
Abstract
We used two sediment cores from the ancient harbour of Rome (Italy) for palaeoenvironmental reconstruction. The imperial harbour of Rome and its town, Portus, were constructed in the Tiber Delta area. Today, they are ~3 km from the coast, close to Leonardo da Vinci International Airport. The port was excavated during the reign of Emperor Claudius, inaugurated by Nero in 64 AD, and substantially enlarged under Emperor Trajan. This paper focuses on analyses of ostracods in sediment cores from the harbour and provides insights into the palaeoenvironmental evolution of the harbour water bodies. Pollen data complement this ostracod-based reconstruction, providing information about the vegetation around the port. One core (PTS13) was taken from the dock area (darsena) excavated at the time of Claudius. The second core (PTS5) was collected from the Trasverso Channel (Canale Trasverso), excavated after the port inauguration and later dredged during the fourth century AD because it had filled in with silt. Radiocarbon, archaeological and pollen data indicate the two cores overlap in time slightly, or perhaps not at all, the core from the dock site (PTS13) containing the older record. Ostracods recovered from the two sites represent very different aquatic environments, driven in one case by the Tiber River input and in the other by “marine” harbour waters. Parallel study of ostracods and pollen in the cores reveals the impact of harbour management activities, which are linked to ship hull maintenance, as well as erosion and siltation in this coastal setting.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Archaeological questions in the Mediterranean region have been resolved recently through palaeoenvironmental investigation of ancient harbour basins (Marriner et al. 2010). Interpretation of such peculiar sediment archives, profoundly affected by both human impacts and natural processes, requires an interdisciplinary approach. Synergy among archaeological, sedimentological and paleontological data can yield valuable insights.
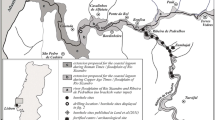
The history of the large harbour built during imperial times at Portus, just south of Rome, is closely related to the evolution and morphology of the Tiber Delta. The Soprintendenza Speciale per i Beni Archeologici di Roma (Superintendence for the Archaeological Heritage of Rome) obtained a suite of deep sediment cores in the area with the aim of reconstructing the palaeoenvironmental evolution of the harbour and the Tiber Delta during Roman times.
This study focuses on two cores drilled in the Claudius harbour area (Fig. 1). The cores were studied previously for stratigraphy (Giraudi et al. 2009) and pollen (Sadori et al. 2010a, b). This study used ostracod community analysis to make inferences about the palaeoenvironment of the Portus area, from the time before the harbour dredging until abandonment of the area.

Satellite image of the modern location of Portus in the Tiber Delta area showing locations where sediment cores PTS5 and PTS13 (white) and core TRXIX (black) (from Goiran et al. 2010) were collected
Historical background
In 42 AD, Emperor Claudius decided to build a new harbour along the Latium coast, near the mouth of the Tiber River. Many factors were considered in deciding on the harbour location: (1) it was difficult to reach Rome by ship due to currents in the mouth of the Tiber River (Strabo, Geografia 5.3.5), (2) there had been recent food shortages in Rome (Suetonius, Vita Claudii 18, in De Vita Caesarum), Claudius needed to undertake popular projects, and there were obvious benefits to following in the footsteps of popular figures from the past, such as Julius Caesar (Suetonius, Vita Claudii 20 in De Vita Caesarum). Moreover, it was thought that a new canal, linking the harbour to the Tiber River, would reduce flooding in Rome by enabling excess waters to flow quickly to the sea. Reference to flood mitigation is contained in an inscription from 46 AD. Indeed, it has been suggested that Tiber River flood frequency was high at the beginning of the first century AD and during the following century (Camuffo and Enzi 1995; Bersani and Bencivenga 2001). Catastrophic events are also reported for a harbour along the Arno River (Benvenuti et al. 2006; Mariotti Lippi et al. 2006), where high-magnitude floods occurred between the second century BC and the fifth century AD.
The building site for the new harbour of Rome was chosen carefully, to be close to the city and to the Tiber River, making it easy for commodities to reach the town via this waterway. The harbour was excavated on the bank of the Tiber River where a coastal lagoon opened to the sea, and it was protected by a beach ridge to the west (Giraudi 2004). Claudius’s project included excavation of a channel, the Fossa Traiana, today’s Canale di Fiumicino, to connect the harbour to the Tiber River. Claudius did not survive to see the completion of this ambitious project and it was finished in 64 AD, under Emperor Nero. The harbour was built to accommodate both merchant and military ships. Warehouses and granaries were built along its shores to store abundant commodities that arrived from the provinces. In the early second century AD, Claudius’s harbour at Portus was enlarged in response to the need for increased shipping, the problem of siltation, and the exposure of moles, i.e. massive stone works emplaced to provide protection from waves. The harbour was modified by Emperor Trajan to have a protected hexagonal inner basin (Juvenal, Satura 12.74–82). Several years later, during the early Flavian period, after 69 AD, the Canale Trasverso was completed to connect to the enlarged Fossa Traiana, which reached the Tiber River. In 533, Procopius (De Bello Gothico, 1, 26, 7–13) stated that the Trajan harbour was in use and that the connection of the Fossa Traiana with the river (Capo Due Rami) was 2,675 m (15 stadi) from the coastline. Between the fourth and the sixth century AD, Portus underwent gradual decline, probably as a consequence of increased barbarian raids during the Gothic Wars. Moreover, malaria was already well established in the large coastal lagoons during the fifth century AD (Sallares 2006). During the early Medieval Period, the area was abandoned. Only the Bishop’s Palace (Episcopio di Porto) survived, and can still be seen today along the Fossa Traiana. During the eleventh century AD, St. Peter Damian, Bishop of Ostia, wrote about the unhealthiness of the area, alluding to the presence of malaria (Sallares 2006). In a Vatican Museum fresco, dated to 1582, Claudius harbour is represented as being dry, whereas the Trajan hexagonal basin, together with the dock, forms a continuous water body and remnants of ancillary channels still contained some water. The area of Portus and the abandoned ruins were gradually buried and remained so until archaeological excavations began in the nineteenth and twentieth centuries.
Geological and morphological background
The Tiber River drains a basin that goes from the central Apennines to the Tyrrhenian Sea. Over its 406-km length, it erodes limestones, flysch, siliciclastic and volcanic sediments. The Tiber Delta possesses an exposed area that covers about 150 km2 and its underwater extent is ~500 km2, with a maximum sediment thickness of 80 m (Bellotti et al. 2007). The Late Quaternary stratigraphy of the area has been the subject of several multidisciplinary studies, as development of the delta involved complex interactions among sediment discharge from the Tiber River, postglacial sea level rise and tectonic uplift (Bellotti et al. 1994, 2007; Giordano et al. 2003).
The current morphology of the coastal plain is the result of landscape evolution that took place during the last 5,000 years (Rendell et al. 2007; Giraudi et al. 2009). Once sea level stabilised, sediment input was mainly linked to solids discharged by the Tiber River itself. In recent times, after the building of several dams upstream of Rome, the river sediment load dropped from about 12 × 109 to 1 × 109 kg/year (Bellotti et al. 2007). Portus is now located within the upper delta plain (Fig. 1). In the Portus area, sediment cores studied by Bellotti et al. (2007, 2009), Giraudi et al. (2009) and Goiran et al. (2010) represent material from two different depositional systems.
The pre-Roman age of delta sediments at the base of the cores was obtained by radiocarbon dating macrofossils. The upper sediments have an age consistent with the time of harbour construction and subsequent infilling phases. Following Giraudi et al. (2009), the lower portion of the drill holes presented here has a delta origin. These old deposits display coarser particles with lighter colour than the sediments associated with the later harbour filling. Wood fragments, ceramic sherds and ornamental stones often occur in the harbour infill.
Materials and methods
The analysed cores are part of a wider drilling campaign carried out by the Soprintendenza Speciale per i Beni Archeologici di Roma. The PTS13 core was taken in the southern part of Claudius harbour, in front of the entrance to the dock area (darsena) (Fig. 1). It was drilled at 0.19 m below sea level and is 9.5 m long, allowing a general view of the sediment that filled the area. The PTS5 core was drilled at 2.12 m a.s.l. to correspond with the elevation of the Canale Trasverso, the channel that connects the harbour to the Fossa Traiana, one of the channels of the Tiber (Fig. 1). Core PTS5 is 12.50 m long and contains a record of the fluvial influence on the harbour waters.
Ostracod analyses were carried out on both cores. Nineteen samples from core PTS13 were analyzed (Fig. 2), as were 27 samples from core PTS5 (Fig. 2). Each sample was disaggregated in 5% H2O2, washed on a 0.63-mm mesh and dried. When possible, at least 300 ostracod valves/sample were handpicked under the stereomicroscope (Nikon SMZ-U 15-150x) and identified. Each species frequency was normalised to 10 g of dried sieved sample and the percent relative abundance of each taxon was determined. Species represented only by juveniles or by single adult valves with occurrences <10, were considered displaced valves (C population structure sensu Whatley 1988; allochthonous low-energy taphocoenosis sensu Boomer et al. 2003). Interpretation of the assemblages used autoecological data from the literature.

Lithology of sediment cores PTS5 and PTS13
Pollen and micro-charcoal analyses were also carried out on both cores (Sadori et al. 2010a). A total of 21 sediment samples from PTS5 core and of 15 samples from PTS13 were chemically processed with strong acids and base, following standard procedures. The pollen sum includes all terrestrial taxa. The sum used for algae percentages was obtained by adding them to the basic pollen sum. Selected palynological curves were plotted against ostracod data to better understand characteristics of the aquatic environment.
Results
Ostracod assemblages
Ostracods were well preserved, mainly as loose valves, as opposed to whole carapaces. Several samples yielded few juvenile valves, whereas most samples were characterised by a mixture of remains from adults and immature ostracods. The ostracod assemblage of core PTS13 is very diverse, with 2,581 valves recovered and 72 taxa recognised (Fig. 3a). At the base of the core (6.76–6.35 m), the ostracod fauna is scarce or represented predominantly by C. torosa, indicating a brackish environment. In the subsequent samples (6.15–5.07 m) the marine signal becomes increasingly evident, with an association of typical coastal species such as Carinocythereis carinata, Cytheridea neapolitana, Xestoleberis communis, Xestoleberis dispar, Pontocythere turbida (Fig. 4n), Palmoconcha turbida (Fig. 4q), seven species of Semicytherura and five species of Leptocythere (Appendix A, see Electronic Supplementary Material). Cyprideis torosa is present throughout the section (Fig. 4c). Pontocythere turbida and Palmoconcha turbida dominate at depths between 6.15 and 5.00 m. Most of the marine taxa are phytal species that require algae or higher plants for their life cycle (Loxoconcha spp., Aurila spp.). Many of them are linked to the presence of Neptune grass (Posidonia oceanica) meadows, i.e. Mediterranean tapeweed (Lachenal 1989). The environment was truly marine, shallow and vegetated, with some influence from the nearby river and/or swamps, as seen in the low occurrence of C. torosa and some juvenile specimens of Candoninae species. The sample recovered at 5.00–4.93 m depth was barren of ostracods, whereas the overlying sample (4.59–4.52 m) displays a truly marine association, though less rich in numbers and less diverse in taxa. Most likely, the waterbody was under some sort of stress. The next samples (4.34–3.90 m) are characterised by a very sparse marine fauna, dominated by juvenile specimens. At 2.56 m, a sudden change is recorded. C. torosa becomes dominant, Heterocypris salina is also present and the occurrence of marine species is sporadic until 1.70 m depth. The uppermost two samples (1.16 and 0.90 m) are characterised by a well-balanced, low oligohaline assemblage with the coexisting dominants C. torosa, Candona neglecta (Fig. 4a) and Ilyocypris gibba.

Frequencies of the dominant ostracod species in cores a PTS5 and b PTS13

SEM pictures of selected ostracod species: a Candona neglecta, RV, PTS13 (sample −1.11/−1.16); b Ilyocypris monstrifica, LV, PTS5 (sample −6.10/−6.15); c Cyprideis torosa, RV, PTS13 (sample −1.11/−1.16); d Leptocythere ramosa, LV, PTS13 (sample −5.64/−5.69); e Loxoconcha littoralis, LV, PTS13 (sample −5.64/−5.69); f Semicytherura sulcata LV (sample −4.28/−4.34); g Hemicytherura defiorei LV (sample −4.08/−4.15); h Semicytherura dispar, LV, PTS13 (sample −5.64/−5.69); i Procytherideis subspiralis, LV, PTS13 (sample −5.07/−5.13); j Callistocythere flavidofusca, RV, PTS13 (sample −5.64/−5.69); k Costa batei, RV, PTS13 (sample −6.10/−6.15); l Pontocythere turbida, LV, PTS13 (sample −6.10/−6.15); m Loxoconcha rhomboidea, RV, PTS13 (sample −5.07/−5.13); n Neocytherideis subulata, RV, PTS13 (sample −5.07/−5.13); o Palmoconcha turbida, LV, PTS13 (sample −5.07/−5.13). (scale bars = 100 μm)
In core PTS5, a total of 3,809 valves were recovered and 26 taxa were identified (Pl. 1, Fig. 3b). The ostracod assemblage is not diverse, with dominance of C. torosa in association with Heterocypris salina, from the base of the core to a depth of 3.71 m. The opportunistic species Cyprideis torosa occurs in a wide range of salinities from nearly fresh to hypersaline waters, with an optimal salinity of about 2–16.5‰. It is a typical inhabitant of coastal ponds, lakes, lagoons, estuaries, fjords, deltas and salt marshes (Meisch 2000).
At the base of the core (12.22–7.91 m) the ostracod fauna is poorly represented, mostly by C. torosa. The presence of Cytheridea neapolitana and Pontocythere turbida at the base indicates shallow marine conditions, whereas the occurrence of H. salina and C. neglecta higher in the core indicates lower salinities (low mesohaline). From 7.79 to 7.31 m, the combined presence of C. neapolitana and Pontocythere turbida, along with the striking dominance of C. torosa is again indicative of shallow marine waters. From 6.99 to 6.11 m C. torosa occurs together with H. salina and Sarsicypridopsis aculeata, pointing to a less saline environment. From 5.79 to 5.11 m, the association is richer in shallow marine components, pointing to a marginal marine environment. From 4.99 to 4.15 m, salinity declines again and the marine group is gradually replaced by brackish-water taxa. From 3.79 to 1.56 m, the environment changes dramatically. C. torosa almost disappears and the Candoninae assemblage is dominant. Pseudocandona marchica and C. neglecta represent taxa that live in freshwater to low oligohaline salinities.
Ostracod autoecology
Four main ecological groups of ostracods were distinguished using the ecological tolerances of the autochthonous species, represented by both juveniles and adults (Fig. 3). As this is a marginal marine environment, the boundary between these four groups is not sharp, but is determined by the dominant species (Horne and Boomer 2000; Meisch 2000; Anadón et al. 2002; Ruiz et al. 2003; Cabral et al. 2006):
-
1.
Marine coastal: dominated by C. carinata, C. neapolitana, Bairdia spp., and Semicytherura spp. (Fig. 4f, h). Most species are known to live in P. oceanica meadows, P. oceanica detritus or in algae (Lachenal 1989).
-
2.
Marine lagoonal: dominated by Pontocythere turbida (Fig. 4n) and Palmoconcha turbida (Fig. 4q), with accompanying Loxoconchidae, Xestoleberididae and C. torosa. These species are associated with all transitional environments between marine and fresh waters, and are characterised by high tolerance to salinity variations and trophic gradients (Carbonel 1982).
-
3.
Brackish lagoonal: dominated by C. torosa (mainly smooth forms, Fig. 4c), with accompanying H. salina. C. torosa has its greatest development within the salinity range 2–16.5‰, but it can survive up to 60‰. Smooth forms of C. torosa occur at salinities >5‰ (Meisch 2000). H. salina can tolerate salinities up to 20‰ (Meisch 2000).
-
4.
Fresh water: the assemblage is characterised by Candonidae (C. neglecta, S. aculeata, P. marchica, P. albicans), I. gibba, I. monstrifica (Fig. 4 b), Herpetocypris sp. and C. vidua. C. neglecta tolerates salinities up to 15‰, typical of swampy areas, but dominates in the two cores when C. torosa is absent or represented only by juveniles. C. vidua tolerates only slight increases of salinity. The occurrence in one sample of a single species (P. marchica) could point to an isolated event of stagnant or slowly running waters.
Gypsum occurrence
During analyses of the washed residues, microscopic crystal rosettes were found in some intervals of core PTS13, from the dock area (Fig. 3). Samples were analysed using XRD and were shown to be composed of gypsum. The gypsum rosettes are concentrated in four samples (5.92–5.86, 5.13–5.07, 2.56–2.50, 2.36–2.30 m). Surprisingly, a diverse ostracod fauna co-occurs with gypsum in the two lowermost samples, though the assemblages are dominated by juvenile forms. In both samples, the dominant species is Palmoconcha turbida, an opportunistic shallow marine species, whose tolerance to low oxygen levels allows its survival in hypoxic conditions (Bodergat et al. 1998). The two uppermost samples are characterised by a sparse assemblage that is dominated by C. torosa. C. torosa occurs in facies containing gypsum and halite, indicating hypersaline conditions, most likely very shallow lagoons that underwent desiccation and developed sabkha (salt flat) conditions (Pugliese and Stanley 1991). Masson (1955) found similar gypsum crystals in the Laguna Madre, in southwest Texas, and concluded that the gypsum precipitated from highly saline seawater, which periodically moves in windblown sheets across the surface of the mudflats and sinks into the underlying sediments. The occurrence of gypsum rosette crystals is indicative of very warm, continental sediments containing a high concentration of plant humic compounds (Cody and Cody 1988). In experimental conditions, gypsum rosettes mostly occur at high tannic acid concentrations and moderate salinities and are most widespread at high temperature (Cody and Cody 1988).
Discussion
The two bottom samples in the PTS13 core (6.70–6.10 m depth) are characterised by an association dominated by C. torosa, with C. neglecta and P. marchica as subordinate species. The occurrence of this latter species sets a salinity limit for the water body at a maximum of 5.5‰ (Meisch 2000). The six samples subsequent to the foundation of the harbour (5.92–4.52 m depth) bear a rich and diverse marine assemblage, with a total of 36 species. Here, Palmoconcha turbida and Pontocythere turbida dominate, with C. carinata, C. neapolitana, Leptocythere spp. and Semicytherura spp. as the most abundant accompanying species. At 4.34 m depth, a sudden change in salinity occurs. Marine species become scarce and are represented mostly by juvenile specimens.
The pollen record starts at 5.70 m, with the port-filling sediments (Fig. 5). Only selected taxa, whose presence could be related to the aquatic environment, and thus to ostracod occurrences, are shown in the figure. Amaranthaceae, herbs of possible brackish environments, are very close to 50%, in agreement with the high values for marine ostracods. Tamarix (tamarisk) trees, which are resistant to the saline water table, nevertheless display very low percentages. A peak of algae at 4.90 m matches the highest occurrence of freshwater ostracods. Algal remains are mostly ascribed to Botryococcus. Most species belonging to this genus are members of the freshwater phytoplankton, though some tolerate water salinities up to 8‰. These colonial algae are commonly considered to be indicators of river inflow (Mudie et al. 2011). The Botryococcus peaks are followed by a slight increase (>5%) of Alnus, a freshwater riparian tree, and more importantly, by an expansion of Tamarix (>21%), matching the last expansion of marine ostracods.

Stratigraphic comparison between selected ostracod groups and pollen taxa in cores PTS5 and PTS13
Occurrence of timber remains, specifically a 23-cm-thick piece of deciduous Quercus and a 22-cm-thick piece of Ulmus (Sadori et al. 2010a), between 3.60 and 2.90 m depth, marks a sudden change in salinity. In the following samples C. torosa and H. salina become the dominant species and their occurrences outnumber those of the less abundant marine species. The pollen record, which mainly provides information on the terrestrial environment, indicates an increase in riparian freshwater trees like Alnus (alder) above the occurrence of timber. This trend towards freshwater conditions is in agreement with the ostracod record, indicating a shallow and brackish marshy environment above the timber stratum. In the uppermost pollen sample, an Amaranthaceae peak suggests an increase in soil salinity. This salinity trend, detected also from ostracods, continues in the topmost samples, which correspond to the period of harbour abandonment. The ostracods record a permanent water body, probably linked to the presence of a slightly salty bog.
The ostracod assemblage reported by Goiran et al. (2010) from a core just outside the dock area (core TRXIX) highlights the complex environment of an artificial harbour. Of the 26 species reported from TRXIX, 20 are shared with PTS13. What differs between the cores is the species dominance. In core TRXIX, typical coastal marine species (Pontocythere elongata, C. carinata, Loxoconcha lisse) coexist with freshwater (Candona spp.) and brackish (C. torosa) species. Occurrence of these two latter taxa is likely linked to the influence of the Tiber River waters. Such influences are negligible in core CL3 (Goiran et al. 2010), located south of the northern pier. It contains a typical coastal marine association.
Occurrence of gypsum crystals in the PTS13 core gives rise to the hypothesis that the dock area was dredged or underwent partial desiccation several times. It may be possible to distinguish between these two hypotheses by observing microfossil differences between the gypsum-bearing samples. The lowermost sample (5.92–5.86 m) is characterised by a low-diversity marine assemblage with abundant juvenile specimens. It could correspond to the initial phase of the harbour, just after it was founded, when the water level was shallow, allowing construction of the walls bounding the dock. The second sample with gypsum crystals (5.13–5.07 m) is associated with the most abundant and diverse marine assemblage recorded in the core. There, gypsum deposition could have occurred in a shallow-water environment rich in plant debris, as confirmed by the occurrence of abundant phytal taxa that could have been transported to the dock area along with Posidonia oceanica debris. The last two samples with gypsum occur just below and above the wood remains, respectively, suggesting that the wood could have belonged to a ship hull that was being repaired or abandoned at the dock, in very shallow and muddy waters.
The PTS5 core reflects a complex setting, with influences of freshwater detected in the ostracod assemblages. The bottom of the PTS5 core dates to a time before 3,560–3,380 cal yr BP (Giraudi et al. 2009). The three bottom samples (12.22–9.12 m) are dominated by scarce C. torosa and H. salina, with reworked valves of marine ostracods. Such samples are related to the deposition of sands and silty sands in the Tiber Delta, and pertain to the “beach, dune and fluvial mouth bar sand association” identified in other drill cores from the area (Bellotti et al. 2007). In the subsequent samples, the palaeoenvironment evidently changes dramatically. C. torosa dominates the assemblage from 8.79 m to 3.31 m, and some marine taxa are always present and represented by complete populations, i.e. adults and juveniles, of Pontocythere turbida, C. whitei and Loxoconcha spp. Such a change could be attributed to the dredging of the harbour that occurred during the fourth century AD (Paroli 2004), which allowed water exchange between the Tiber River and the harbour. In this interval, other marine species are represented by juvenile valves of Bairdia sp., Cytheretta subradiosa, Xestoleberis spp., Urocythereis sp., and Tyrrhenocythere amnicola. These ostracods could have been transported from the harbour basin and the open sea by a combination of waves, tidal action and storms (Boomer and Eisenhauer 2002), or through transport on ship hulls.
The pollen record is pertinent to the “Canale Trasverso” sediments. It shows high values (>55%) of Tamarix, opposite to the Amaranthaceae curve (Fig. 5). The contrast between the relative abundances of “indicator” taxa yields clear inferences. Riparian freshwater tree pollen (Alnus) shows very low percentages, and displays no relation with saline-resistant taxa like Tamarix. Comparing the tamarisk curve with the marine ostracod record, the first two expansions of Tamarix contrast with the brackish-water conditions recorded by the ostracods. A single correspondence occurs around 5.50 m, where the highest peaks of the tree pollen are accompanied by dominance of the marine crustaceans.
The occurrence of high percentages of tamarisk pollen, in contrast with the ostracod data supporting brackish conditions, was considered evidence of strong human interference in the harbour area (Sadori et al. 2010a). It was thought that such trees were planted to prevent erosion and protect the inner zones from strong marine winds. Later correspondence between the pollen and ostracods suggests that natural conditions were restored, once the area was abandoned.
The only large peak of algal remains (Botryococcaceae) is found in association with an Alnus decrease and the less saline interval recorded by the ostracods in the “Canale Trasverso” infill. At 2.99–2.91, only one species, P. marchica, occurs, represented both by juveniles and adults. This species lives in temporary and permanent water bodies, in the benthic and interstitial zones of rivers, in brackish coastal and inland waters, with salinities up to 5.5‰. Bloom events in shallow temporary pools have been recorded in Northern Europe (Meisch 2000). The occurrence of such an assemblage could suggest a similar bloom event in a shallow temporary pool, and that the channel was no longer in use. Uppermost samples are characterised by C. neglecta and scarce valves of C. torosa. C. neglecta lives in temporary and permanent water bodies and is common in coastal waters and brackish inland waters up to 16‰ (Meisch 2000).
Stratigraphic and chronologic correlation between cores is challenging. In PTS13, two radiocarbon dates were provided by Sadori et al. (2010a): (1) 1805 ± 40 years BP (calibrated age, 2 σ interval: 85–340 AD, corresponding to ca. 212 ± 128 AD) on the wood remains occurring between 3.23 and 3.17 m, and (2) 1,780 ± 25 (calibrated age, 2 σ interval: 130–340 AD, corresponding to 235 ± 105 AD) on pollen extracted from the organic-matter-rich level at 1.75 m. PTS5 could not be dated directly. Its sediments represent a time frame bounded between the date of dredging of the Canale Trasverso (fourth century AD) and that of the top sediments (Renaissance).
Comparison with the cores studied by Goiran et al. (2010) allows the following inferences: freshwater input increased from west to east, and the harbour foundation surface is shallower in the dock area. Marginal marine ostracod assemblages often include allochthonous specimens from freshwater or open marine sources, and passive introduction by waterbirds cannot be excluded (Boomer and Eisenhauer 2002). In settings such as Portus, human activities like dredging and transport on ship hulls cannot be excluded. In any case, analyses of the ostracod communities recovered from the two sediment cores record three main changes in the water body, occurring at different times (Fig. 6). At the bottom, both cores are characterised by deltaic sediments and the ostracod fauna is scarce and represented mainly by C. torosa, a colonizing species that tolerates sandy bottoms typical of estuaries, salt marshes and deltas (Meisch 2000). A sudden change in ostracod assemblages marks different moments in each of the cores. In PTS13, the transition to a typical coastal marine assemblage indicates phases after construction of the harbour, when it was open to the sea and the marine microfauna could easily colonize the new environment. In PTS5, the change in the ostracod assemblage indicates the boundary between the Tiber Delta sediments deposited before the construction of the harbour and the base of the Trasverso Channel deposits. Waters of the Tiber River could flow through this channel and mix with the more saline waters of the harbour, as also indicated by the sporadic occurrence of Botryococcaceae. The last phase, probably corresponding to the Renaissance period, corresponds in both cores to a marshy environment that developed after abandonment of the harbour. In PTS5 only, there is a subsequent phase characterised by fluvial sands and a sparse, reworked ostracod fauna, probably pointing to the reestablishment of a fluvial environment. Sadori et al. (2010a, b) provided evidence from PTS5, which contains the youngest deposits, for intensive land use. Such land modification was likely related to the development of Portus, the harbour town.

Correlation of the cores and palaeoenvironmental interpretations
Conclusions
Ostracod assemblages in two sediment cores recovered from the harbour of Portus, Italy, record different environmental conditions, which can be related to two different chronological periods on the basis of radiocarbon, archaeological, and historical data. In core PTS13, gypsum crystals in the interval 5.92–2.60 m (at present ~2.80 m b.s.l.), indicate harbour management activities such as partial drainage of the basin. In PTS5 the ostracod assemblage records abrupt salinity variations that are also related to harbour management activities.
Comparison of ostracod and pollen data allowed us to discriminate between natural and anthropic environmental changes. Because ostracods are sensitive to changes in ambient water chemistry, we focused on pollen of taxa that could be linked to waters of different salinity. Good agreement was found between Amaranthaceae and ostracod percentages. Salt-tolerant plants at first expanded when ostracods record brackish conditions. Pollen peaks are therefore related to species (e.g. Salicornia and Sarcocornia) that live in salty soils.
Botryococcaceae peaks, likely related to increases in river input, are never found in intervals, identified by other lines of evidence, to be marine. These algae thus correspond to episodes of low salinity in the harbour. Lack of correspondence between most peaks of Tamarix and peaks of marine/brackish ostracods is attributed to strong human influence, e.g. tree planting, associated with harbour management. In coastal settings, where human impacts can overwhelm natural processes, interdisciplinary studies are required to distinguish between natural and anthropic signals in the palaeoenvironmental record.
References
Anadón P, Gliozzi E, Mazzini I (2002) Paleoenvironmental reconstruction of marginal marine environments from combined paleoecological and geochemical analyses on ostracods. In: Holmes AH, Chivas AR (eds) The ostracoda: applications to Quaternary research. AGU Geophys Monogr, vol 131, pp 227–247
Bellotti P, Chiocci FL, Milli S, Tortora P, Valeri P (1994) Sequence stratigraphy and depositional setting of the Tiber delta: integration of high resolution seismics, well logs and archaeological data. J Sediment Petrol B64:416–432
Bellotti P, Calderoni G, Carboni MG, Di Bella L, Tortora P, Valeri P, Zernitskaya V (2007) Late Quaternary landscape evolution of the Tiber River delta plain (Central Italy): new evidence from pollen data, biostratigraphy and 14 C dating. Z Geomorph NF 4:505–534
Bellotti P, Mattei M, Tortora P, Valeri P (2009) Geo-archaeological investigations in the area of the imperial harbours of Rome. Méditerranée 112:51–58
Benvenuti M, Mariotti Lippi M, Pallecchi P, Sagri M (2006) Late Holocene floods in the terminal Arno river (Pisa, Central Italy): hydro-climatic catastrophic events and short-term sealevel changes from the history of the Roman riverine harbour. Holocene 16:863–876
Bersani P, Bencivenga M (2001) Le piene del Tevere a Roma dal V secolo a.C. all’anno 2000. In: Servizio idrografico e mareografico nazionale. Presidenza del Consiglio dei Ministri. Dipartimento per i Servizi Tecnici Nazionali, Roma pp 1–100
Bodergat AM, Ikeya N, Irzi Z (1998) Domestic and industrial pollution: use of ostracods (Crustacea) as sentinels in the marine coastal environments. J Rech Oceanogr 23:139–144
Boomer I, Eisenhauer G (2002) Ostracod faunas as palaeoenvironmental indicators in marginal marine environments. In: Holmes AH, Chivas AR (Eds) The ostracoda: applications to Quaternary research. AGU Geophys Monogr, vol 131, pp 135–149
Boomer I, Horne DJ, Slipper IJ (2003) The use of ostracods in palaeoenvironmental studies, or what can you do with an ostracod shell? In: Park LE, Smith AJ (Eds) Bridging the gap. Trends in the ostracode biological and geological sciences. Paleontol Soc paper, vol 9, pp 153–180
Cabral MC, Freitas MC, Andrade C, Cruces A (2006) Coastal evolution and Holocene ostracods in Melides lagoon (SW Portugal). Mar Micr 60:181–204
Camuffo D, Enzi S (1995) The analysis of two bi-millenary series: Tiber and Po river floods. In: Jones PD, Bradley R, Jouzel J (eds) Climatic variations and forcing mechanism of the last 2000 years. NATO-ANSI series, 1: global environmental change, vol 41. Springer, Berlin, pp 433–445
Carbonel P (1982) Les ostracodes, traceurs des variations dans des systèmes de transition eaux douceseaux salées. Mémoire de la Société géologique de France 144:17–128
Cody RD, Cody AM (1988) Gypsum nucleation and crystal morphology in Analog saline terrestrial environments. J Sediment Res 58:247–255
Giordano G, Esposito A, De Rita D, Fabbri M, Mazzini I, Trigari A, Rosa C, Funiciello R (2003) The sedimentation along the roman coast between middle and upper Pleistocene: the interplay of eustatism, tectonics and volcanism—new data and review. Il Quaternario 16:121–129
Giraudi C (2004) Evoluzione tardo-olocenica del delta del Tevere. Il Quaternario 17:477–482
Giraudi C, Tata C, Paroli L (2009) Late Holocene evolution of Tiber river delta and geoarchaeology of Claudius and Trajan Harbor, Rome. Geoarchaeology 24:371–382
Goiran JP, Tronchère H, Salomon F, Carbonel P, Djerbi H, Ognard C (2010) Palaeoenvironmental reconstruction of the ancient harbors of Rome: Claudius and Trajan’s marine harbors on the Tiber delta. Quatern Int 216:3–13
Horne D, Boomer I (2000) The role of ostracoda in saltmarsh meiofaunal communities. In: Sherwood BR, Gardiner BG, Harris T (eds) British saltmarshes. Forrest Text, Cardigan, for the Linnean Society of London, London, pp 181–202
Lachenal AM (1989) Écologie des ostracodes du domaine méditerranéen. Application au Golfe du Gabès (Tunisie orientale): les variations du niveau marin depuis 30000 ans. Documents des Laboratoires de Géologie de Lyon 108:1–239
Mariotti Lippi M, Bellini C, Trinci C, Benvenuti M, Pallecchi P, Sagri M (2006) Pollen analysis of the ship site of Pisa San Rossore, Tuscany, Italy: the implications for catastrophic hydrological events and climatic change during the late Holocene. Veget Hist Archaeobot 16:453–465
Marriner N, Morhange C, Goiran JP (2010) Coastal and ancient harbour geoarchaeology. Geol Today 26:21–27
Masson PH (1955) An occurrence of gypsum in southwest Texas. J Sediment Petrol 25:72–77
Meisch C (2000) Freshwater ostracoda of Western and Central Europe. In: Schwoerbel J, Zwick P (eds) Süsswasserfauna von Mitteleuropa 8/3. Spektrum Akademischer Verlag, Heidelberg, pp 1–522
Mudie PJ, Leroy SAG, Marret F, Gerasimenko N, Kholeif SEA, Sapelko T, Filipova-Marinova M (2011) Nonpollen palynomorphs: indicators of salinity and environmental change in the Caspian–Black Sea–Mediterranean corridor. Geol Soc Am Spec Pap 473:1–27
Paroli L (2004) Il porto di Roma nella tarda antichità. II seminario ANSER, Roma-Ostia Antica 16–17 aprile 2004. In: Gallina Zevi A, Turchetti R (eds) Le strutture dei porti e degli approdi antichi. Rubbettino Editore, Cosenza, pp 247–266
Pugliese N, Stanley DJ (1991) Ostracoda, depositional environments and late Quaternary evolution of eastern Nile Delta, Egypt. Il Quaternario 4:275–302
Rendell HM, Claridge AJ, Clarke ML (2007) Late Holocene Mediterranean coastal change along the Tiber Delta and Roman occupation of the Laurentine shore, central Italy. Quat Geochron 2:83–88
Ruiz F, González-Regalado ML, Muñoz JM, Pendón JG, Rodríguez-Ramírez A, Cáceres L, Rodríguez-Vidal J (2003) Population age structure techniques and ostracods: applications in coastal hydrodynamics and paleoenvironmental analysis. Palaeogeogr Palaeoclimatol Palaeoecol 199:51–69
Sadori L, Giardini M, Giraudi C, Mazzini I (2010a) The plant landscape of the imperial harbour of Rome. J Archaeol Sci 37:3294–3305
Sadori L, Mercuri AM, Mariotti Lippi M (2010b) Reconstructing past cultural landscape and human impact using pollen and plant macroremains. Plant Biosyst 144:940–951
Sallares R (2006) Role of environmental changes in the spread of malaria in Europe during the Holocene. Quatern Int 150:21–27
Whatley RC (1988) Population structure of ostracods: some general principles for the recognition of palaeoenvironments. In: De Deckker P, Colin J-P, Peypouquet J-P (eds) Ostracoda in the earth sciences. Elsevier, Amsterdam, pp 245–256
Acknowledgments
We thank Lidia Paroli from Soprintendenza Speciale per i Beni Archeologici di Roma who kindly provided the cores drilled in the archaeological area. We acknowledge the help of Dr. Sergio Lo Mastro (Roma 3 University) for XRD analyses of the gypsum crystals and useful discussions. The work was partly financed by the Italian PRIN (2008FJCEF4) project PICAR (Cultural landscape and human impact in circum-Mediterranean countries). The comments and suggestions of two anonymous referees and of Dr. Mark Brenner (University of Florida) considerably improved the manuscript.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Mazzini, I., Faranda, C., Giardini, M. et al. Late Holocene palaeoenvironmental evolution of the Roman harbour of Portus, Italy. J Paleolimnol 46, 243–256 (2011). https://doi.org/10.1007/s10933-011-9536-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10933-011-9536-7