct remotely sensed imagery for each corresponding year(s) for successful crossing and roadkill locations (Table 2).
For the roadway data, we downloaded paved roadway shapefiles from the Texas Department of Transportation (2022) and cross-referenced satellite imagery. From 1982 to 2017, based on satellite imagery, we documented no new roadways constructed in the focal study area. The only large-scale road expansions and improvements of existing roads were initiated in the mid-2010s to install wildlife crossing structures on SH100 (starting in 2016) and FM106 (starting in 2013). In the larger Lower Rio Grande Valley region, however, large-scale road construction took place closer to major and secondary roads in and around the cities of Brownsville, Harlingen, San Benito, Los Fresnos, and Port Isabel, which were located outside (> 10 km) of known ocelot telemetry locations.
We classified roads as unpaved and paved based on imagery from the USFWS (2017) and satellite LANDSAT 5 imagery (30-m) and conducted ground assessments to verify the substrate of each roadway in March 2019 and June 2021. Unpaved roads, including dirt, earthen two-track, gravel, and caliche gravel surfaces were not used in this analysis because wildlife road mitigation projects (i.e., wildlife crossing structures) are largely tied to paved highways and roads and there has never been a documented ocelot road mortality on an unpaved road in Texas.
Following similar methodologies as Poessel et al. (2014), we simulated movement paths for individual ocelots by creating lines between consecutive observed locations using ArcMap (version 10.8.1 ESRI, Redlands, CA). We identified movement paths that intersected roads, thereby indicating a successful paved road crossing (Chruszcz et al. 2003; Lewis et al. 2011; Poessel et al. 2014). VHF telemetry data can be potentially biased in identifying the exact coordinates of locations, so there was a potential that some of the crossing events across paved roads were due to location error rather than actual crossings (Lewis et al. 2011); therefore, we refer to these as “apparent successful crossings.” Confirmed crossings were validated through a four-step process that began with the identification of telemetry locations on each side of a paved roadway. Three additional criteria were used to refine crossing classifications: (1) ≥ 1 sequential location was recorded on both sides of the paved road to reduce the likelihood that the crossing was simply a location error (Lewis et al. 2011); (2) locations on each side of the road were < 72 h apart to avoid the probability that the ocelot took a different route that did not include a road crossing; (3) a crossing location was the mean minimum daily distance traveled for ocelots calculated from the telemetry data (Blackburn et al. 2021a; Veals et al 2022a, b) from the endpoint (e.g., dead-end) of a paved road to ensure the individual crossed rather than travel around the road. We do acknowledge that due to the lower temporal frequency of VHF radio collars it is likely there were more road crossings than we estimated.
Landscape structure analyses
We obtained remotely sensed imagery for the years within successful ocelot crossings (1982–2001) and known ocelot-vehicle mortality locations (1982–2017) to examine the spatial structure, shape, and complexities of woody cover within a REZ around each successful crossing roadkill location on paved roads. We used previously classified LANDSAT 5 and LANDSAT 8 imagery created by Blackburn et al. (2021a) and analyzed new imagery for additional years to ensure that we accounted for potential changes in woody cover over time. We conducted an unsupervised classification in ERDAS IMAGINE 2018 (Hexagon Geospatial, Norcross, GA). We selected scenes that had minimal cloud cover (< 10%) during the cropland growing season (March–May) to distinguish among cover types. We followed a classification scheme developed by Blackburn et al. (2021a) for the region: woody cover [i.e., thornshrub communities, and taller tree species (i.e., brasil, ebony, and mesquite], herbaceous cover (i.e., native and invasive grassland and coastal prairie), bare ground (i.e., bare earth, oil pads, inland, and coastal dunes), water (rivers, coastal and inland playas, ox-bow lakes [sic resaca], estuarine areas, and the intercoastal waterway). Areas of row-crop agriculture and urban areas were manually digitized for each time period based on data acquired Texas Natural Resources Information System (TNRIS, Austin, TX) and we mosaicked these digitized layers into the classified imagery for temporal representative images for the corresponding year(s). We used 200 random points to conduct an accuracy assessment our classified imagery until we exceeded an accuracy threshold of > 85% (Lombardi et al. 2020; Blackburn et al. 2021a).
Based on road ecology literature indicating the importance of REZs within 1 km of highways (Clevenger and Huijser 2011), and the landscape structure of ocelot road mortality locations in Blackburn et al. (2021a), we developed a methodology to examine the role of woody spatial structure on ocelot crossing patterns. We quantified seven class-level landscape metrics within five spatial extents surrounding confirmed successful ocelot crossing locations and mortality sites to examine potential scales of road effects (Fig. 2). We used a subset of spatial extents used by Blackburn et al (2021a, 2022) to measure road effects (i.e., 150, 300, 450, 600, 1050 m). We selected 1050 m as the final buffer size because our previous research found that areas further away may be more likely to be affected by landscape-level processes than direct road effects (Clevenger and Huijser 2011; Veals et al. 2022a, b).

Example of location of identified ocelot M170 successful paved road crossing location and hypothesized road effect zone (150, 300, 450, 600, 1050 m) used around each paved road crossing or mortality location to examine landscape structure of woody cover
To explore if there were differences in the spatial structure of woody cover based on the four objectives we defined a set of seven class-level landscape metrics: edge density (ED, m/ha), percent land cover (PLAND, %), patch density (PD, patches/ha), mean patch area (AREA_MN, ha), mean nearest neighbor (ENN_MN, m), largest patch index (LPI, %), and aggregation index (AI, %). These metrics have previously been used to evaluate the spatial structure of woody cover and landscape features for ocelots across their geographic range (Jackson et al. 2005; Garcia et al. 2019; Schmidt et al. 2020; Lombardi et al. 2021; Blackburn et al. 2021a, 2022). We used package landscapemetrics in Program R (Hesselbarth et al. 2019) to calculate landscape metrics of woody cover at each spatial extent for all crossing and roadkill locations. Some ocelots crossed paved roads more than one time so metrics of successful crossing locations for individual ocelots were averaged to obtain an average set of metrics for each ocelot. To compare metrics from successful crossing locations to the historic database of ocelot-road mortality in southern Texas, we used landscape metric values for the remaining unmonitored roadkill ocelots originally used by Blackburn et al. (2021a) in their study.
Statistical Analyses
Once metrics were quantified for each successful crossing and roadkill location, we used a permutational multivariate analysis of variance (PERMANOVA) to determine whether there were significant differences in the landscape structure of woody cover between ocelot successful crossing locations and roadkill locations using Primer v7 (PRIMER-e, Albany Auckland, New Zealand; Anderson 2001). Metrics were normalized before analysis and converted to a dissimilarity matrix based on Euclidean distances (Anderson 2001). We conducted three different PERMANOVAs at each spatial extent: (1) the mean value of metrics of successful crossings of all monitored ocelots to the locations of all known ocelot roadkills (hereafter, all-ocelot analysis); (2) a paired analysis of ocelots that were roadkilled while monitored of the mean value of metrics of successful crossings to their respective roadkill locations (hereafter, paired analysis); (3) the mean value of metrics of successful crossings of roadkilled ocelots to ocelots with an unknown fate (hereafter, successful crossing analysis). If a PERMANOVA was significant, we conducted a one-way similarity percentages (SIMPER) analysis to determine the relative percentage that differences in individual metrics contributed to overall differences in the landscape structure (Clark 1993). We also conducted a separate principal components analysis (PCA) for each spatial extent and PERMANOVA design to identify patterns in the landscape structure of crossings/roadkills and how these patterns relate to individual landscape metrics. We conducted all PCAs in R v4.1.2 using the prcomp function in the Stats package; data were centered and scaled before analysis.
Results
From 1982 to 2001, we documented 69 paved road crossing events from 16 individuals (7 females, 9 males). Of these individuals, 5 (1 female, 4 males) suffered vehicle-attributed road mortalities. The average number of paved road crossings was 4.31 (range: 1–15) for males and 5 (range: 1–9) for females. Of the monitored individuals who died from vehicle-attributed mortality, 1 female crossed paved roads at least 7 times, whereas the 4 males each had at least 2 crossings. We documented 26 road killed ocelots between 1982 and 2017, 5 of which were being monitored at the time (see Fig. 1 in Blackburn et al. 2021a).
We found statistically significant differences in the attributes of the locations of successful crossings to roadkill locations for the all-ocelot analysis at the 150 m spatial extent (pseudo-F1,41 = 4.85, P(perm) = 0.008, permutations = 9949). The SIMPER analysis revealed that the largest patch index contributed the greatest to the differences between successful crossings and roadkill locations (15.94%), followed by mean patch area (15.44%), percent landscape cover (15.18%), aggregation index (14.53), Euclidean nearest neighbor (13.47%), edge density (13.08%) and patch density (12.36%). All other PERMANOVAs at the other spatial extents showed no significant differences at the 0.10 level (Supplemental Table A1-3).
Our PCA results, however, revealed consistent clustering patterns, especially when comparing successful crossings to roadkill locations. Ordination clustering patterns were most apparent at finer spatial extents (150–300 m) where there was the greatest distinction between roadkill locations and mean successful crossing locations. Although we did not see statistically significant effects at 300 m buffer, the pattern is similar to the 150 m buffer result (Fig. 3). At broader spatial extents, there was more overlap between successful crossing and roadkill locations. At each spatial extent, the structure of woody cover for roadkill locations tended to have greater Euclidean distances between patches, higher patch densities, and smaller patch areas than the successful crossing locations (Figs. 3 and 4). This pattern held true for the all-ocelot analysis and the paired analysis. For the successful crossings analysis, ocelots that were road killed tended toward locations with higher patch densities, higher mean patch areas, and greater Euclidean distances between patches (Fig. 5).

Principal components analysis plot of woody cover metrics of the average successful crossing locations of all monitored ocelots and all known roadkilled locations between 1982 and 2001 at each of five spatial extents. Principal component (PC) axes 1 and 2 are shown. Metrics included percent landscape (PLAND; %), patch density (PD; patches/100 ha), edge density (ED; m/ha), mean patch area (AREA_MN; ha), largest patch index (LPI), aggregation index (AI), and Euclidean nearest neighbor distance (ENN_MN; m). Correlation between landscape metrics and PC axes are represented by radial lines

Principal components analysis plot of woody cover metrics of the average successful crossing locations of individual ocelots and their roadkill location at each of five spatial extents. Principal component (PC) axes 1 and 2 are shown. Metrics included percent landscape (PLAND; %), patch density (PD; patches/100 ha), edge density (ED; m/ha), mean patch area (AREA_MN; ha), largest patch index (LPI), aggregation index (AI), and Euclidean nearest neighbor distance (ENN_MN; m). Correlation between landscape metrics and PC axes are represented by radial lines

Principal components analysis plot of woody cover metrics of the average successful crossing locations of monitored ocelots that were eventually roadkilled compared with cats with an unknown fate at each of five spatial extents. Principal component (PC) axes 1 and 2 are shown. Metrics included percent landscape (PLAND; %), patch density (PD; patches/100 ha), edge density (ED; m/ha), mean patch area (AREA_MN; ha), largest patch index (LPI), aggregation index (AI), and Euclidean nearest neighbor distance (ENN_MN; m). Correlation between landscape metrics and PC axes are represented by radial lines
Discussion
Within the road effect zone, landscape structure within 300 m of highways is likely to be the best indicator of whether a crossing event will be successful or unsuccessful (i.e., roadkill) for ocelots in Southern Texas. Ordination clustering of roadkill locations of telemetered ocelots suggests landscape structure is of lower quality compared to mean values of successful crossing locations. Locations with road mortalities of both telemetered and unmonitored ocelots all had longer distances between woody patches and smaller patch sizes, which is indicative of low suitable woody cover for ocelots (Lombardi et al. 2021). These results put previous studies on ocelot road mortalities into context, where road mortality sites had more woody cover, patches aggregated closer together and larger patches than random sites on the landscape (Blackburn et al. 2021a; Schmidt et al. 2020). However, ordination clustering indicates ocelots who successfully cross paved roads are doing so between patches close to the highway that are larger and closer together, an indication of a typical high landscape structure suitability (Lombardi et al. 2021).
Ordination clustering in poor landscape structure in areas immediately closer to highways is not surprising, particularly in regard to highway construction and design. Typically, when roads are constructed through areas of intact habitat, there is a considerable amount of disturbance in areas within the first 50-m from the center of the highway, depending on the size of the road. Highways and other paved roads typically also include paved shoulders and 20 m of mowed rights of way for drainage and utility lines. Often, when paved roads are built, large intact patches of woody cover are fragmented into smaller patches or are cleared outright. As a result, this directly impacts the movement and connectivity of wildlife species, especially those like ocelots who prefer woody environments (Jackson et al. 2005; Schmidt et al. 2020; Blackburn et al. 2021a; Lombardi et al. 2021). These 150 m areas around roadways were found to be significant in identifying where ocelots were more likely to be killed across southern Texas compared to randomized locations on the landscape. Blackburn et al. (2021a) reported these areas contained more woody cover than random locations. These results coupled with our findings suggest ocelots may seek out larger patches of woody cover in areas closer to roads, compared to random locations but more distant and smaller patches may lead to an increased likelihood of mortality rather than successful crossings.
Of monitored individuals that did succumb to vehicle mortality in locations with poorer landscape structures, 75% were male. However, across all mortalities examined regardless of a telemetered individual, males represented over half of ocelot road mortalities (Blackburn et al. 2021b). Upon closer inspection of the paired analysis results, we discovered of the five roadkill locations of monitored cats, one individual (M65) showed a successful/roadkill pattern that was opposite to the other four cats. This individual successfully crossed in areas with high PD and ENN_MN, smaller AREA_MN at fine spatial extents (150 and 300 m), more associated with roadkill locations of the other cats but died in an area of higher-quality structure that most resembled where the other cats successfully crossed.
Male ocelots are highly territorial and occupy large home ranges to encompass 3–5 females (Logan and Sweanor 2001; Hunter 2015). In areas with a mosaic of urban and road development, agriculture, and natural cover, male ocelots are more likely to encounter roadways. Veals (2022a) indicated male ocelots were more likely than females to use areas closer to high-traffic volume roads, which are attributed to having higher mortality risks (Blackburn et al. 2021b). Dominant males are likely to occupy areas of the larger and more suitable woody structure and cover (Veals et al. 2022a; Lombardi et al. 2021), which may aid in selecting areas to cross if a paved road is found in their home range. Whereas smaller, younger, or more subordinate males may occupy a more marginal habitat with smaller and distant patches of cover, which may influence their ability to find safe places to cross. However, the one male that died in areas with the best woody structure of all monitored ocelots, suggests that there is always a chance that crossing roads is dangerous for ocelots.
At the broader landscape-level (450–1050 m) REZs, we did not observe the same level of clustering of habitat characteristics regardless of whether ocelots successfully crossed or died. This suggests that beyond the suggested REZ (> 450 m), ocelots may be using woody cover in proportion to what is available on the landscape and less likely because of a direct road effect. Veals et al. (2022a) showed that ocelots in this region exhibited proportional use of woody cover at the third order (home range) in areas proximate to paved roads of different traffic volumes. In fragmented areas as seen in this study, the woody structure at the landscape level (Lombardi et al. 2021) along with the connectivity of patches (Veals et al 2022b) will be important to steer ocelots to an ideal woody structure adjacent to areas that appear more conducive to successful crossings (i.e., larger patches [> 0.87 ha; Lombardi et al. 2021] close together within 300 m of roadways).
Increased ordination clustering at finer spatial extents (< 300 m) compared to more landscape-level extents need to be accounted for when designing where wildlife crossing structures are built. If ocelots are using woody cover structures available on the landscape at larger extents around roads, then landscape-level factors to get ocelots to roadways only represent one-half of the puzzle. Wildlife crossing structures in this region have historically been constructed in areas where ocelots were killed by vehicles, regardless of the landscape-level extent (Blackburn et al. 2022). Blackburn et al. (2021a) showed that ocelot road mortality sites had a 20% greater proportion of woody cover compared to randomized sites at 150 m spatial extents. However, this study now demonstrates this may not represent an ideal structure conducive to successful crossing events.
Our results indicate the importance of considering landscape structure and its influence on species ecology within appropriate spatial extents (Moraga et al. 2019; Blackburn et al. 2021a; Lombardi et al. 2020, 2021; Seo et al. 2021). Knowledge of the REZ effects for different species is critical when designing road mitigation measures. Road effect zones can differ widely for wildlife species and road types (Benítez-López et al. 2010). Larger carnivores such as grizzly bears (Ursus arctos) shift their REZ (0.2 to 0.9 km) when encountering major and minor roads (Kasworm and Manley 1990), whereas western chimpanzees (Pan troglodyes verus) have much larger REZs (major: 17.2 km; minor: 5.4 km; Andrasi et al. 2021). However, REZ can also vary by sex; female moose in Alaska have larger REZs than male moose (Shanley and Pyare 2011). Beyond the identification of REZs for species crossing and mortality locations, landscape metrics can serve important roles in wildlife crossing site selection (Wiens 1989, Snow et al. 2014). Our study demonstrates that future research in REZ of ocelots or similar-sized carnivores should incorporate landscape metrics.
We acknowledge potential shortcomings of this research, including non-significant differences between roadkill and successful crossings. We contend, however, that the significant differences observed at the 150 m extent for all ocelots, and the consistency in ordination clustering in the PCA in the all-ocelot and the paired-ocelot analyses provide convincing evidence of differences in landscape structure between successful crossing locations and roadkill locations despite the small sample size. Ocelots have been endangered in the United States since 1982, and during the time of telemetry monitoring (1982–2001), this population represented > 90% of the known individuals in the United States. Consequently, this study represents the total available data for ocelots in the United States in a fragmented landscape with varying road densities. Studies like this form a baseline analysis for future studies on rare and endangered species when data are lacking to make strong inferences. Further, traffic volume and road characteristics were similar on all paved roads in our study area, precluding our ability to conduct a meaningful analysis to discern potential effects on successful crossing or road mortality events.
Based on the results of this study, we recommend that placement of wildlife crossing structures factor in spatial structure of the woody cover within 300 m of highways and not select locations solely based on roadkilled individuals (Teixeira et al. 2017; Ascensão et al. 2019). Furthermore, connectivity at the landscape level will be an important aspect to consider in further research to facilitate ocelot movement to and from the highway (Veals et al 2022b). Vegetation restoration work should be focused on maintaining and restoring patches of woody cover to increase the size and decrease the distance between adjacent patches proximate to highways. Further, vegetation restoration efforts beyond 300 m from highways should focus on a landscape level approach, using suitable woody structure characteristics (Lombardi et al. 2021), and connectivity (Veals et al 2022b) to ensure woody cover is conducive to a higher probability of ocelot use and home range placement (Veals et al. 2022a).
Data availability
Metadata used in the manuscript is supplied as supplemental material.
Code availability (software application or custom code)
N/A
References
Aaron Economic Consulting (2017) South Padre Island Resort Market Analysis, Aaron Economic Consulting, LLC
Ament R, Clevenger AP, Yu O, Hardy A (2008) An assessment of road impacts on wildlife populations in US National Parks. Envir Manag 42:480–496
Anderson MJ (2001) A new method for non-parametric multivariate analysis of variance. Aust Ecol 26:32–46
Andrasi B, Jaeger JAG, Heinicke S, Metcalfe K, Hockings KJ (2021) Quantifying the road-effect zone for a critically endangered primate. Conserv Let 14:e12839
Ascensão F, Kindel A, Teixeira FZ, Barrientos R, D’Amico M, Borda-de-Água L, Pereira HM (2019) Beware that he lack of wildlife mortality records can mask a serious impact of linear infrastructures. Glob Ecol Conserv 19:e00661
Benítez-López A, Alkemade R, Verweij PA (2010) The impacts of roads and other infrastructure on mammal and bird populations: a meta-analysis. Biol Conserv 143:1307–1316
Bennett AF (1991) Roads, roadsides and wildlife conservation: a review. In: Saunders DA, Hobbs RJ (eds) Nature conservation 2: the role of corridors. Surrey Beatty, Chipping Norton, pp 99–117
Blackburn A, Anderson CJ, Veals AM, Tewes ME, Wester DB, Young JH Jr, DeYoung RW, Perotto-Baldivieso HL (2021a) Landscape patterns of ocelot–vehicle collision sites. Landsc Ecol 17:1–5
Blackburn A, Heffelfinger LJ, Veals AM, Tewes, ME, Young Jr JH (2021b) Cats, cars, and crossings: the consequences of road networks towards the conservation of an endangered felid. Glob Ecol Conserv :e01582
Blackburn A, Veals AM, Tewes ME, Wester DB, Young JH Jr, DeYoung RW, Perotto-Baldivieso HL (2022) If you build it, will they come? A comparative landscape analysis of ocelot roadkill locations and crossing structures. PLoS ONE 17:e0267630
Chruszcz B, Clevenger AP, Gunson KE, Gibeau ML (2003) Relationships among grizzly bears, highways, and habitat in the Banff-Bow Valley, Alberta, Canada. Can J Zool 81:1378–1391
Clark (1993) Nonparametric multivariate analyses of changes in community strucure. Aust J Ecol 18:117–143
Clevenger AP, Chruszcz B, Gunson KE (2003) Spatial patterns and factors influencing small vertebrate fauna road-kill aggregations. Biol Conserv 109(1):15–26
Clevenger AP, Huijser MP (2011) Wildlife crossing structure handbook: design and evaluation in North America. Report FHWA-CFL/TD-11–003. U.S. Department of Transportation
Ditmer M, Rettler S, Fieberg J, Iaizzo P, Laske T, Noyce K, Garshelis D (2018) American black bears perceive the risks of crossing roads. Behav Ecol 29:667–675
Dulac J (2013) Global land transport infrastructure requirements. Estimating road and railway infrastructure capacity and costs to 2050. OECD, International Energy Agency, Paris
Forman RT, Alexander LE (1998) Roads and their major ecological effects. Annu Rev Ecol Syst 29(1):207–231
Forman RT, Deblinger RD (2000) The ecological road-effect zone of a Massachusetts (USA) suburban highway. Conserv Biol 14:3646
Forman RT, Sperling D, Bissonette JA, Clevenger AP, Cutshall CD, Dale VH, Fahrig L, France RL, Goldman CR, Heanue K, Jones J (2003) Road ecology: Science and solutions. Island press
García RS, Botero-Cañola S, Sánchez-Giraldo C, Solari S (2019) Habitat use and activity patterns of Leopardus pardalis (Felidae) in the Northern Andes, Antioquia, Colombia. Biodiversity 20:5–19
Griffith G, Bryce S, Omerik J, Rogers A (2007) Eco-regions of Texas. Austin, TX, USA, Texas Commission on Environmental Quality
Grilo C, Borda-de-Água L, Beja P, Goolsby E, Soanes K, le Roux A, Koroleva E, Ferreira FZ, Gagné SA, Wang Y, González-Suárez M (2021) Conservation threats from roadkill in the global road network. Glob Ecol Biogeog 30:22002210
Haines AM, Tewes ME, Laack LL (2005) Survival and sources of mortality in ocelots. J Wildl Manag 69:255–263
Hesselbarth MH, Sciaini M, With KA, Wiegand K, Nowosad J (2019) landscapemetrics: an open-source R tool to calculate landscape metrics. Ecography 42:1648–1657
Hunter L (2015) Wild cats of the world. Bloomsbury Publishing
Jackson ND, Fahrig L (2011) Relative effects of road mortality and decrease connectivity on population genetic diversity. Biol Conserv 144:3143–3148
Jackson VL, Laack LL, Zimmerman EG (2005) Landscape metrics associated with habitat use by ocelots in south Texas. J Wildl Manage 69:733–738
Lombardi JV, Sergeyev M, Tewes ME, Schofield LR, Wilkins RN (2022) Spatial capture-recapture and LiDAR-derived vegetation metrics reveal high densities of ocelots on Texas ranchlands. Front Conserv Sci 14:136
Kasworm WF, Manley TL (1990) Road and trail influences on grizzly bears and black bears in northwest Montana. Bears Biol Manage 8:79–84
Klar N, Fernández N, Kramer-Schadt S, Herrmann M, Trinzen M, Büttner I, Niemitz C (2008) Habitat selection models for European wildcat conservation. Biol Cons 141:308–319
Lehnen SE, Sternberg MA, Swarts HM, Sesnie SE (2021) Evaluating the population connectivity and targeting conservation action for an endangered cat. Ecosphere 12:e03367
Leslie DM Jr (2016) An international borderland of concern: conservation of biodiversity in the Lower Rio Grande Valley. US Geological Survey, Stillwater
Lewis JS, Rachlow JL, Horne JS, Garton EO, Wakkinen WL, Hayden J, Zager P (2011) Identifying habitat characteristics to predict highway crossing areas for black bears within a human-modified landscape. Landsc Urb Plan 101:99–107
Logan KA, Sweanor LL (2001) Desert puma: evolutionary ecology and conservation of an enduring carnivore. Island Press, Washington, DC
Lombardi JV, Perotto-Baldivieso HL, Tewes ME (2020) Land cover trends in South Texas (1987–2050): potential implications for wild felids. Remote Sens 12:659
Lombardi JV, Perotto-Baldivieso HL, Sergeyev M, Veals AM, Schofield L, Young JH Jr, Tewes ME (2021) Landscape structure of woody communities for endangered ocelots in southern Texas. Remote Sens 13:4001
Main MB, Allen GM (2002) Landscape and seasonal influences on roadkill of wildlife in southwest Florida. Fla Sci 65:149–158
Meijer JR, Huijbregt MAJ, Schotten KCGJ, Schipper AM (2018) Global patterns of current and future road infrastructure. Envir Res Lett 13:604006
Moraga AD, Martin AE, Fahrig L (2019) The scale of effect of landscape context varies with the species’ response variable measured. Landsc Ecol 34:703–715
Neumann W, Ericsson G, Dettki H, Bunnefeld N, Keuler NS, Helmers DP, Radeloff VC (2012) Difference in spatiotemporal patterns of wildlife road-crossings and wildlife-vehicle collisions. Biol Conserv 145:70–78
Norwine J, Kuruvilla J (2007) The changing climate of south texas 1900–2100: Problems and prospects, impacts, and implications. CRESTRESSACA, Texas A&M University-Kingsville, Kingsville
Poessel SA, Burdett CL, Boydston EE, Lyren LM, Alonso RS, Fisher RN, Crooks KR (2014) Roads influence movement and home ranges of a fragmentation-sensitive carnivore, the bobcat, in an urban landscape. Biol Conserv 180:224–232
Qin X, Wang Y, Cui S, Liu S, Liu S, Wangari VW (2022) Post-assessment of the eco-environmental impact of highway construction–A case study of Changbai Mountain Ring Road. Envir Imp Assess Rev 98:106963
Schmidt GM, Lewison RL, Swarts HM (2020) Identifying landscape predictors of ocelot road mortality. Landsc Ecol 35:1651–1666
Semlitsch RD, Ryan TJ, Hamed K, Chatfield M, Drehman B, Pekarek N, Spath M, Watland A (2007) Salamander abundance along road edges and within abandoned logging roads in Appalachian forests. Conserv Biol 21:159–167
Seo H, Choi C, Lee K, Woo D (2021) Landscape characteristics based on effectiveness of wildlife crossing structures in South Korea. Sustainability 13:675
Shanley CS, Pyare S (2011) Evaluating the road-effect zone on wildlife distribution in a rural landscape. Ecosphere 2:1–16
Snow NP, Williams DM, Porter WF (2014) A landscape-based approach for delineating hotspots of wildlife-vehicle collisions. Landsc Ecol 29:817–829
Taylor BD, Goldingay RL (2010) Roads and wildlife: impacts, mitigation and implications for wildlife management in Australia. Wild Res 37:320–331
Teixeira FZ, Kindel A, Hartz SM, Mitchell S, Fahrig L (2017) When road-kill hotspots do not indicate the best sites for road-kill mitigation. J App Eco 54:1544–1551
Texas Department of Transportation (TXDOT) (2022) http://gistxdot.opendata.arcgis.com/datasets/txdot-aadt-annuals
Tremblay TA, White WA, Raney JA (2005) Native woodland loss during the mid-1900s in Cameron County, Texas. Southwest Nat 50:479–519
United States Fish and Wildlife Service (USFWS) (2017) Laguna Atascosa National Wildlife Refuge Visitor Guide Booklet. https://www.fws.gov/uploadedFiles/LA%20NWR%20Tear%20-Booklet2017%20(3).pdf. Accessed 11 July 2019
van der Ree R, Smith DJ, Grilo C (2015) Handbook of road ecology. Wiley
Veals AM, Holbrook JD, Blackburn A, Anderson CJ, DeYoung RW, Campbell TA, Young JH Jr, Tewes ME (2022a) Multiscale habitat relationships of a habitat specialist over time: the case of ocelots (Leopardus pardalis) in South Texas from 1982–2017. Ecosphere 13:e4204
Veals AM, Holbrook JD, Cherry MJ, Campbell TA, Young JH, Tewes ME (2022b) Landscape connectivity for an endangered carnivore: habitat conservation and road mitigation for ocelots in the US. Landsc Ecol 1–9
Wiens JA (1989) Spatial scaling in ecology. Funct Ecol 3:385–397
Acknowledgements
We thank the previous graduate students and technicians who helped collect the telemetry data and collected information on roadkill data used in this analysis. We thank the US Fish and Wildlife Service for cooperative assistance with ocelot captures and monitoring and mortality locations. This manuscript is #22-117 of the Caesar Kleberg Wildlife Research Institute.
Funding
This analytical research and funding for JVL, CJA, AB, and TJY. were funded by the Texas Department of Transportation.
Author information
Authors and Affiliations
Contributions
Conceptualization, C.J.A., A.B., J.V.L., T.J.Y.; methodology, C.J.A., A.B., J.V.L., T.J.Y.; formal analysis, J.V.L., T.J.Y.; investigation, J.V.L., T.J.Y.; resources, J.V.L., C.J.A., T.J.Y. A.B.; visualization, J.V.L., T.J.Y.; writing—original draft preparation, J.V.L. and T.Y. writing—review and editing, all authors; funding acquisition, M.E.T., supervision J.H.Y. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Conflicts of interest/competing interests
N/A
Additional information
Jason Lombardi and Thomas Yamashita are shared-first authors and contributed equally on this paper.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lombardi, J.V., Yamashita, T.J., Blackburn, A. et al. Examining the Spatial Structure of Woody Cover Within a Highway Road Effect Zone for Ocelots in Texas. Urban Ecosyst 26, 1057–1069 (2023). https://doi.org/10.1007/s11252-023-01350-y
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-023-01350-y