
Abstract
Three sediment cores from neighbouring estuaries along the Portuguese Algarve coast were analysed for the comparison of grain-size distribution, geochemical composition and microfossil associations. The goal of this study is to reconstruct the environmental evolution of the estuaries. Our reconstructions based on foraminifera and ostracods reveal several distinct chronological stages, including the initial situation of the palaeo-valleys, the ingression of the sea into the river mouths at about ~7500-8000 cal BP in the Quarteira and São Lourenço estuaries and ~8000 cal BP in the Carcavai estuary, and the silting-up following the sea-level high stand after ~6500 cal BP until the present.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
An interdisciplinary project aimed to reconstruct regional vegetation change and coastal evolution in response to climatic and anthropogenic impacts on sedimentation processes along the Algarve coast was initiated in 2000. The areas of investigation are the estuaries of Alvor, Alcantarilha, Quarteira, Carcavai and São Lourenço. Studies on the sedimentology, geochemistry and palynology were performed for comprehensive information on the development of the first four estuaries. Until now, scientific studies on the genesis of these coastal areas have been lacking, except for those by Boski et al. (2008), Andrade et al. (2004), Hilbich et al. (2008), Schneider et al. (2010) and Trog et al. (2013). The present work investigates the evolution of three estuaries using a multi-proxy approach and represents a first regional study on the Holocene landscape development of the river valley of the Ribeira de São Lourenço. Our goal was to reconstruct the landscape evolution of the estuaries of the Ribeira de Quarteira, Ribeira de Carcavai and Ribeira de São Lourenço for the last ~8000 years. The depositional conditions of the lagoonal sediments were characterised using sedimentological, geochemical and microfaunal analyses of the sediment cores.
Micropalaeontological analyses are a focus of this work contributing to the microfaunal interpretation of the Holocene evolution along the Portuguese Algarve coast. Previous studies have considered the distribution of foraminifers or ostracods to reconstruct the genesis of Iberian coastal areas. Along the south-western and southern coast, the investigations of Alday et al. (2013) at the Albufeira lagoon, Cabral et al. (2006) and Cearreta et al. (2007) at the Melides lagoon, Cearreta et al. (2003) at the Santo André, Alday et al. (2006) and Loureiro et al. (2009) at the Mira river estuary, Ruiz Muñoz et al. (1996) and Mendes et al. (2010) at the Guadiana river mouth, Ruiz et al. (2004) and Pozo et al. (2010) at the Doñana National Park in southwest Spain and Blázquez and Usera (2010) in Alicante can be highlighted.
Regional settings
The Algarve coast is a graded shore line, which is largely dominated by cliffs which are occasionally interrupted by flat, sandy coastal areas.
The study areas of the Quarteira and Carcavai estuaries are located 20 and ~13 km, respectively, to the west of Faro on the south coast of Portugal (Fig. 1a). The estuary of the Ribeira de Quarteira (Fig. 1b) has a length of 5 km and a maximum width of 2 km and has a catchment area of 405 km2 that extends into the Serra do Caldeirão (Schneider et al. 2010). The estuary of the Ribeira de Carcavai (Fig. 1c) has a length of about 1.5 km and a maximum width of 0.5 km. It drains a relatively small catchment of 67 km2. At present, both estuaries are separated from the open sea by ephemeral sand barriers (Schneider et al. 2010) and are mostly used for agriculture and golf courses. A small part of the Quarteira estuary includes a bird sanctuary. For more detailed descriptions, see Teixeira and Pinto (2002), Terrinha et al. (1999), Teixeira (2005) and Teixeira et al. (2005) and Carvalho et al. (2012).

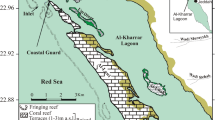
Location of study areas (a) and detailed maps of estuaries of Quarteira (b), Carcavai (c) and São Lourenço (d) with coring positions (black dots)
The third area of investigation is the estuary of the Ribeira de São Lourenço (Fig. 1d), which extends from the present coastline to the north about 5 km inland, with a width of up to 2.5 km. The Ribeira de São Lourenço flows approximately 6 km west of Faro into the Atlantic. The coastal area ahead of the mouth region can be assigned to the western area of the lagoons, dunes and marshes of the Ria Formosa (Moura et al. 2006). The estuary drains an area of Jurassic and Cretaceous limestone, marl, calcareous marl and dolomite. The coastal zone is dominated by non-consolidated sands of the Plio-Pleistocene (Boski et al. 2002). The floodplain area of the Ribeira de São Lourenço and the Ria Formosa includes upstream alluvium, beaches and dunes. Since the Neogene, the shelf area was affected by the development of strong terrestrial sedimentation, resulting in an accumulation of sand bodies several meters in thickness. The sands and gravels of this ‘Red Formation’ are currently exposed in the cliffs between Albufeira and Faro (Boski et al. 2002).
The climate of the Algarve coast is described by Teixeira and Pinto (2002) as typical Mediterranean with dry, hot summer months. The average annual rainfall is 500–600 mm, with 80 % of the rain falling from October to March. A moderate wind regime prevails mainly from a south-westerly direction with mean annual wind speeds of 6–9 km/h (Barnhardt et al. 2002). The mean annual wave height is ~1 m, while storm events the average wave height reaches ~3 m (Reyes et al. 1999). Wave heights of 6 m are expected on a 30- to 50-year return interval (Andrade et al. 2004). Semi-diurnal tides occur with a mean tidal range of 2 m and maximum spring tide of 3.5 m (Boski et al. 2002; Teixeira and Pinto 2002).
Materials and methods
Field work and sampling
In total, 53 sediment cores (Quarteira: 43, Carcavai: 6; São Lourenço: 4) were retrieved with a piston corer to reconstruct the development of the three estuaries. One core of each investigated area was used for detailed analyses including lithology, geochemistry, palynology, micropalaeontology and carbon dating.
The core locations (Table 1) were marked using a differential GPS (Magellan ProMark X) with reference to the Pinhal trigonometric point in Vilamoura (UTM 29, 4105685.129N, 579626.643E; m.s.l.: 60.25) as detailed in Schneider et al. (2010).
In the laboratory, core sections were split longitudinally, visually described and photographed. While one-half of the core was archived, the other half was subsampled at 4-cm intervals, with 1-cm-thick subsamples taken for various analyses.
Sedimentological and geochemical analyses
For ease of comparability between all study areas, the same analytical methods were used for all investigated cores. After pre-treatment, particle sizes and distributions were measured using a Beckmann Coulter Laser Diffraction Particle Size Analyser (LS 13320). The geochemical investigations of total carbon (TC), total organic carbon (TOC), total nitrogen (TN) and total sulphur (TS) were determined with an Elemental Analyser (Vario EL). Carbonate content was determined according to the wet sediment’s reaction to HCl (AG Boden 2005). Detailed descriptions can be found in Schneider et al. (2010) and Trog et al. (2013).
Micropalaeontological and statistical analyses
The micropalaeontological analysis follows the procedure described by Trog et al. (2013) and is based on 29 sediment samples from the Quarteira core and 34 from Carcavai. Only four samples of the São Lourenço core were available for analysis. A 2-g sample of sediment was washed with tap water through sieves of >63-, >200-, >630- and >2000-μm mesh size and dried in the different size fractions at 40 °C for 24 h. Foraminifers, ostracods, plant remains, bivalves and gastropod fragments were counted. Because of the very small sample size, all available tests and valves were picked and studied. Extraction and identification were performed under a standard binocular microscope (magnification 2.5 × 10) and using various identification guides, e.g. Sliter (1980), Galhano (1963), Murray (1979, 1991, 2006) and Loeblich and Tappan (1987) for foraminifers and, e.g. Athersuch (1976), Athersuch et al. (1989), Frenzel et al. (2010), Meisch (2000) for ostracods. [A complete taxonomic reference list is given in Appendices 1 and 2. Images of some species are shown in Figs. 8 and 9 (see below)]. Ecological classification also relies on those publications. For diversity calculations, the Shannon index (cf. Murray 2006) and the Fisher alpha index (Fisher et al. 1943) were used. In contrast to the Shannon index, the alpha diversity index has a high discriminant ability for species numbers. The Renkonen index was used to compare the microfaunas of succeeding samples in order to identify significant changes in species composition as a base for defining microfossil units (Müller 1991). A threshold of 60 % was applied. As in Trog et al. (2013), salinity classification followed the Venice system (Symposium on the Classification of Brackish Waters 1958).
The material is housed at the Senckenberg Research Institute and Natural History Museum Frankfurt am Main.
Radiocarbon dating
Dating procedures described by Schneider et al. (2010) were used to subsample datable material from the cores. These were dated at external laboratories (Beta Analytic, USA and Erlangen, Germany) and the resulting ages calibrated with OxCal v.3.10 (Ramsey 2001). To avoid hard water effects (Soares and Dias 2006, 2007; Soares and Martins 2009), terrestrial organic matter was used. The results of the AMS C14 dating are listed in Table 2.
Results
Sedimentological, geochemical and radiocarbon dating results
Sedimentary units
Based on sedimentological research in the Quarteira and Carcavai estuaries, Schneider et al. (2010), Dabrio et al. (2000), Andrade et al. (2004), Boski et al. (2008), Hilbich et al. (2008) and Vis et al. (2008) have classified sediment deposits along the Algarve coastline into six facies. Trog et al. (2013) extended the number of units based on new results from the estuaries of Alvor and Alcantarilha. A complete list of all six units, along with their grain-size distribution, colour, calcium carbonate content and macro remains of plants and microfossils is given in Table 3.
A full description of the core units from Quarteira (P01-5) and Carcavai (VdL PB2) estuary is given in Schneider et al. (2010), hence only a brief synopsis of results follows. The new São Lourenço core, see section “São Lourenço estuary (SAL 02-03/08)”, will be described in more detail below and in Fig. 2.

Sedimentological analyses of São Lourenço (SAL 02-03/08)
Quarteira estuary (P01-5)
Core P01-5 of the Quarteira estuary can be subdivided into five units, I, IIa, IIb, IIc and III (lithological profile in Figs. 3 and 4). The basal facies I (700–664 cm) contains yellowish brown to brown sands and gravel. Nitrogen and sulfur levels are low (close to detection limit). CaCO3 is lacking due to the absence of macrofossils. Unit IIa (664–389 cm) comprises brownish grey to brown, coarse to medium sands with scattered gravels. Organic carbon, sulfur and nitrogen occur at low levels, and inorganic carbon content increases significantly up to ~4 %. The carbonate content ranges between 7 and 10 %. Macro-remains of molluscs and foraminifera are common. Two samples of organic matter from the lower part of this unit yielded radiocarbon ages of 4763 ± 39 BP (5330–5590 cal BP) and 4619 ± 42 BP (5070–5470 cal BP). Between 389 and 350 cm depth, the deposits are assigned to unit IIb. They are characterised by upward fining brownish grey silty sands to sandy silts. Nitrogen, sulfur and organic and inorganic carbon values increase. Carbonate content ranges from 2 to 4 %. Mollusc and foraminiferal remains are frequent. Two radiocarbon dates place this unit between 3977 ± 38 BP (4290–4530 cal BP) and 2979 ± 42 BP (3006–3324 cal BP). Unit IIc (350–150 cm) comprises mottled grey and brown clayey silt with increasing calcium carbonate content upwards. Sulfur decreases to the detection limit at the surface. The unit is barren of macrofossils. The upper part is younger than ~950 cal BP. The sediment of unit III (150–0 cm) is composed of dull reddish brown clayey silts with small sand content. Nitrogen and organic content as well as inorganic carbon increase toward the top. Calcium carbonate ranges between 7 and 10 % and calcareous fragments of terrestrial snails are evident.
Carcavai estuary (VdL PB2)
The basal unit I of core VdL PB2 (832–796 cm) consists of greyish yellow medium to coarse sand, devoid of gravels, mollusc fragments and calcium carbonate (lithological profile in Figs. 5 and 6). Sulfur content remains barely above the detection limit. Carbonate content is not detectable due to the absence of mollusc and snails shell fragments. At a depth of 810 cm, plant remains yielded a radiocarbon age of 7300 ± 40 BP (8010–8180 cal BP). Unit IIa is absent from this core and is replaced with a tripartite unit IIb. The basal part between 796 and 620 cm consists of brownish black silty clays with a low sand and carbonate content. Carbonate, nitrogen and sulfur contents vary considerably. The middle section, between 620 and 450 cm, consists of grey sand with decreasing clay and silt content. Molluscs and microfossils increase in frequency upwards, as reflected in the increasing carbonate content. The upper part of this unit (450–336 cm) comprises grey silty sands. Values of sulfur, carbonate and nitrogen increase. Ten samples of this facies ware dated with an age range from 6210 ± 40 BP (7000–7250 cal BP) to 2351 ± 41 BP (2213–2679 cal BP). The deposits in unit IIc (336–189 cm) consist of brownish black and dark greyish yellow silts with thin layers of sand. Carbonate, nitrogen and sulfur content increase from the middle of this section. Due to the absence of datable material, a radiocarbon age of 1500 ± 40 BP (1410–1610 cal BP) was inferred from a peak in pollen (Schneider et al. 2010). The top layer sediment of unit III (189–0 cm) is made of brownish black to brown clayey silts with high values of sand and silt. While carbonate and sulfur remain at the same level, nitrogen increases significantly. Charcoal particles, as well as plant remains, are common. A distinct layer of medium to coarse sands with fragments of shells and snails is imbedded in this unit between 114 and 126 cm. The three radiocarbon datings for this unit yields ages of 990 ± 40 BP (800–960 cal BP), 160 ± 40 BP (0–290 cal BP) and 306 ± 59 BP (154–500 cal BP). Possible explanation for this inversion is the redeposition of the dated charcoal material by fluvial transport (cf. Schneider et al. 2010).
São Lourenço estuary (SAL 02-03/08)
Core SAL 02-03/08 is composed of five units following the definition of Schneider et al. (2010) and Trog et al. (2013) (see Fig. 6, below). Unit I is split into a lower and an upper part. The lower part, between 860 and 772 cm, is composed of yellow brownish, loamy sediment with a mean sand content of 63 %. The carbonate content of this section is 4-7 %. Mollusc fragments are lacking, whilst fossil plant remains are common. The upper part (772–696 cm) consists of brownish-grey, sandy deposits with scattered mollusc fragments. Sand content rises from 17 to 90 % and sulfur also increases.
Unit IIa (696–656 cm) is comprised of dark grey, primarily silty sediment. Sand content decreases abrupt from 94 to 26 %, sulfur and TOC content fluctuate between 0.3 to 2.6 % and 0.1 to 2.3 %, respectively. Calcium carbonate decreases from 2-4 % to 0.5-2 % (Fig. 2). There are many molluscs fragments, with whole clams and snails included.
Unit IIb is characterised by medium to dull grey slightly sandy silts. In the lower part between 656 and 508 cm silts and clays (66-97 %) are common. Sand content amounts to only 3-34 %. Sulfur concentration shows a declining trend from 2.0 to <0.1. The deposits have a carbonate content of 0.5-2 %, whilst coarser mollusc fragments are imbedded in the sediment. The upper part (508–166 cm) contains characteristic sand layers of a few centimetres thickness. Sulfur fluctuates markedly around 1.4 %, with decreases corresponding to the sandy layers. Carbonate content varies between 1 and 7 %. Fragments of molluscs, unbroken shells and snails are common in the whole of unit IIb. In addition to the grey colour of the sediment, black stiae are visible, resulting from indissoluble ferrous sulfide reduction under anaerobic conditions (Håkanson and Jansson 1983).
Two radiocarbon datings indicate ages of 3100 ± 40 cal BP at 431.5 cm and 1310 ± 40 cal BP at 265 cm. The grey, dull olive coloured sediments of unit IIc (166–93 cm) are mottled greyish blue and contain rust marks. The clay and silt fractions capture, on average, 78 % of the grain-size spectrum. Sand content decreases toward the top from 32 to 2 %. Sulfur concentration changes strongly from 2.4 to >0.1 %. Carbonate concentration ranges between 0.5 to 7 % related to the presence of mollusc fragments, and decreases upwards.
The upper 93 cm (unit III) of the core are coloured reddish-brown. The sediment consists of 54 to 82 % silt, while sand content is reduced to an average of 2 %. Sulfur content remains very low (<0.1 %). Increasing TOC indicates a rising deposition and preservation of organic matter. Carbonate concentration increases to 4-7 % and mollusc fragments were detected only sporadically.
Micropalaeontological results
Quarteira estuary (P01-5)
The distribution of ecological groups within foraminifer and ostracod associations paints the following picture of the changing environment:
The core section between 700 and 500 cm was not analysed. The lowermost section analysed for microfossils is unit A (500–410 cm). The largest abundances of marginal marine and marine-brackish foraminifer taxa are found within this unit (Fig. 3). Marginal marine foraminifera (Elphidium advenum, E. margaritaceum) occur but marine-brackish taxa (Ammonia spp., Haynesina germanica, Cribroelphidium excavatum, C. oceanensis) with high salinity tolerances are dominant. The brackish phytal species Cribroelphidium williamsoni are less common. In terms of ostracods (Fig. 4), the section is dominated by brackish water taxa (Cyprideis torosa, Cytherois sp., Leptocythere sp., Loxoconcha elliptica and Callistocythere murrayi). The taxa indicative of other habitats, such as freshwater and phytal environments, are less common, probably transported (Fig. 4).

Abundances of foraminifers taxa in the Quarteira estuary (P01-5) with microfossil units, microfossil hiatus (H1–H3), diversity indices and ecological groups

Abundances of ostracods taxa in the Quarteira estuary (P01-5) with microfossil units, microfossil hiatus (H1–H3), diversity indices and ecological groups
The next unit containing microfossils is B (380–290 cm) and comprises nearly exclusively marine-brackish foraminifera taxa (e.g. Ammonia spp., Haynesina germanica). Marginal marine taxa as well as phytal taxa are absent. Cyprideis torosa and Loxoconcha elliptica, both brackish water species, are the only representatives of ostracods in this unit but rare. Both are rare and the valves reworked, probably due to a transport to this location.
The next microfossil-bearing section after the second hiatus is unit C (240–210 cm) which includes mainly marine-brackish taxa (Ammonia spp., Haynesina germanica, Cribroelphidium spp., C. poeyanum, Discorynopsis aguayoi) and one instance of marginal marine foraminifera (Elphidium advenum). Present ostracods of this habitat are the brackish water forms Leptocythere sp. and Loxoconcha elliptica.
The near surface core section between 150 cm and the top was not subjected to detailed analyses because the sediments clearly appeared to be overbank deposits, probably redeposited material with poor variations.
Carcavai estuary (VdL PB2)
Unit A1 (790–710 cm) in the Carcavai core is the first microfossil zone with preservation of foraminifers and ostracods. The dominant foraminifers are marginal marine (Elphidium margaritaceum) and marine-brackish taxa (e.g. Ammonia spp., Haynesina germanica, Cribroelphidium excavatum, C. oceanensis, C. poeyanum) (Fig. 5). The only identified typical brackish-water taxon is Cribroelphidium williamsoni. The most common ostracods (Cyprideis torosa, Loxoconcha elliptica, Leptocythere spp., Cytherois fischeri, Callistocythere cf. murrayi and Loxoconcha malcomsoni) are characteristic of brackish habitats (Fig. 6). Marginal marine species (e.g. Urocythereis britannica) are rare.

Abundances of foraminifers taxa in the Carcavai estuary (VdL PB2) with microfossil units, microfossil hiatus (H1–H3), diversity indices and ecological groups

Abundances of ostracods taxa in the Carcavai estuary (VdL PB2) with microfossil units, microfossil hiatus (H1–H3), diversity indices and ecological groups
The following zone A2 (710–460 cm) shows only two foraminiferal taxa of marine-brackish habitats (Ammonia, Haynesina germanica) and shore line inhabiting trochamminid foraminifers. The ostracod assemblage is also characterised by brackish ostracods (Cyprideis torosa (noded and unnoded forms), Cytherois fischeri, Loxoconcha elliptica). Phytal ostracods are represented by rare Xestoleberis communis.
A thick sand layer accompanying unit A3 (460–350 cm) yields a similar ostracod inventory as unit A1, but with a higher proportion of brackish-water species (Cyprideis torosa [noded and unnoded forms], Cytherois fischeri, Leptocythere spp., Loxoconcha elliptica) and some phytal taxa (Xestoleberis aurantia, Xestoleberis communis). Marine-brackish species decline in this unit whereas rare marginal-marine taxa (Thaerocythere hoptonensis, Urocythereis britannica) appear to be brought in by storm surges. The most common foraminiferal taxa indicate marginal marine (e.g. Elphidium margaritaceum, E. advenum, miliolids) and marine-brackish (e.g. Ammonia spp., Haynesina germanica, Cribroelphidium oceanensis, C. excavatum, C. poeyanum) habitats. In addition, brackish-water and a phytal species are documented.
Microfossil unit B (260–130 cm) is almost barren of foraminiferal tests. The few individuals of Ammonia and Haynesina germanica are representative of a marine-brackish milieu. The ostracod assemblage is dominated by freshwater taxa (Candona juv., Darwinula stevensoni, Herpetocypris sp., Heterocypris salina, Ilyocypris bradyi). Marine species are absent.
São Lourenço estuary (SAL 02-03/08)
Only four sediment samples have been investigated for microfauna and two were barren. Altogether 11 taxa of ostracods and 9 foraminiferal taxa are present in the São Lourenço core (Fig. 7). Samples from 824 and 744 cm are barren of ostracod valves and foraminifer tests probably due to the sandy, well-sorted sediment. At a depth of 680 cm, marginal-marine foraminifera (Quinqueloculina sp.) dominate the sample whereas marine-brackish taxa (Haynesina germanica, Ammonia spp.) occur in small numbers (Figs. 7, 8, 9). Marine-brackish forms (Xestoleberis spp., Pontocypris sp., Loxoconcha rhomboidea and Leptocythere macallana) and brackish taxa (Cyprideis torosa, Cytherois spp. and Loxoconcha elliptica) dominate the ostracod assemblage. Marine species, such as Aurila arborescens and Leptocythere pellucida, are rare.

Abundances of foraminifers and ostracods taxa in the São Lourenço estuary (SAL 02-03/08) classified into ecological groups

Foraminifera. a, b Ammonia tepida (Cushman, 1926) P01-5–31: a1 spiral view; a2 lateral view; b umbilical view (SMF XXVII 14414); c Haynesina germanica (Ehrenberg, 1840) VdL PB2–524: spiral view (SMF XXVII 14415); d Cribroelphidium oceanensis (d’Orbigny, 1826) P01-5–37: d1 spiral view; d2 lateral view (SMF XXVII 14416); e Cribroelphidium poeyanum P01-5–37: e1 spiral view; e2 lateral view (d’Orbigny, 1839) (SMF XXVII 14417); f Cribroelphidium excavatum (Terquem, 1875) P01-5–33: spiral view (SMF XXVII 14418); g Discorinopsis aguayoi (Bermúdez, 1935) P01-5–9: spiral view (SMF XXVII 14419); h Helenina anderseni (Warren, 1957) P01-5–38: oblique view (SMF XXVII 14420); i Cribroelphidium williamsoni (Haynes, 1973) VdL PB2–570: spiral view (SMF XXVII 14421); j Elphidium margaritaceum Cushman, 1930 VdL PB2–570: spiral view (SMF XXVII 14422); k Cibicides sp. VdL PB2–524: spiral view (SMF XXVII 14423); l Quinqueloculina sp. VdL PB2–586: lateral view (SMF XXVII 14424)

Ostracoda. a Thaerocythere hoptonensis (Brady, Crosskey and Robertson, 1874) VdL PB2–524: juvenile left valve (SMF Xe 23117); b Xestoleberis aurantia (Baird, 1838) VdL PB2–524: female left valve (SMF Xe 23118); c Callistocythere murrayi Whittaker, 1978 VdL PB2–570: juvenile right valve (SMF Xe 23119); d Darwinula stevensoni (Brady and Robertson, 1870) VdL PB2–532: female left valve (SMF Xe 23120); e Cytherois fischeri (Sars, 1866) VdL PB2–524: adult right valve (SMF Xe 23121); f Leptocythere sp. VdL PB2–539: juvenile left valve (SMF Xe 23122); g Loxoconcha elliptica Brady, 1868 P01-5–38: female left valve (SMF Xe 23123); h Urocythereis britannica Athersuch, 1977 VdL PB2–539: juvenile right valve (SMF Xe 23124); i Cyprideis torosa (Jones, 1850) VdL PB 2–539: female right valve (SMF Xe 23125); j Heterocypris salina (Brady, 1868) VdL PB 2–515: adult left valve (SMF Xe 23126); k Cyprideis torosa (Jones, 1850) VdL PB 2–540: male left valve (SMF Xe 23127)
The sample from 580 cm depth is richer in foraminifer taxa than the previous one. The balance of marginal-marine (Bolivina sp., Elphidium crispum, Nonionella sp., Quinqueloculina sp.) and marine-brackish foraminifera (Ammonia spp., Cribroelphidium excavatum, Haynesina germanica) is almost equal. The ostracod association is dominated by Loxoconcha rhomboidea, Cytherois spp., and Leptocythere pellucida. The former taxon is a typical marine-brackish species and the latter is a marginal-marine species like the less common Aurila arborenscens. Many Cytherois species are typical brackish taxa. Some other marine-brackish species were also found, e.g. Leptocythere macallana, Palmoconcha laevata, Pontocypris sp. and Xestoleberis spp.
Discussion
On the basis of micropalaeontological, sedimentological, geochemical and geochronological results a reconstruction of the genesis of the three estuaries can be produced (Fig. 10). Following the models of Schneider et al. (2010) and of Trog et al. (2013), three stages of development are discussed: (1) the pre-transgressional to transgressional evolution, (2) the end of relative sea-level rise as a standstill for intensive marine influence, and (3) decreasing tidal influence including post-estuarine development.

Synopsis of multi-proxy analyses for all three cores studied
Pre-transgressional to transgressional evolution (before ~6500 cal BP)
The basal deposits (unit I) of the estuaries are fluvially transported fine-grained sediments characterising the palaeovalley of the rivers, as seen in previous studies by Hilbich et al. (2008), Schneider et al. (2010) and Trog et al. (2013). Based on the sea-level curve of Vis et al. (2008), the top of the basal fluvial sediments in the Quarteira estuary date to ~7500-8000 cal BP suggesting the start of marine transgression into the Quarteira palaeovalley at this time (Schneider et al. 2010), whilst a radiocarbon date from the top of the lowermost core section of Carcavai provides an age of 7300 ± 40 BP (8010–8180 cal BP). The flooding of the Carcavai palaeovalley by brackish water started at approximately 8000 cal BP, as indicated by the lack of microfossils in unit I and the subsequent change to marine-brackish to brackish conditions in unit A1. Such conditions are indicated by the foraminiferal species Cribroelphidium excavatum, occurring in inter- to subtidal habitats with changing salinities and temperatures (Murray 2006) and Quinqueloculina seminulum associated with Haynesina germanica “suggesting a restricted and brackish environment, hardly reached by marine water from the open sea” (Alday et al. 2013, p. 80) or “marine areas close to the river mouth” (Font et al. 2013, p. 7). The representative ostracod taxa Cyprideis torosa and Loxoconcha elliptica are both index forms for brackish water (Athersuch et al. 1989).
In the subsequent coarse-grained sediments of unit IIa in Quarteira with its sands and gravels, and the fine sediments of unit IIb in Carcavai, textural characteristics (changing from coarse sand to silty clays with some very prominent sand layers) and geochemical properties (strongly fluctuating calcium carbonate content) indicate tidal channel deposits along a braided river system comparable to the Lower Tagus valley (Vis et al. 2008; Vis and Kasse 2009). The ostracod association of Carcavai (unit A2) consists of mainly brackish taxa that are adaptable to a wide range of salinities and temperatures. Ruiz et al. (2000, p. 345) assume that Loxoconcha rhomboidea is “mainly developed in the quiet tidal channels connected with the main estuarine channels in the marine domains” and “Loxoconcha elliptica is dominant in channels and channel margins with fine sediments, high salinity variations and low to moderate hydrodynamic levels”. After Athersuch et al. (1989), the species Cyprideis torosa is cosmopolitan e.g. in estuaries and lagoons and can be “found in a wide range of salinities”. The dominating marine-brackish foraminifers, e.g. Ammonia spp. and Haynesina germanica, prefer loamy or sandy habitats with varying organic carbonate content (Murray 2006) and tolerate salinities from 15 to 31. The less frequent occurrence of shore line and marginal-marine foraminifers suggests tidal transport within the estuary. In summary, the microfaunal remains also point to a marine ingression into the river valley similar to that reconstructed for the nearby Santo André Lagoon (Freitas et al. 2002b, 2003; Cearreta et al. 2003), the Mira estuary (Alday et al. 2006) and the Lower Tagus valley (Vis et al. 2008; Vis and Kasse 2009).
The grain-size spectrum of the stratigraphic unit I of São Lourenço also indicates the sediment to be transported by a stream (e.g. Boggs 1987). The absence of mollusc fragments and the very low sulfur content point to a fluvial depositional environment. Plant residues deposited in increasing amounts in the sediment could be derived from floodplain vegetation or have been transported by the river from the catchment. It seems very likely that this section represents a fluvial–terrestrial sediment complex and thus the former river valley before postglacial transgression. At the time of sea-level low stand, the river valley was therefore formed by the interaction of processes of fluvial erosion, accumulation of alluvial fans and soil formation. From about 736 cm total core depth, an increasing marine influence on the water body is evident from a slight increase in sulfur concentration (Schlesinger 1991), as well as the first occurrence of marine mollusc fragments. This could be interpreted as the initiation of postglacial transgression processes and the incipient marine flooding of the river valley. In consideration of the altitude (0.48 m above m.s.l.) and the graph of relative sea-level rise of Vis et al. (2008), the period of deposition in the palaeo-valley of the Ribeira de São Lourenço can be estimated. The fluvial sediments accordingly represent a period of ~8000–7500 cal BP. Schneider et al. (2010) set the age of the Quarteira estuary palaeovalley at ~7500 cal BP using the same approach.
The sharp decline in sand content in unit IIa of São Lourenço points to a landward retreat of the beach front and increasing water depth in the estuary. Under these conditions, the deposition energy and sedimentation of fine particles and organic material (TOC) decrease. Mollusc fragments, including whole clams and snails, are present in increasing numbers, indicative of a marginal marine depositional environment. Moreover, ostracods and foraminifera associated with brackish to marginal marine environments occur for the first time. The ostracod genus Xestoleberis is characteristic for shallow water areas (e.g. Cabral and Loureiro 2013). The foraminifer genus Quinqueloculina occurs at salinities from 32 to 65 in marginal marine environments (e.g. Murray 2006). Therefore, we suggest that an influx of sea water induced the development of a small, shallow estuary with strong water exchange with the open sea. Using the environmental tolerances of the identified microfauna, a reconstruction of the transition from fluvial to marine environment is possible. Unit IIa represents sedimentation during sea-level rise as part of the post-glacial transgression leading to inflow of sea water and increasing water depths in the estuary. River valleys are usually the starting point for the development of estuaries during the Pleistocene sea-level low stand (e.g. Nichols and Biggs 1985). The first evidences of the marine transgression in Gilão-Almargem estuary, which is located in the east of São Lourenço estuary and the Ria Formosa, were dated at ~8000 cal BP (Boski et al. 2008).
End of relative sea-level rise—a phase of intensive marine influence (between ~6500 and ~1000 cal BP)
A transition from open-marine to marginal marine and marine-brackish environments occurred after the sea level reached its Holocene high stand. In the Quarteira estuary, the lithological units IIa, IIb and parts of IIc and microfossil units A and B are included in this transition. The grain-size distribution shifts to fine-grained deposits with an increase of nitrogen. Typical taxa for this confined marine-brackish habitat are Ammonia beccarii, Haynesina germanica and Discorynopsis aguayoi (Murray 2006). The ostracod species Cyprideis torosa and Loxoconcha elliptica are indicative for estuaries with oligo- to polyhaline environments (Frenzel et al. 2010) reflecting brackish water influence.
In the Carcavai estuary, this phase is mirrored in lithological units IIb and IIc and in the microfaunal units A2, A3 and part of B. Between the middle and the top part of unit IIb, an increase in sand fraction is visible. This, and the frequency of marine mollusc fragments as well as the significant changes in the sulfur, TOC and carbonate content, suggest a change in the depositional environment. A distinctive layer of coarse sand with broken shells in the middle part of the unit is detectable in two parallel cores. The absence of these sands in core VdL PB2 may be due to small-scale geomorphological differences in the estuarine system. Dates between 462 and 433 cm yielded various ages between 5486 ± 57 BP (6296–6403 cal BP) and 3070 ± 40 BP (3290–3370 cal BP), which would correspond to a sedimentation rate of only 29 cm in 3000 years (see Schneider et al. 2010). It is more likely that there was an interruption in deposition, maybe caused by a regional climate change around 4000–3000 BP (cf. Queiroz and Mateus 2004). As seen in Alvor, the hypothesis of a storm event or tsunami “is supported by low average sedimentation rates in this period, reflecting a loss of surface sediments after backwash events” (Trog et al. 2013, p. 153). This coincides with the occurrence of marginal marine foraminifers, e.g. Elphidium margaritaceum, and marine-brackish taxa (Cribroelphidium oceanensis, Cribroelphidium excavatum, C. poeyanum; Murray 1979) as well as the brackish water indicator Cyprideis torosa (e.g. Pint et al. 2012) and the marine ostracod Thaerocythere hoptonensis (see Cabral and Loureiro 2013).
After that disturbance, regular silting up restarts at ~3000–2300 cal BP reflected by fine sediments. The formation of a sandy barrier in front of the shallow estuary is probable, comparable to the Santo André Lagoon (Freitas et al. 2002a). Schneider et al. (2010) interpreted the small sandy layers embedded in clayey silts in the upper part of unit IIb as marine deposits resulting from instability in the inferred barrier system. Unit IIc (~2300 and ~1000 cal BP) indicates “the accumulation of mostly fluvially derived material from the catchment and possibly temporary disturbances to the barrier system recorded in several small sandy layers of marine origin” (Schneider et al. 2010, p. 16). The microfossil assemblage is dominated by brackish species but phytal and freshwater species as Ilyocypris bradyi (e.g. Carbonel and Jouanneau 1982; Ruiz et al. 2013) are also detectable in the top of this phase.
Due to the consistently low sand content in the lower part of unit IIb (508–656 cm) in the São Lourenço core, low-energy depositional conditions in the estuary can be deduced (e.g. Boggs 1987). It is likely that the estuary was isolated from marine influence after the beginning of the Flandrian transgression. It is also possible that a spit formed at the mouth of the estuary as a result of attenuation of post-glacial sea-level rise (Freitas et al. 2002a). The microfauna of the analysed sediment sample from 580 cm total core depth also demonstrate a brackish-marine depositional environment in the estuary.
The sand layers in the upper segment (166–508 cm) indicate phases of higher-energy environmental conditions. The entire core section is characterised by fragments of molluscs and carbonate content between 2 and 25 %. Sand layers may represent smaller storm events, leading to input of sediment eroded from the sand barriers. Two charcoal samples from sand layers yielded ages of 3305 and 1235 cal BP (431–432 and 265 cm total core depth). Similar records of temporary barrier opening were established, among others, in the lagoon of Melides and in the Carcavai estuary. With increased permeability of the barrier, marine input is characterised by the deposition of coarse-grained sediments. Episodes with a lower barrier permeability favour low-energy deposition of predominantly terrestrial material (Freitas et al. 2002a; Schneider et al. 2010).
The sediment section between ~200 and 250 cm total core depth is characterised by a continuously high sand content. Coarse sand in the entire core occurs almost exclusively in the sand layers, where it constitutes only 1-4 %. In this section (200–250 cm), however, coarse sand comprises 4-24 %. Complete oyster and marine snails were found in this section, suggesting the deposition of this sediment could have been caused by a tsunami or major storm surge.
The dating of charcoal of unit IIb yields ages of ~3100 and ~1310 cal BP, respectively. Assuming that the charcoal originated from older branches or trunks and, taking into account possible reworking of the dated material, our data point to the tsunami triggered in 1755 by the Lisbon earthquake (Baptista et al. 1998; Hindson et al. 1996; Ruiz et al. 2005). According to the age-depth model for the estuary of the Ribeira de São Lourenço, an age of ~1000 cal BP would be obtained for the sand peak at 225 cm core depth. Because tsunami events unleash a strong erosive force (e.g. Kortekaas and Dawson 2007; Morton et al. 2007), a temporal hiatus is probable. The deposition of this massive sand layer by the 1755 tsunami appears to be most likely. However, it cannot be excluded that these sand deposits represent one or more major storm events. The sediment areas outside the sand layers in the upper part of unit IIb are an indication that phases of separation from the open-marine influence have taken place, allowing terrestrial biomass to be incorporated. Thus, sedimentation in the estuary is registered in this core section as a fluctuating imbalance of marine and terrestrial pulses. Because mollusc fragments are continuously present throughout the core segment, and whole snails and clams were found, a predominantly brackish environment is most probable.
The sedimentary unit IIb represents an estuarine system with a fluctuating terrestrial and marine input. The existence of a naturally formed barrier of varying strength and permeability is suggested. Estuarine systems are generally under both marine and terrestrial influence and are characterised by varying tidal and fluvial processes (e.g. Schlesinger 1991; Nichols and Biggs 1985). According to Nichols and Biggs (1985), the development and life span of an estuary depends on the ratio of sea-level rise to sediment delivery from the terrestrial catchment area. After the formation of an estuary, it gradually develops a balance between sea-level rise and fluvial sediment input. If the rate of sea-level rise exceeds the fluvial sediment input, the estuary retains its original size (e.g. Nichols and Biggs 1985).
The formation of natural barriers in the form of spits takes place with stabilisation and weakening of the sea-level rise under some circumstances. Therefore, the material supplied from the sandy cliffs of the western Algarve provides the eastwards longshore current with clastic material (littoral drift; see Boski et al. 2008). The barriers lead to closure and reorganisation of estuaries which will progressively become brackish and lagoonal conditions prevail. The maximum extent of the estuary of the Ribeira de São Lourenço was reached in unit IIb, most likely in the lower low-energy part.
According to Freitas et al. (2002a), the Portuguese coastal area has been impacted by very high rates of sea-level rise since the Lateglacial to about 7000–6000 BP. Boski et al. (2002) estimate the first phase of the Holocene sea-level rise at a rate of 0.85 m/century until ~6500 BP. A second phase with lower rates of 0.3 m/century continued until ~5000 BP when the recent level was reached (Boski et al. 2002). Freitas et al. (2002a) assign the beginning of sea-level stabilisation and the formation of sandy barriers to the same period. The development of barriers in the lagoon of Santo André took place at about 5380 ± 50 BP and in Melides no later than 4500 BP (Freitas et al. 2002a).
Decreasing tidal influence and post-estuarine development (between ~1000 cal BP and the present)
With increasing siltation, the marine influence disappears and brackish to freshwater conditions dominate in the Quarteira estuary in recent times [lithological units IIc (only part of it) and III, microfossil unit C]. The foraminifer Discorynopsis aguayoi typical for confined coastal water bodies (Murray 2006) confirms this assumption. Moreover, the ostracod taxa Loxoconcha elliptica and Leptocythere sp. reflect brackish conditions. According to Schneider et al. (2010, p. 156), “the post-estuarine development is characterised by overbank deposits that accumulated in a floodplain-like area”. Only some small sandy layers provide evidence of interruptions of marine origin.
The deposition of overbank sediments in the Carcavai core [lithological units IIc (part of it) and III, part of microfossil unit B] is characterised by Darwinula stevensoni and Heterocypris salina representing a freshwater to slightly brackish milieu (Meisch 2000) comparable with the uppermost deposits in the Santo André coastal area (Cearreta et al. 2003). As described by Ruiz et al. (2013), D. stevensoni tolerates salinities up to 15 ‰ and indicates freshwater conditions of shallow muddy ponds. Ilyocypris bradyi, also present in this unit, exclusively inhabits environments with salinities lower than 6 (Ruiz et al. 2013).
Deposition is interrupted by a prominent sand layer in unit III, which is most probably associated with the 1755 Lisbon tsunami (Schneider et al. 2010). Studies along the Algarve coast, e.g. Hindson and Andrade (1999), Hindson et al. (1996, 1999), and Allen (2003), show comparable results.
The large proportion of fine material and the decreasing trends among all grain-size parameters in sections IIc (part of it) and III of the São Lourenço core reflect increasingly low-energy depositional conditions. Under these circumstances, increased amounts of organic matter were deposited. The sediment of section IIc shows distinctive rust marks and mottling by iron and manganese oxides produced by a change in redox conditions resulting from groundwater fluctuations (e.g. Kuntze et al. 1994). Judging from the absence of mottling in section III, a groundwater influence can be excluded or it may be assumed that sedimentation occurred above the tidal water-table. A marine influence on sedimentation can also be excluded, since the sulfur concentration decreases below 0.1 % or is undetectable. The sporadic mollusc fragments may have been reworked from marine sediments or come from terrestrial snails.
The sedimentary sections IIc and III include the silting up of the estuary under fluvial, low-energy sedimentation. Alluvial sediments were created, comparable with the material of the lowermost section I. In particular, the TOC content in section III indicates that the newly formed floodplains were colonised by long-term vegetation.
A full closure of the estuary of the Ribeira de São Lourenço from open-marine influence is clearly indicated by the significant decrease in the concentration of sulfur in section IIc. From the subsequently increasing content of organic matter (TOC increase), it is assumed that dense vegetation colonised areas of the estuary under stable conditions.
The silting-up process begins in the area of Quarteira and Carcavai estuaries at ~1000 BP with the accumulation of alluvial fans and interrupted by brief marine pulses, as deduced from palynological and sedimentological results (Schneider et al. 2010). In the estuary of the Ribeira de São Lourenço, similar events cannot be found on the basis of the sedimentological results.
Conclusions
This paper reconstructs the landscape evolution of the estuaries of the Ribeira de Quarteira, Ribeira de Carcavai and Ribeira de São Lourenço for the last ~8000 years. The depositional conditions of the estuarine sediments are characterised using sedimentological, geochemical and microfaunal analyses of three sediment cores. This multidisciplinary study allows the correlation of sedimentological proxies with microfossil zones and three main palaeoenvironmental stages could be defined. These stages supply information about former sea-level, tidal regimes, and sediment reworking as well as transport processes. Furthermore, data on Holocene coastal biodiversity and palaeoecology are provided by documenting the distribution of marginal marine and brackish water foraminifera and ostracods of the Algarve coast for the three investigated estuaries.
The genesis of the three areas shows distinct similarities. The delayed onset of the transgression shifting from ~8000 to ~6500 cal BP in the three estuaries can be explained by the differing size of the rivers and their catchment as well as the morphology of the palaeo-valley.
References
Alday, M., Cearreta, A., Cachão, M., da Freitas, M. C., Andrade, C., & Gama, C. (2006). Micropalaeontological record of Holocene estuarine and marine stages in the Corgo do Porto rivulet (Mira River, SW Portugal). Estuarine, Coastal and Shelf Science, 66, 532–543.
Alday, M., Cearreta, A., Freitas, M. C., & Andrade, C. (2013). Modern and late Holocene foraminiferal record of restricted environmental conditions in the Albufeira Lagoon, SW Portugal. Geologica Acta, 11, 75–84. doi:10.1344/105.000001754.
Allen, H. D. (2003). A transient coastal wetland: from estuarine to supratidal conditions in the less than 200 years - Boca do Rio, Algarve, Portugal. Land Degradation & Development, 14, 265–283.
Andrade, C., Freitas, M. C., Moreno, J., & Craveiro, S. C. (2004). Stratigraphical evidence of Late Holocene barrier breaching and extreme storms in lagoonal sediments of Ria Formosa, Algarve, Portugal. Marine Geology, 210, 339.362. doi:10.1016/j.margeo.2004.05.016.
Athersuch, J. (1976). The genus Xestoleberis (Crustacea: Ostracoda) with particular reference to Recent Mediterranean species. Pubblicazioni della Stazione Zoologica di Napoli, 40, 282–343.
Athersuch, J., Horne, D. J., & Whittaker, J. E. (1989). Marine and brackish water ostracods (superfamilies Cypridacea and Cytheracea): keys and notes for the identification of the species. Leiden: E.J. Brill.
Baptista, M. A., Heitor, S., Miranda, J. M., Miranda, P., & Victor, L. M. (1998). The 1755 Lisbon tsunami; evaluation of the tsunami parameters. Journal of Geodynamics, 25, 143–157. doi:10.1016/S0264-3707(97)00019-7.
Barnhardt, W. A., Gonzalez, R., Kelley, J. T., Neal, W. J., Pilkey, O. H., Jr., & Dias, J. M. A. (2002). Geologic evidence for the incorporation of flood tidal deltas at Tavira Island, southern Portugal. Journal of Coastal Research, SI 36, 28–36.
Blázquez, A. M., & Usera, J. (2010). Palaeoenvironments and Quaternary foraminifera in the Elx costal lagoon /Alicante, Spain). Quaternary International, 221, 68–90.
Boden, A. G. (2005). Bodenkundliche Kartieranleitung (5th ed.). Stuttgart: Schweitzerbart.
Boggs, S. (1987). Principles of sedimentology and stratigraphy. New York: Merrill
Boski, T., Moura, D., Veiga-Pires, C., Camacho, S., Duarte, D., Scott, D. B., & Fernandes, S. G. (2002). Postglacial sea-level rise and sedimentary response in the Guadiana Estuary, Portugal/Spain border. Sedimentary Geology, 150, 103–122. doi:10.1016/S0037-0738(01)00270-6.
Boski, T., Camacho, S., Moura, D., Fletcher, W., Wilamowski, A., Veiga-Pires, C., Correia, V., Loureiro, C., & Santana, P. (2008). Chronology of the sedimentary processes during the postglacial sea level rise in two estuaries of the Algarve coast, Southern Portugal, Estuarine. Coastal and Shelf Science, 77, 230–244. doi:10.1016/j.ecss.2007.09.012.
Brandão, S. (2013). Podocopa. In: Brandão, SN, Angel, MV, Karanovic, I (eds), World Ostracoda Database., Accessed through:World Register of Marine Species at http://www.marinespecies.org/aphia.php?p=taxdetails&id=155944 on 2014-06-12.
Cabral, M. C., & Loureiro, I. M. (2013). Overview of Recent and Holocene ostracods (Crustacea) from brackish and marine environments of Portugal. Journal of Micropalaeontology, 32, 135–159. doi:10.1144/jmpaleo2012-019.
Cabral, M. C., da Freitas, M. C., Andrade, C., & Cruces, A. (2006). Coastal evolution and Holocene ostracods in Melides lagoon (SW Portugal). Marine Micropaleontology, 60, 181–204. doi:10.1016/j.marmicro.2006.04.003.
Carbonel, P., & Jouanneau, J. M. (1982). The evolution of a coastal lagoon system: hydrodynamics determined by ostracofauna and sediments - The Bonne-Anse Bay (Pointe de la Coubre, France). Geo-Marine Letters, 2, 65–70. doi:10.1007/BF02462802.
Carvalho, J., Matias, H., Rabeh, T., Menezes, P. T. L., Barbosa, V. C. F., Dias, R., & Carrilho, F. (2012). Connecting onshore structures in the Algarve with the southern Portuguese continental margin: The Carcavai fault zone. Tectonophysics, 570–571, 151–162.
Cearreta, A., Cachão, M., Cabral, M. C., Bao, R., & de Ramalho, M. J. (2003). Lateglacial and Holocene environmental changes in Portuguese coastal lagoons 2: microfossil multiproxy reconstruction of the Santo André coastal area. The Holocene, 13, 447–458.
Cearreta, A., Alday, M., Freitas, M. C., & Andrade, C. (2007). Postglacial foraminifera and paleoenvironments of the Melides lagoon (SW Portugal): towards a regional model of coastal evolution. Journal of Foraminiferal Research, 37, 125–135.
da Freitas, M. C., Andrade, C., Rocha, F., Tassinari, C., Munhá, J. M., Cruces, A., Vidinha, J., & Marques da Silva, C. (2003). Lateglacial and Holocene environmental changes in Portuguese coastal lagoons 1: the sedimentological and geochemical records of the Santo André coastal area. The Holocene, 13, 433–446.
Dabrio, C. J., Zazo, C., Goy, J. L., Sierro, F. J., Borja, F., Lario, J., González, & Flores, J. A. (2000). Depositional history of estuarine infill during the last postglacial transgression (Gulf of Cadiz, Southern Spain). Marine Geology, 162, 381–404. doi:10.1016/S0025-3227(99)00069-9.
Fisher, R. A., Corbet, A. S., & Williams, C. B. (1943). The relation between the number of species and the number of individuals in a random sample of an animal population. Journal of Animal Ecology, 12, 42–58.
Font, E., Veiga-Pires, C., Pozo, M., Nave, S., Costas, S., Muñoz, F. R., Abad, M., Simões, N., Duarte, S., & Rogríguez-Vidal, J. (2013). Benchmarks and sediment source(s) of the 1755 Lisbon tsunami deposit at Boca do Rio Estuary. Marine Geology, 343, 1–14. doi:10.1016/j.margeo.2013.06.008.
Freitas, da M. C., Andrade, C., Cruces, A., Amorim, A., Cearreta, A., Ramalho, M. J. (2002). Coastal environmental changes at different time-scales: the case of the Melides Barrier-Lagoon System (SW Portugal). Littoral 2002, The Changing Coast (pp. 397–402).
Freitas, M. C., Andrade, C., & Cruces, A. (2002). The geological record of environmental changes in southwestern Portuguese coastal lagoons since the Lateglacial. Quaternary International, 93–94, 161–170. doi:10.1016/S1040-6182(02)00014-9.
Frenzel, P., Keyser, D., & Viehberg, F. A. (2010). An illustrated key and (palaeo)ecological primer for Postglacial to Recent Ostracoda (Crustacea) of the Baltic Sea. Boreas, 39, 567–575. doi:10.1111/j.1502-3885.2009.00135.x.
Galhano, M. H. (1963). Foraminíferos da costa de Portugal (Algarve). Porto: Imprensa Portuguesa.
Håkanson L, Jansson M (1983) Principles of lake sedimentology. New York: Springer.
Hayward, B. W., Cedhagen, T., Kaminski, M., Gross, O. (2014). World Foraminifera Database. Accessed through: World Register of Marine Species at http://www.marinespecies.org/aphia.php?p=taxdetails&id=414702 on 2014-06-12.
Hilbich, C., Mügler, I., Daut, G., Frenzel, P., van der Borg, K., & Mäusbacher, R. (2008). Reconstruction of the depositional history of the former coastal lagoon of Vilamoura (Algarve, Portugal): a sedimentological, microfaunal and geophysical approach. Journal of Coastal Research, 24, 83–91. doi:10.2112/04-0411.1.
Hindson, R. A., & Andrade, C. (1999). Sedimentation and hydrodynamic processes associated with the tsunami generated by the 1755 Lisbon earthquake. Quaternary International, 56, 27–38.
Hindson, R. A., Andrade, C., & Dawson, A. G. (1996). Sedimentary processes associated with the tsunami generated by the 1755 Lisbon earthquake on the Algarve coast, Portugal. Physics and Chemistry of the Earth, 21, 57–63. doi:10.1016/S0079-1946(97)00010-4.
Hindson, R. A., Andrade, C., & Parish, R. (1999). A microfaunal and sedimentary record of environmentsl change within the late Holocene sediments of Boca do Rio (Algarve, Portugal). Geologie en Mijnbouw, 77, 311–321.
Kortekaas, S., & Dawson, A. G. (2007). Distinguishing tsunami and storm deposits: An example from Martinhal, SW Portugal. Sedimentary Geology, 200, 208–221. doi:10.1016/j.sedgeo.2007.01. 004.
Kuntze, H., Roeschmann, G., & Schwerdtfeger, G. (1994). Bodenkunde. Stuttgart: UTB.
Loeblich, A. R., & Tappan, H. (1987). Foraminiferal Genera and their Classification. New York: Van Nostrand Reinhold.
Loureiro, I. M., Cabral, M. C., & Fatela, F. (2009). Marine influence in ostracod assemblages of the Mira River estuary: Comparison between lower and mid estuary tidal marsh transects. Journal of Coastal Research, SI 56, 1365–1369.
Meisch, C. (2000). Crustacea: Ostracoda. Heidelberg: Spektrum.
Mendes, I., Rosa, F., Dias, J. A., Schönfeld, J., Ferreira, Ó., & Pinheiro, J. (2010). Inner shelf paleoenvironmental evolution as a function of land-ocean interactions in the vicinity of th Guadiana River, SW Iberia. Quaternary International, 221, 58–67.
Morton, R. A., Gelfenbaum, G., & Jaffe, B. E. (2007). Physical criteria for distinguishing sandy tsunami and storm deposits using modern examples. Sedimentary Geology, 2, 184–207. doi:10.1016/j.sedgeo.2007.01.003.
Moura, D., Albardeiro, L., Veiga-Pires, C., Boski, T., & Tigano, E. (2006). Morphological features and processes in the central Algarve rocky coast (South Portugal). Geomorphology, 81, 345–360. doi:10.1016/j.geomorph.2006.04.014.
Müller, H. J. (1991). Ökologie. Jena: Gustav Fischer.
Murray, J. W. (1979). British nearshore foraminiferids. In D. M. Kermack, R. S. K. Barnes (Eds.), Synopses of the British Fauna (New Series) (pp. 62). No. 16, Academic Press: London.
Murray, J. W. (1991). Ökologie. Jena: Gustav Fischer.
Murray, J. W. (2006). Ecology and applications of benthic foraminifera (pp. 426). Cambridge University Press, Cambridge, UK.
Nichols, M. M., & Biggs, R. B. (1985). Estuaries. In R. A. Davis Jr. (Ed.), Coastal Sedimentary Environments (pp. 77–186). Heidelberg: Springer.
Pint, A., Frenzel, P., Fuhrmann, R., Scharf, B., & Wennrich, V. (2012). Distribution of Cyprideis torosa (Ostracoda) in Quaternary athalassic sediments in Germany and its application for palaeoecological reconstructions. International Review of Hydrobiology, 97, 330–355.
Pozo, M., Ruiz, F., Carretero, M. I., Vidal, J. R., Cáceres, L. M., Abad, M., & González-Regalado, M. L. (2010). Mineralogical assemblages, geochemistry and fossil associations of Pleistocene-Holocene complex siliclastic deposits from the Southwestern Doñana National Park (SW Spain): A palaeoenviromental approach. Sedimentary Geology, 225, 1–18.
Queiroz, P. F., Mateus, J. E. (2004). Paleoecologia Litoral entre Lisboa e Sines. Do Tardiglaciário aos tempos de hoje. In A. A. Tavares, M. J. F. Tavares, J. L. Cardoso (eds.), Evolução Geohistórica do Litoral Português e Fenómenos Correlativos. Geologia, História, Arqueologia e Climatologia, Actas (pp 257-304). Lisboa Universidade Aberta.
Ramsey, C. B. (2001). Development of the radiocarbon calibration program. Radiocarbon, 43, 355–363.
Reyes, J. L., Martins, J. T., Benavente, J., Ferreira, Ó., Gracia, F. J., Dias, J. M. A., & López-Aguayo, F. (1999). Gulf of Cadiz beaches: A comparative response to storm events. Boletin Instituto Espanol de Oceanografia, 15, 221–228.
Ruiz Muñoz, F., González-Regalado Montero, M. L., & Morales González, J. A. (1996). Distribución y ecologíade los foraminíferos y ostrácodos actuales del estuario mesomareal del río Guadiana (SO España). Geobios, 29, 311–317.
Ruiz, F., Gonzalez-Regalado, M. L., Baceta, J. I., & Muñoz, J. M. (2000). Comparative ecological analysis of the ostracod faunas from low- and high-polluted southwestern Spanish estuaries: a multivariate approach. Marine Micropaleontology, 40, 345–376. doi:10.1016/S0377-8398(00)00048-7.
Ruiz, F., Rodríguez-Ramírez, A., Cáceres, L. M., Vidal, J. R., Carretero, M. I., Clemente, L., Muñoz, J. M., Yañez, C., & Abad, M. (2004). Late Holocene evolution of the southwestern Doñana National Park (Guadalquivir Estuary, SW Spain): a multivariate approach. Palaeogeography, Palaeoclimatology, Palaeoecology, 204, 47–64.
Ruiz, F., Rodríguez-Ramírez, A., Cáceres, L. M., Vidal, J. R., Carretero, M. I., Abad, M., Olías, M., & Pozo, M. (2005). Evidence of high-energy events in the geological record: Mid-holocene evolution of the southwestern Doñana National Park (SW Spain). Palaeogeography, Palaeoclimatology, Palaeoecology, 229, 212–229. doi:10.1016/j.palaeo.2005.06.023.
Ruiz, F., Abad, M., Bodergat, A. M., Carbonel, P., Rodríguez-Lázaro, J., González-Regalado, M. L., Toscano, A., García, E. X., & Prenda, J. (2013). Freshwater ostracods as environmental tracers. International Journal of Environmental Science and Technology, 10, 115–1128. doi:10.1007/s13762-013-0249-5.
Schlesinger, W. H. (1991). Biogeochemistry: An Analysis of Global Change. San Diego: Academic Press.
Schneider, H., Höfer, D., Trog, C., Busch, S., Schneider, M., Baade, J., Daut, G., & Mäusbacher, R. (2010). Holocene estuary development in the Algarve Region (Southern Portugal) - A reconstruction of sedimentological and ecological evolution. Quaternary International, 221, 141–158. doi:10.1016/j.quaint.2009.10.004.
Sliter, W. V. (Eds.) (1980). Studies in marine micropaleontology and paleoecology: a memorial volume to Orville L. Band. Washington, D.C: Cushman Foundation for Foraminiferal Research.
Soares, A. M. M., & Dias, J. A. (2006). Coastal upwelling and radiocarbon-evidence for temporal fluctuations in ocean reservoir effect off Portugal during the Holocene. Radiocarbon, 48, 45–60.
Soares, A. M. M., & Dias, J. A. (2007). Reservoir effect of coastal waters off western and northwestern Galicia. Radiocarbon, 49, 925–936.
Soares, A. M. M., & Martins, J. M. M. (2009). Radiocarbon dating of marine shell samples. The marine radiocarbon reservoir effect of coastal waters off Atlantic Iberia during Late Neolithic and Chalcolithic Periods. Journal of Archaeological Science, 36, 2875–2881. doi:10.1016/j.jas.2009.09.027.
Symposium on the Classification of Brackish Waters. (1958). The Venice System for the Classification of marine waters according to salinity. Oikos, 9, 311–312.
Teixeira, S. B. (2005). Evolução holocénica do litoral em regime transgressivo: o caso da costa de Quarteira (Algarve, Portugal). In: M. C. Freitas, T. Drago (Eds.) Iberian Coastal Holocene Paleoenviromental Evolution, Coastal Hope 2005 - Proceedings (pp. 121–124). Universidade de Lisboa.
Teixeira, S. B., & Pinto, C. A. (2002). Submarine Evidences of Holocene Shoreline Migration on Quarteira Coast (Southern Algarve-Portugal), Littoral 2002. Ed. Eurocoast – Portugal. Eurocoast Littoral The Changing Coast (pp. 411–416).
Teixeira, S. B., Gaspar, P., Rosa, M. (2005). Holocene sea-level index points on the Quarteira coast (Algarve, Portugal). In Iberian Coastal Holocene Paleoenviromental Evolution, Coastal Hope 2005 - Proceedings (pp. 125–127). Universidade de Lisboa.
Terrinha, P., Dias, R. P., Ribeiro, A., & Cabral, J. (1999). The Portimão Fault, Algarve Basin, South Portugal. Comunicações do Instituto Geológico e Mineiro, 86, 107–120.
Trog, C., Höfer, D., Frenzel, P., Camacho, S., Schneider, H., & Mäusbacher, R. (2013). A multi-proxy reconstruction and comparison of Holocene palaeoenvironmental changes of the Alvor and Alcantarilha estuaries (southern Portugal). Revue de Micropaleontologie, 56, 131–158. doi:10.1016/j.revmic.2013.10.003.
Vis, G.-J., & Kasse, C. (2009). Late Quaternary valley-fill succession of the Lower Tagus Valley, Portugal. Sedimentary Geology, 221, 19–39. doi:10.1016/j.sedgeo.2009.07.010.
Vis, G.-J., Kasse, C., & Vandenberghe, J. (2008). Late Pleistocene and Holocene palaeogeography of the Lower Tagus Valley (Portugal): effects of relative sea level, valley morphology and sediment supply. Quaternary Science Reviews, 27, 1682–1709. doi:10.1016/j.quascirev.2008.07.003.
Acknowledgements
This research was funded by the German Research Foundation (DFG; MA 1308/09). We thank Dr. Heike Schneider and Dana Höfer for supporting analyses and discussions, Dr. Gerhard Daut for access to the facilities of the laboratory of the Department of Physical Geography and the technicians of the sedimentological laboratory (Brunhilde Dreßler, Kati Hartwig) for analysing the geochemical parameters, PD Dr. Jussi Baade for measuring the exact positions of the core, Laura Pereira and Dr. Sarita Camacho for supporting identification of the microfauna and numerous students of the University of Jena for help during field work. The authors thank Dr. Simon Connor for the language corrections and the referees Prof. Alan Lord and Prof. Maria Cristina Cabral for their detailed and constructive reviews. This work is a contribution to IGCP Project 4959.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Trog, C., Hempel, R., Frenzel, P. et al. Holocene palaeoenvironmental changes in three lagoons on the Algarve coast of Portugal. Palaeobio Palaeoenv 95, 203–221 (2015). https://doi.org/10.1007/s12549-015-0185-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-015-0185-0