
Abstract
The soundscape was recorded in four selected places in Sequoia National Park CA, to quantify and assess the diurnal and seasonal character of the park’s soundscape. The recording sites were selected to represent a combination of elevation and vegetation diversity. Hour-long sound recordings were made by four individuals at each place during fall, spring, summer and winter at dawn, midday, dusk, and midnight with identical recording instrumentation. The recordings of the soundscape were made in an old growth forest (Crescent Meadow), in a foothill oak savanna (Sycamore Spring), in an upland savanna chaparral (Shepherd Saddle) and in a foothill riparian location adjacent to the Kiawah River (Buckeye Flat). Sound recordings were analyzed using a normalized Power Spectral Density (PSD) algorithm and partitioned into 1 kHz intervals based on 12 subsamples from each of the 64 h-long sound recordings. Biological signals (biophony) were based on the highest PSD value within the range of 2–8 kHz. A multilevel analysis (MLA) was used to examine temporal patterns of biophony at four locations in Sequoia National Park. Unsupervised Landsat Thematic Mapper Satellite Imagery identified 25 vegetation regimes in Sequoia National Park. Satellite signatures of the habitat where recordings were made were extracted from the imagery to scale to the region.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The National Parks in the USA are locations where people go to experience nature in its splendor. Untouched by humans and set aside for future generations to experience, the National Parks represent society’s effort to preserve a unique diversity of ecosystems and their services. Sequoia and Kings Canyon National Parks represent two of the National Parks located in the southern Sierra Nevada mountain range (Strong 1996). Visitors to National Parks rate their acoustic experiences as high as those that are visual (Pilcher and Turina 2008). Determining how the soundscape varies across a National Park is of considerable interest to the National Park Service (National Park Service 1995). An annotated bibliography of visitor experience and soundscapes was developed to provide a base of information about soundcapes in National Parks (Pilcher and Turina 2008).
Noise from recreational vehicles, air flight tours and nearby roads impact the natural sounds that occur in many National Parks (Habib et al. 2007; Dumyahn and Pijanowski 2011). An assessment of the frequency and relative contribution of human generated sounds across a park is also an important component of national park management (Dumyahn and Pijanowski 2011).
New methods of evaluating the character of a landscape from a bioacoustics perspective combines the sound-producing properties of the vocal organisms and other sounds within that habitat to establish base line data of the landscape’s acoustic signature.
Schafer (1977, 1994) defines a soundscape as a region of acoustic activity homogeneous in a feature of interest. Our concept of a soundscape is more analogous to a landscape and thus we consider the soundscape to represent the spatial variation of sounds that are emitted from the elements of a landscape. Biologists have long recognized that vertebrate species vocalizations vary according to the habitats which the species prefer (Baptista and Gaunt 1997; Hopp et al. 1998). We also recognize that sound and the habitat conditions within a landscape are intimately linked through the biological and physical characteristics which synergistically produce the soundscape. We therefore developed a general approach to record, quantify and analyze the acoustics in different habitats to characterize their acoustic attributes (Krause 1987, 1998, 2002).
Soundscape signatures comprise three components: biophony, geophony and anthrophony (Gage et al. 2001, 2004; Napoletano 2004; Pijanowski et al. 2011b). Biophony is the combined sound that living organisms produce in a given habitat. Geophony is comprised of geophysical sounds in the environment, such as the sounds of wind, thunder, water flow, earth movement, etc. Anthrophony is sound produced by human-generated mechanical sounds, such as aircraft, automobiles, generators, snowmobiles, jet-skis, radios, television sets, boom boxes, bells, wind turbines or automobile sound systems. Soundscapes represent the holistic combination of biophony, geophony and anthrophony representative of a particular geographic space (landscape).
The above classification assists in the interpretation of the soundscape. We argue that the measurement of the soundscape is an ideal metric to assess landscape stress whether the stress is caused by physical (e.g. hurricane), biological (e.g. invasion) or by human technologies (e.g. snowmobile). Sounds vary over time of day and season and can thus capture temporal ecosystem dynamics. The scope of this investigation has been to record base line acoustics recordings sampled from key landscapes that are characteristic of the Sequoia National Park and to evaluate them with regard to the biological component of the soundscape.
The objectives of the study were to: (1) record sounds at four places within Sequoia National Park to measure variation in the diurnal and seasonal sounds within the landscape to demonstrate to park managers the variability in the soundscape; (2) develop soundscape metrics from the recordings to enable a comparison of the biological component of the soundscape over the diurnal and seasonal cycle; and (3) determine the significant differences between diurnal and seasonal change for all recording places and within each recording place.
Methods
Recording locations
The four places (Table 1) were selected, with the assistance of NP personnel, to represent a combination of elevation and vegetation diversity within the park (Tweed 1982; Strong 1996) in order to capture the dynamic range of the soundscape within the selected habitats in Sequoia National Park during four seasons (fall, winter, spring and summer) at different times of day (morning, midday, evening and night). Crescent Meadow, located at 7,000 feet (2154 m), N36° 33.364 W118° 44.867, is a meadow rimmed by sequoia trees (Sequoia sempervirens). John Muir called Crescent Meadow the ‘gem of the Sierra’. Another place selected for sound recording was Shepherd Saddle, located at 3,000 feet (925 m), N36° 29.470 W118° 51.142, is a dry savannah chaparral. Buckeye Flat, located at 2,900 feet (890 m), N36° 31.185 W118° 45.692 is a riparian area associated with the Kaweah River. Sycamore Spring, located at 2,100 feet (645 m), N36° 29.470 W118° 51.225, is a foothill site dominated by oak savannah. Four individuals used calibrated recording instrumentation to record sounds during fall, winter, spring and summer. Recordings of the sounds were made at each of the four sites for approximately 60 min during morning, midday, evening and night. The recording location at Buckeye Flat was changed after the fall recording, as stream flow sounds (geophony) saturated the acoustic signal.
Recording equipment
The recording equipment used was a Sony PCM M1 DAT tape recorder; a Sony MDR 7506™ headphone; a Sennheiser MKH 30™ microphone; a Sennheiser MKH 40™ (cardioid) microphone; a Sound Devices™ Mix/preamp; a Rykote™ shock mount suspension units w/pistol grip; a Rykote™ zeppelin windscreen; a Rykote™ high wind cover; tripod (microphone mount); and miscellaneous cables, connectors, batteries and DAT™ tape. Recording instrumentation was calibrated to a common recording level prior to recording in the field. Sound recordings were made at a location within each place to represent the general characteristics of the habitat. An hour-long high quality recording was made at each of the four places four times per day during a selected day during each season of the year resulting in 64 h of sound recordings.
Data processing
The data were transferred from DAT (44.1 kHz and 16 bit sampling) M-S to matrixed stereo on both hard drive (wav format) and archived to stereo-encoded audio CDs. CDs containing raw wav files were sent to the Remote Environmental Assessment Laboratory at Michigan State University and were archived on a terabyte server for later quantitative processing (see below). A web site provides access to a subset of the recordings [(http://real.msu.edu/projects/one_proj.php?proj=snp) for pictures of the places where recordings were made, the soundscape samples, associated sonograms and the soundscape metrics used in this analysis].
Each of the 64 h-long sound recordings was sub-sampled by extracting a 30 s sound at 5 min intervals, thus producing 12 sub-samples from each recording resulting in a potential of (12 × 4 × 4 × 4) 768 sound files. A few recordings were slightly shorter than 1 h so 759 sound samples were used in the analysis. The samples were converted to 22 kHz monaural. A normalized Power Spectrum Density value (PSD in Watt/Hz) (Welch 1967) was computed for each 1 kHz interval for all recordings by running a script programmed by Gage and Joo using MATLAB (Gilat 2004).
Data analysis
Biophony, the biological sounds produced from vocal organisms within a landscape, generally occurs at acoustic frequencies from 2 to 8 kHz (Pijanowski et al. 2011b). We determined biophony to be that with the highest PSD between 2 to 8 kHz. A multilevel analysis (MLA) was used to examine temporal patterns of biophony at four locations, because the biophony measures in this study contain hierarchical temporal and spatial data structure (Merlo et al. 2005; Gelman and Hill 2007). The data were grouped into two levels of structure: (1) level 1 considers variation of temporal variables (e.g. time of day and season); (2) level 2 considers variation in location (e.g. four places) as a random effect.
In MLA, we used biophony as a dependent variable, and temporal variables include four times of day and the four seasons as fixed effects. In this paper, MLA was implemented using function lmer (Bates and Maechler 2010) in R-package (Ihaka and Gentleman 1996, R development Core Team 2010).
We developed a multilevel linear model with the temporal variables season and time of day as fixed effects (Table 2, Model 2). The equation for this model is:
where Y ij is the arcsine of biophony values measured as PSD derived from each individual sound sample. Y ij values were arcsine-transformed to change a proportional data type to a continuous type to conform to the MLA statistical model based on a parametric statistical test. β o is the intercept (the overall grand mean) of the linear model, and. β 1 accounts for the coefficient of the temporal variables at level 1. ɛ j is random effect residuals at the place level (level 2), and ɛ i is residuals at the temporal level (level 1). An intra-class correlation method (ICC) was computed to determine the correlation of the data collected within each place (Merlo et al. 2005). Akaike’s Information Criterion (AIC) value and likelihood ratio tests were used to find the best-fit multilevel linear model for the data (Akaike 1981).
Results
The results of the multilevel analysis (MLA) are summarized in Table 2. Model 1 showed that it was appropriate to use a multilevel analysis because the ICC was 0.3629, implying that biophony measures were correlated within a place. As the AIC value in Model 2 is lower than that in Model 1 (Model 1 AIC = 572; Model 2 AIC = 459), Model 2 was determined as the best MLA model. A likelihood ratio test between the two MLA models showed that ‘Time-of-day’ (RECORD_TIME), ‘Months’ (SEASON), and interaction between the two factors were significant variables at level 1 (Χ 2 df=15 = 256.07; P < 0.001). In Model 2, the proportional change in variance showed that 27.33% of β variance by the temporal variables at level 1 including Time of day (9.50%), months (13.75%), and interaction between month and Time-of-day (4.1%).
The multilevel analysis showed significant temporal variability of biophony occurred at the study locations. The results from this multilevel analysis indicated that month and time-of-day contribute significantly to biophony variability (Fig. 1). By post-hoc pairwise multiple comparison, the biophony measures at nightime (21:00) in fall (October) and morning (6:00) in spring (May) in model 2 were significantly higher than those in the other months and times of day, implying that the high level of biophony is produced by the vocal activities of birds in the dawn chorus) and of insects in the night chorus at the study locations. In contrast, the biophony values were significantly lower during daytime (12:00) in the fall, and evening and nightime in winter, indicating that the least vocal activities by terrestrial organisms occurred during these periods.

Temporal changes in biophony for four different times of day and four different seasons. Lowercase letters refer to means contrasts among different times of day and seasons using Tukey’s HSD tests (P < 0.05). Values are means ± SE
To further examine the characteristics of the temporal change at each of the recording places, we used a similar statistical process. The Buckeye Flat (BF) site was dominated by the sounds of the nearby river and showed little significant difference in time of day or season. However, during the fall in the evening the insect chorus dominated the soundscape and the biophony was significantly greater at this time (Fig. 2; upper left panel).

Temporal changes in biophony for four different times of day and four different, for each of the recording places selected in the Sequoia National Park. These were Buckeye Flat (BF) Crescent Meadow (CM), Shepherd Saddle (SH) and Sycamore Spring (SY) (Table 1). Lowercase letters refer to means contrasts among different times of day and seasons using Tukey’s HSD tests (P < 0.05). Values are means ± SE
The Crescent Meadow (CM) site had the highest elevation of the four recording places. The wet meadow was surrounded by giant Sequoia trees and was the most distant from the base camp and difficult to get to (especially in winter). At Crescent Meadow (CM) there were no seasonal differences in biophony during the daytime. The evening biophony was significantly different from the daytime biophony, except during winter when it was biologically very quiet. The nightime biophony was similarly quiet, and surprisingly, in spring at nightime the biophony was significantly lower than in other seasons (F df = 15,169 = 16.076, P < 0.001). The mornings at Crescent Meadow were biologically active, especially in spring when the echo of the dawn chorus was dominant due to the surrounding giant forest. One of the joys of field recording is to be at a place like this and listen to the environmental symphony at different times of the day and season. However it was at Crescent Meadow that a black bear visited the recording station during the fall at nightime and not only worried the recordist but caused the recording session to be shortened because the bear licked the microphone and eventually knocked the recorder off the rock that was used as a recording platform. This was a recording to remember. Fall and summer also exhibited vibrant biophony which was significantly greater than the other times of day at Crescent Meadow (Fig. 2; upper right panel).
Shepherd Saddle (Fig. 2; lower left panel) was most quiet during daytime in fall and during nightime in winter. However, nightime during fall and spring at Shepherd Saddle were the most biophonically active followed by the dawn chorus during spring. Summer morning, nightime and evening had significantly more biophony than any of the daytime biophony values (Fdf=15,169 = 19.291, P < 0.001). Biophony was dominate at nightime during fall, spring and summer. The next most intense time for biophony at Shepherd Saddle occurred during daytime in summer, during the evening in spring and summer, and during spring and summer mornings. The quieter times were during fall and winter (Fig. 2; lower left panel).
The Sycamore Spring (SY) recording place had similar biophony patterns as Shepherd Saddle, but the organisms creating the biophony were different. Acorn woodpeckers (Melanerpes formicivorus) were a dominant source of biophony at this place. Nightime biophony in fall, spring and summer were dominant at Sycamore Spring. The biophony intensity during the morning in spring was similar to morning sounds in summer. The evening sounds in spring and summer were similar to daytime sounds in summer. Biophony during winter nights was not different from biophony during fall, spring and winter daytime, fall and winter evenings and fall and winter morings (F df = 15,180 = 32.596, P < 0.001) .
The analysis of these acoustic observation and the resulting patterns enabled us to examine the biophony signal in detail and to determine the significant patterns of the diurnal and seasonal cycle across all recording places as well as withing each of the different sites. These observations substantiate the theoretical diurnal and seasonal variability patterns illustratedin the BioScience article by Pijanowski et al. (2011b).
Discussion
This study was conducted before the advent of automated digital recorders, before quantitative methods had been developed to process soundscape metrics from digital wav files and before the principles of soundscape ecology were fully elucidated (Pijanowski et al. 2011a, b). However, the digital recordings from the Sequoia National Park Soundscape study were preserved and carefully archived with appropriate metadata. These recordings and early metrics associated with the recordings were placed on a website and are publically accessible (http://real.msu.edu/projects/one_proj.php?proj=snp) We were able to download and analyze the soundscape recordings using new statistical methods that have been developed for analyzing repeated measurements such as those represented in the Sequoia National Park recording dataset. These sound recordings were made during a relatively dry and warm year (2001–2002). The 64 h of recordings represent both a diurnal and a seasonal record of Sequoia NP soundscapes. This acoustic resource provides a reference point to enable biologists and managers to examine these recordings to detect changes in the ecosystems in Sequoia NP due to climate shifts, pollution increases, changes public visit intensity or changes in policy that enable expanded resource exploitation. The careful documentation of the recording location and the storage and access to digital sound archives provides an opportunity to return and record acoustics at the same locations at regular intervals to assess the changing soundscapes that reflect changes in ecosystem dynamics.
The main thrust of this study was to conduct an analysis of the biophony in different seasons and during different times of day. However, during the course of archiving acoustic signals, we did hear many of the prominent vocal avian species which were captured on tape, digitized and stored in a digital archive. We suggest, however, that species specific identities should be assessed by those with more knowledge of the sounds of Sequoia wildlife using the digital archive we have created see (http://www.real.msu.edu/projects/one_proj.php?proj=snp) for a project description and access to the sound library.
In addition, the ability of the public to listen to the soundscape of Sequoia NP over the internet can provide immense enjoyment and a better understanding of the ecological importance of Sequoia NP. The drumming of a Pileated woodpecker (Dryocopus pileatus), the sound of the wing beat of a raven (Corvus corax) as it passes overhead, the call of a western scrub jay (Aphelocoma californica) , the song of black-headed grosbeak (Pheucticus melanocephalus) or the call of the acorn woodpecker (Melanerpes formicivorus) are species recorded in the archive.
The sound recordings have been analyzed for the peak biophony in the sound recordings. Based on the sound recordings made during the four seasons, we were able to examine the general temporal change across recording places as well as examining the temporal change at each of the places where recordings were made. Because of the nature of the sampling technology and the need to use human observers to go to each site to make the recordings, only four locations could be recorded simultaneously.
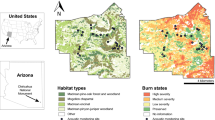
To couple satellite imagery to each recording site to demonstrate the capacity to scale to the Sequoia National Park, we obtained a TM Landsat image of the study region for May 2001 from http://www.landsat.org. Twenty-five cover types ranging from old growth forest to open grassland were identified. Cover types within a radius of 500 m from each recording place were used as place signatures based on an estimate of the mean microphone sensitivity distance. Figure 3 shows the vegetation gradient draped over elevation and the location of the recording locations. The darker shade is old growth forest, lightest shade is grasslands. Snowfields are white (Fig. 3).

A vegetation gradient classified from a Landsat ETM + Scene. The 25 vegetation classes were overlaid onto a digital elevation model of the area. Sampling locations are labeled and circled. Signatures of each site were obtained from within the circles. Dark color represents most dense vegetation. Snowfields (right edge) are white

Habitats identified as being a match with a signature extracted from within each of the sample locations (circles) and matched with the vegetation image shown. Visible areas are habitats similar to that sampled at each sound sampling site. Percent coverage noted below location name
A signature of the vegetation at each place was based on a 500 m circle around the recording location as a general estimate of the recording distance. The resulting areas of the park that matched the signature is shown in Fig. 4 where each panel displays the signature match for each place. For example, other areas, similar to the old growth forest surrounding Crescent Meadow, are located at higher elevations in northern parts of the park (Fig. 4, Crescent Meadow panel). The oak savanna area, represented by the Sycamore Spring recording place, is dominant in the south western region of the park (Fig. 4 Sycamore Spring panel). The dry savanna chaparral, represented by the Shepherd Saddle, occupies the central area of the park (Fig. 4 Shepherd Saddle panel). The riparian site (Buckeye Flat) flows from the north central part of the park to the southwest. The signature search outlined the courses of the main rivers in the park (Fig. 4 Buckeye Flat panel).
Base-line acoustic recordings were collected from representative locations but even the extrapolated areas represented only 9% of the area in Sequoia NP (Fig. 4).
The current use of bioacoustics in conservation is generally that of conducting a census of vocal organisms to inventory the biological diversity to monitor population change and to identify individuals (Baptista and Gaunt 1997). We have carefully calibrated acoustic monitoring instrumentation and collected and archived recorded sounds necessary to compare soundscape changes over time.
Caution is advised in the selection of locations for sound recording as we learned from our placement of a recording site near the Kaweah River. Although our recordings of the American Dipper (Cinclus mexicanus) were memorable, the dominance of the river’s geophony overwhelmed our riparian soundscape, especially during spring runoff. These “flux” patterns of soundscapes (Pijanowski et al. this special issue) were evident in this location. We also recognize that the elevation gradient of the sound recording places influenced our interpretation of spring. For example, spring at Crescent Meadow, due to its higher elevation, is later that at lower elevations. This could be alleviated through the use of automated recording technology but such technology was not available at the time of the study (see below).
Advances in automation and communication of ecological information are emerging paradigms to investigate ecological change (Porter et al. 2005). Parallel advances in sound recording instrumentation and telemetry of acoustic signals provide a significant opportunity to establish permanent recording sites to monitor the Sequoia soundscape at higher temporal resolution than we were able to accomplish by four recording specialists. This could provide park managers information, through listening to the parks changing soundscapes, about the changing character of the park’s ecosystems that will undoubtedly be impacted by natural forces and increases in human disturbance (Habib et al. 2007, Dumyahn and Pijanowski 2011).
References
Akaike H (1981) Likelihood of a model and information criteria. J Econom 16:3–14
Baptista LF, Gaunt SLL (1997) Bioacoustics as a tool in conservation studies. In: Clemmons JR, Buchholz R (eds) Behavioral approaches to conservation in the wild. Cambridge University Press, Cambridge, pp 212–242
Bates D, Maechler M (2010) lme4: Linear mixed-effects models using S4 classes. R package version 0.999375-37, URL http://CRAN.R-project.org/package=lme4
Dumyahn S, Pijanowski B (2011) Beyond noise mitigation: managing soundscapes as common pool resources. Landscape Ecol. doi:10.1007/s10980-011-9637-8
Gage SH, Napoletano B, Cooper M (2001) Assessment of ecosystem biodiversity by acoustic diversity indices. J Acoust Soc Am 109:2430
Gage SH, Ummadi P, Shortridge A, Qi J, Jella P (2004) Using GIS to develop a network of acoustic environmental sensors. Paper presented at ESRI international user conference 2004, San Diego CA, 9–13 Aug 2004, pp 1–10
Gelman A, Hill J (2007) Data analysis using regression and multilevel/hierarchical models. Cambridge University Press, New York, NY, p 625
Gilat A (2004) MATLAB: an introduction with applications, 2nd edn. John Wiley & Sons, Hoboken
Habib L, Bayne E, Boutin S (2007) Chronic industrial noise affects pairing success and age structure of ovenbirds Seiurus aurocapilla. J Appl Ecol 44:176–184
Hopp SL, Owren MJ, Evans CS (1998) Animal acoustic communication: sound analysis and research methods. Springer-Verlag, Berlin, p 421
Ihaka R, Gentleman R (1996) R: A language for data analysis and graphics. J Comput Graph Stat 5(3):299–314
Krause B (1987) Bioacoustics, habitat ambience in ecological balance. Whole Earth Rev 57:1–6
Krause B (1998) Into a wild sanctuary: a life in music and natural sound. Heyday Books, Berkeley, p 200
Krause B (2002) Wild soundscapes: discovering the voice of the natural world. Wilderness Press, Berkeley
Merlo J, Yang M, Chaix B, Lynch J, Råstam L (2005) A brief conceptual tutorial on multilevel analysis in social epidemiology: investigating contextual phenomena in different groups of people. J Epidemiol Commun H 59:729
Napoletano B (2004) Measurement, quantification and interpretation of acoustic signals within an ecological context, MS thesis, Michigan State University, East Lansing
National Park Service (1995) Report on the effects of aircraft overflights on the National Park System, edited by Department of Interior, Denver, CO
Pijanowski B, Farina A, Gage S, Dumyahn S, Krause B (2011a) What is soundscape ecology? An introduction and overview of an emerging new science. Landscape Ecol. doi:10.1007/s10980-011-9600-8
Pijanowski BC, Villanueva-Rivera LJ, Dumyahn SL, Farina A, Krause BL, Napoletano BM, Gage SH, Pieretti N (2011b) Soundscape ecology: the science of sound in the landscape. BioScience 61:203–216
Pilcher E, Turina F (2008) Visitor experience and soundscapes: annotated bibliography. U.S. Soundcape Workshop. National Park Service and Colorado State University. p 39
R Development Core Team (2010) R: A language and environment for statistical computing, ISBN 3-900051-07-0, URL http://www.R-project.org, R Foundation for Statistical Computing, Vienna, Austria
Schafer RM (1977) Tuning of the world. Knopf Press, New York
Schafer RM (1994) The soundscape: our sonic environment and the turning of the world. Destiny Books, Rochester
Strong D (1996) From pioneers to preservationists: a brief history of Sequoia and Kings Canyon National Parks. Sequoia Natural History Association, Three Rivers, CA
Tweed WT (1982) A guide to and history of the western third of the High Sierra Trail, Crescent Meadow to Kaweah Gap, Sequoia National Park. Sequoia Natural History Association, Three Rivers, CA
Welch P (1967) The use of fast Fourier transform for the estimation of power spectra: a method based on time averaging over short modified periodograms. IEEE T Acoust Speech 15:70–73
Acknowledgments
Rudy Trubitt and Jack Hines provided professional recording expertise and conducted recordings at the selected sites during all seasonal visits. They were also instrumental in processing and archiving acoustic samples. Annie Esperanza was our principal contact at Sequoia National Park and provided significant logistical assistance including recording place selection, laboratory and lodging facilities during recording intervals. Dave Graber, Sequoia National Park Research Superintendent, understood our vision and reason for the study and arranged for recording permits and general access to the Park. Brian Napoletano, a graduate student at Michigan State University, accompanied Gage to the Sequoia National Park on one of the four visits. His MS work focused on quantifying acoustic signals. Susan Wang helped with the classification of the Landsat TM Image used to extrapolate recording places to the park habitats. We thank Steve Friedman for providing important insights and suggestions for improving an earlier version of the manuscript. Finally, we thank Bill Schmidt of the National Park Service for his support, interest and vision. His passing is a significant loss to the NPS and we dedicated this article to him.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Krause, B., Gage, S.H. & Joo, W. Measuring and interpreting the temporal variability in the soundscape at four places in Sequoia National Park. Landscape Ecol 26, 1247–1256 (2011). https://doi.org/10.1007/s10980-011-9639-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-011-9639-6