
Abstract
Besides the excavation of underground nest chambers, Atta vollenweideri grass-cutting ants build ventilation turrets on the topmost openings of their huge nests. Turret construction was studied in a laboratory colony, addressing the question whether turrets are simply heaps of disposed soil, or result from the import and a particular spatial arrangement of materials. The colony was daily offered different building materials, i.e., clay, coarse and fine sands, which workers collected and deposited around a nest opening to construct a turret. After 10 days, the spatial arrangement of the different building materials, offered either simultaneously or consecutively in independent experiments, was characterized via thin sections and micromorphological analysis of the turret’s walls. Workers did not select particular materials to be imported for turret building, but were selective in their spatial distribution and assembly into the turret structure. Particular types of microstructures were observed depending on the available materials, such as simple piles of sands, porous fabrics only composed of clay pellets, or fabrics with clay and sands combined. Turrets were very dynamic structures: while most imported materials were initially piled up at the shortest distance from the source, a marked subsequent material redistribution occurred as the turret grew, resulting in a new, mixed structure. Material re-assembly was particular evident when clay and sands were offered consecutively: a remarkable tendency to construct microstructures combining both materials was observed, i.e., clays were mobilized and included in sandy walls, and sands were intercalated in much compact clay walls. Irrespective of the materials used, walls showed a marked regular porosity in the range 50–60%, with the exception of secondary galleries that occasionally permeated the turret structure, which evinced lower porosity and therefore a more compact microstructure. Ants appeared to respond to local variations in the structural stability of the construction, since clay coatings smoothed and reinforced the surface of unstable sandy gallery walls. The observed building responses appear to be aimed at the maintenance of a porous yet mechanically-stable structure. The attained turret porosity may represent a compromise between high structural stability and low construction costs because of savings in material import.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Social insects construct nests that are regarded as adaptive structures, because they offer the colony inhabitants both protection and a long-term more stable microclimate (Korb 2003; Hansell 2005; Bollazzi et al. 2008). Among ants, colonies belonging to a wide range of subfamilies excavate nests in soil, which are organized in chambers big enough to hold food, brood items and the workers that tend them. While most underground ant nests result from the space generated by digging into the soil, a number of ant species adapt their nests to special needs by importing specific materials, digging under stones, or by constructing specific structures above ground (McCook 1877; Jonkman 1980a; Sudd 1982; Kleineidam et al. 2001; Bollazzi and Roces 2007)
Irrespective of the underground nest architecture, the members of an ant colony need to dispose of the excavated soil and prevent it to fall back into the structure. As a consequence, the formation of crater-like conical rings of soil around the nest openings represents the most economical way to dispose of the soil (Sudd 1982). In addition to the deposition of the excavated soil around nest entrances forming a so-called nest mound (Wehner 1970; Tschinkel and Bhatkar 1974; Tofilski and Ratnieks 2005; Robinson et al. 2008), a number of ant species transport materials from the surroundings to the nest surface (Smith and Tschinkel 2005), which may be used to stabilize the mound structure and provide protection from erosion (Whitford 2003), or to control the heat or humidity exchanges with the environment (Scherba 1962; MacKay and MacKay 1985; Bollazzi and Roces 2010a, c)
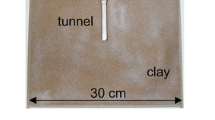
Leaf-cutting ants of the genus Atta excavate the largest and likely most complex nests among ants, with several thousands of underground chambers, mostly for rearing a symbiotic fungus on the plant fragments cut and carried by foragers, and for refuse (Jonkman 1980b; Moreira et al. 2004). Colonies of the grass-cutting ant Atta vollenweideri, occurring in the Chaco region of northern Argentina, build conspicuous conical- or ellipsoidal-shaped mounds uncovered of vegetation, commonly 6 to 8 m in basal diameter and about 0.80 m in high (Fig. 1). The mound does not solely result from a passive accumulation of the excavated soil, because workers import material to reinforce and stabilise the construction (Cosarinsky and Roces 2007), and also build structures on the top of several central nest openings that function as ventilation turrets (Jonkman 1980a; Kleineidam et al. 2001). A mature nest may contain up to 200 nest openings, most of them very large, with diameters up to 10 cm (Jonkman 1980b). The peripheral nest openings are used as entrances and connect to up to 100 m-long foraging trails, whereas the central nest openings, which have conspicuous turrets, do not function as entrances/exits for foragers (Jonkman 1980a, b).

a Nest of the leaf-cutting ant Atta vollenweideri in Formosa, Argentina; b Close-up of one ventilation turret from the nest shown above; c Ventilation turret removed from another field nest and held on the hand, to underline its structural stability; d Schematic side view of the nest arrangement for the study of turret construction in the laboratory, showing the two adjacent building platforms, the turrets built by the ants (vertical section) with the sources for building materials offered at each side (s: reddish clay on the left platform, dishes with sand grains on the right platform), the fungus chambers, refuse chamber and the foraging arena
The shape of the nest mound promotes the ventilation of the underground chambers via a passive mechanism driven by wind (Kleineidam and Roces 2000; Kleineidam et al. 2001). The turrets, because of their elevation beyond the mound surface, are expected to increase the pressure differences between their openings and the most basal ones at the peripheral nest surface, thus enhancing wind-driven nest ventilation (Kleineidam et al. 2001), like the windtowers built by human architects (Bahadori 1978). In fact, it has been shown that whereas the openings at the nest periphery behave as inflow conduits, the turrets located at the nest center act as outflow conduits (Kleineidam et al. 2001).
Almost nothing is known about the variables that motivate the construction of turrets, and the behavioral rules workers use for their construction. It is known that their height increases after heavy precipitation (the height of the turrets can reach 20–30 cm after rains), and that workers close the openings of several turrets in winter (Jonkman 1980a) and also after rains, re-opening them shortly thereafter (Kleineidam and Roces 2000). In our previous work, the micromorphology of the different parts of a large Atta vollenweideri nest was analysed and compared with that of the surrounding soil (Cosarinsky and Roces 2007). The turrets were composed of rounded soil pellets partially welded, forming a very porous wall. This structural feature was not so clear at the turret’s surface because of erosion by rain and wind, but it was evident in the infillings of recently-closed turrets. Pellets showed a composition similar to that of the surrounding soil, and originated from a deep soil horizon where most nest chambers are commonly excavated. Small fragments of twigs, grasses and leaves were in addition imported from the surroundings, and included among pellets in the wall of the mound and turrets, probably to reinforce the structure (Cosarinsky and Roces 2007).
The aim of the present study was to investigate the construction of ventilation turrets in the grass-cutting ant Atta vollenweideri by characterizing the assembly of different building materials using micromorphological analyses. Different experimental series, as described below, were performed to answer the main question whether turrets are simply heaps of collected material, or result from a particular arrangement of the available building materials. To answer this question, a laboratory nest was arranged so as to reproduce the spatial distribution of the main nest structures observed in natural nests. In independent experiments, workers were offered different building materials that they collected and deposited around a nest opening to construct a turret. Building materials, i.e., clay, coarse and fine sands, were offered singly or in combination, simultaneously or consecutively, and after 10 days, the spatial arrangement of the different building materials in the turret structure was characterized via thin sections, so as to investigate both the assembly and potential re-arrangement of materials depending on their availability. Further, artificial watering of one side of some of the turrets during the building process allowed the effects of rains on building behavior to be simulated and analysed. In a final experimental series, single workers were observed while collecting their loads, and the time needed to collect a load, as well as the relationship between load size and worker size were quantified, in an attempt to extrapolate individual behavior to the colony-wide building effort.
Materials and Methods
Laboratory Set-up
The colony of Atta vollenweideri (Forel) used in the experiments was collected in the Reserva Ecológica El Bagual, Formosa, Argentina, and reared at the Biozentrum, University of Würzburg, for 4 years in a room at 25°C and 50% relative humidity, under a Light:Dark cycle of 12 h:12 h. A single colony was used for the experiments because of its large size; small colonies seldom showed turret construction in the laboratory. The colony was fed mainly with leaves of privet (Ligustrum vulgaris) and dog rose (Rosa canina). The nest arrangement reproduced the spatial distribution of the main nest structures observed in natural nests (Jonkman 1980b; Cosarinsky and Roces 2007), with openings aimed at nest ventilation in the upper level, and nest chambers located at a lower level (Fig. 1d). The lower level consisted of three interconnected plastic boxes filled with ca. 1.5 l of fungus gardens each, one foraging arena and one refuse chamber, connected via transparent plastic tubes, 25 mm in internal diameter. The upper level consisted of two independent building platforms with vertical connections to the lower level. Each platform was covered with a Perspex box (56 cm × 56 cm × 56 cm; Fig. 1d), representing the “nest surface” and contiguous outside environment.
The colony was provided daily with fresh leaves placed in the foraging arena. Workers could move freely from the foraging arena into the first fungus chamber and subsequently to the next one. At the middle of the tube connecting two adjacent fungus chambers, a vertical Perspex tube (50 mm in internal diameter) allowed workers to gain access to the nest surface (one of the two building platforms) placed 40 cm above. Because of the slippery internal surface of the vertical tube, climbing of workers was facilitated by placing a wooden stick inside. The opening of the vertical tube, i.e., the access to the building platform, was reduced to a diameter of 15 mm by placing a ring of moist clay around the opening. During the experimental series, two sources of building materials were presented on these platforms, at both sides of the nest opening (Fig. 1d). Workers coming from the nest collected building materials and deposited them around the nest opening, forming a turret. Leaves were never offered as food on the building platforms, to avoid confusing behavioral responses in the contexts of foraging and building. This went also along with field observations: only the peripheral nest openings, and not the central ones, are used as entrances/exits during foraging. The experimental nest was maintained in a controlled room at 24°C, 80% relative humidity inside the building boxes, and under a Light:Dark cycle of 12 h:12 h.
Experimental Series
The rationale of the experimental design was to provide building materials ad libitum daily on each building platform. After 10 days, each experiment was finished and the structure of the turret built by workers was characterized via micromorphological analysis. For that, samples of the turret walls were carefully taken to preserve their structure, and impregnated with blue polyester resin (see below). Thin sections from the samples were later obtained for microscopic examination. The materials offered for turret construction were commercial clay used for house building (Claytec Baulehm gemahlen 0–0.5 mm, Germany), and commercial sands (Dorsilit Kristall-Quarzsand, Germany). Coarse and fine sand grains were 0.6–1.2 mm and 0.1–0.5 mm in diameter, respectively.
Eight different experiments were performed, as follows. Two initial experiments were aimed at characterizing the micromorphology of turrets built with single materials, either clay (Experiment A1) or coarse sands (Experiment A2). Four additional experiments investigated the microstructure of turrets constructed with two materials, clay and coarse sands (Experiment B1 and B2), or clay and fine sands (Experiment C1 and C2), to evaluate whether materials are evenly mixed in the turret structure, or workers re-arrange them. By offering the two materials either adjacent to each other (Experiments B1 and C1) or at the opposite side of the building platform (Experiments B2 and C2), it was also possible to examine whether the deposition of the collected materials depended on their initial spatial distribution, i.e., whether workers incorporate them into the turret at the shortest distance from the source, or whether workers re-arranged them over time. In addition, since workers are known to close nest openings or modify the turret structure after rains (Jonkman 1980a; Kleineidam and Roces 2000), the effects of artificial watering on the assembly of building materials was investigated. To that aim, one side of the turrets constructed in the experiments A1, B1, B2, C1 and C2 was watered once a day during the 6th and 7th experimental day. At the end of the experiments, the micromorphology of both the previously-watered and the dry turret sides was compared.
Two additional experiments explored the existence of behavioral flexibility in the incorporation of building materials into the turret structure. For that, workers were allowed to construct a turret with a single material for 5 days, and thereafter, the initial material was removed and replaced with an alternative one. In these two complementary experiments, either clay was provided over 5 days as single material and then replaced with coarse sands (Experiment D1), or vice versa (Experiment D2), i.e., while in the experiments B1 and B2 clay and coarse sand were offered simultaneously as described above, in the experiments D1 and D2 they were offered successively.
In a final experimental series offering single materials, performed independently to avoid interferences with the process of turret construction in the previous ones, 50 workers were observed while collecting their loads and the time needed to remove particles of the offered material, clay or sand, was recorded from the initial contact until the worker turned back with its load and started walking (henceforth called “loading time”). Immediately after the observed workers began to run loaded to the turret being constructed, they were collected together with their loads, and both ant body mass and load mass were determined at the nearest 0.01 mg.
Experimental Procedure
Each experiment began with the presentation of the building materials ad libitum, offered as single sources at both sides of the platform opening. Each building material was previously weighed and placed on a plastic dish, 20 cm to one side of the opening (Fig. 1d). Every day, the remaining building material was removed, weighed, and replaced with new one, in order to avoid desiccation. Clay and sands were presented as single masses (averaging 100 g), with clay having a water content of 20%, and sands of 18%. Two experiments were carried out simultaneously, one at each building platform. There was no evidence that they influenced each other, since workers carrying building materials repeatedly run back and forth from the material source to the turret, and were not observed to enter the nest and pass through a fungus chamber to reach the neighbouring platform. Therefore, the results of each experiment were presented and discussed independently. Detailed observations of the building behavior of individuals will be presented in a separate study.
The external morphology of the turrets was recorded twice a day with a photo camera. Experiments were finished typically after 10 days, when no further increase in the size of the constructed turrets was evident, and the collection of building materials markedly dropped. Finally, turrets were removed from the nest and dried at 40°C for 24 h, before sampling and impregnation with resin for micromorphological analysis.
Micromorphological Analysis
A total of 8 to 9 samples were collected from different parts of each single turret constructed, including the external and internal walls, taken from the top to the base of the turret, from both sides. The walls surrounding the principal shaft as well as the narrow secondary vertical shafts and galleries occasionally permeating the turret structure were also sampled. Samples of the experimentally-watered side of the turret were also collected and compared with those of the dry side. Thin sections (30 μm thick) were prepared after the impregnation of the samples with stained polyester blue resin, following a methodology commonly used for micromorphological analysis of soils (Murphy 1986), and observed under a petrographic microscope. The terminology employed to describe the microstructural types and micromorphological features was based on previous studies on termites and soils (Stoops 1964, 2003), but some new names are proposed herein to describe two so far non-described microstructural types (pelletal grain structure and spongy grain structure, see results). Porosity was visually estimated as a proportion of the void space, filled with blue resin, of the total area observed in the thin section, compared with graphics of abundance of black objects as a percentage of visual fields with various particle sizes (Fitzpatrick 1984; Bullock et al. 1985).
Results
Building Behavior and Turret Construction
The construction of a turret began with the collection and deposition of the offered building materials on the clay ring located around the opening leading to the nest interior. Workers were observed to pull out particles from the material source with their mandibles, and to also use their forelegs for the formation and collection of a load. Most workers deposited their loads at the shortest distance from the source. Other workers were observed to walk around the opening before unloading, or to stay and work on the emerging turret, i.e., to pick-up recently deposited loads and to redistribute them. During the first days of material collection and deposition, the initial turret became often conical-shaped like a crater, showing a single central vertical shaft connected to the nest entrance. But later on, as the structure grew, the shaft was sometimes closed, or it shifted laterally because of the asymmetrical deposition of material, frequently branching into several minor galleries (5 to 7 mm in diameter) that crossed through the turret wall and opened as small holes on the surface. Some conical turrets turned out to be domed over time, with a sub-circular or elongated base. Occasionally, a number of vertically-oriented, narrow secondary shafts developed around the initial opening, and some of them were eventually closed, as described for young field nests (Jonkman 1980a; Kleineidam and Roces 2000).
Building Materials and Turret Micromorphology
The morphology and microstructure of the turrets constructed with single materials, either clay or coarse sands, are shown in Fig. 2 (Experiments A1 and A2). The turret composed of clay (Fig. 2a and b) showed a “pelletal structure”, an open fabric with abundant, mammillated voids, formed by the clay loads that were observed as round aggregates or pellets composed of clay and silt, partially welded. Clay loads were named pellets because of their similarity with the micromorphological term employed for termite nests as “common micromorphological features of termite constructions recognized as spheroidal or ovoid bodies composed of organic and mineral soil materials or various combinations of both” (Sleeman and Brewer 1972). Pellets were all empty aggregates and similarly sized, ranging from 1 to 2.8 mm in diameter, exhibiting an internal, central or lateral void, and an external mammillated surface. Frequently, single pellets looked like curled chains composed of units of 0.5 to 1 mm (Fig. 2b). No particular coating was observed at the surface of the main shaft.

Experiments A1 and A2: Turrets built with single materials, either clay (a Experiment A1) or coarse sand (c Experiment A2). b Thin section of the clay turret showing the pelletal structure (pt, pellet; stars: voids, all in blue). d Close-up of one of the gallery openings constructed on the coarse sand turret, reinforced with clay pellets imported from a nearby location inside the nest. Inset: thin section of a reinforced gallery, at 2 cm depth from the nest surface, showing a smooth-edged coating of clay surrounding the aperture, indicated by the black arrow (same scale)
When offered solely coarse sands, ants had no other possibility than to build a turret consisting of piled loose grains, a “single grain structure”. However, the walls of several thin galleries were reddish and composed of sand cemented with clay (Fig. 2c and d, inset). The coated galleries were very thin and delicate, and easily collapsed during sampling. The source of the employed clay could only be the clay ring initially placed around the nest opening, thus indicating active removal of clay and its incorporation into the sandy structure, probably for mechanical stabilization. Some clay coatings were markedly smooth (Fig. 2d, inset), and many others showed mammillated borders, suggesting a pelletal origin and a posterior plastering. Interestingly, in some galleries the clay intercalation occurred immediately adjoining rather than on the surface of the gallery, forming so-called clay hypocoatings, which indicated the reworking of the surface after the deposition of clay.
When clay and coarse sands were offered simultaneously (Fig. 3, Experiments B1 and B2), the external morphology of the turrets initially depended of the spatial distribution of the offered materials, either adjacent or at the opposite sides of the platform (Fig. 3a and c). When the two materials were offered at opposite sides, the turret exhibited at the beginning an asymmetrical composition: mostly composed of sand at the side facing the sand source, and of clay close to the clay source. As the turret grew, however, this asymmetry gradually vanished and the surface was composed of both sand and clay (Fig. 3c). The resulting microstructure of the turret walls at the end of the two complementary experiments, particularly at the turret’s side facing the location of the sand source (Fig. 3c), was very similar in both cases, resulting in an intricate fabric of clay aggregates and sand grains, which is termed here “spongy grain structure”. This was a porous structure composed of abundant sand grains cemented with masses of clay, with voids variably shaped (Fig. 3b and d), indicating the active reorganization of the incorporated building material over time.

Experiments B1 and B2: Turrets built with clay and coarse sand offered simultaneously either adjacent (a Experiment B1) or at the opposite sides of the building platform (c Experiment B2). b Thin section showing the spongy grain structure. d Spongy grain structure as in (b), observed at the turret’s side facing the location of the sand source, indicated by the white arrow. Voids in blue
In turrets constructed with clay and fine sands (Fig. 4, Experiments C1 and C2), the composition of the external wall depended on the initial location of the materials. When the materials were offered adjacent to each other, the turret walls included clay mixed with sand grains (Fig. 4a). Such microstructure was termed here “pelletal grain”, and showed clusters of fine sands adhering to the pelletal fabric or filling the mammillated voids (Fig. 4b). When the materials were offered at opposite sides, however, a marked spatial redistribution occurred, with the turret surface being initially covered with fine sands, and finally re-covered with clay to some extent, resulting in a marble-like, reddish and white structure (Fig. 4c). At the most peripheral wall facing the sand source, the higher proportion of sand resulted in a “granular structure” with abundant granules composed of sands adhered to pellets or small clay aggregates (Fig. 4d).

Experiments C1 and C2: Turrets built with clay and fine sand offered simultaneously either adjacent (a Experiment C1) or at the opposite sides of the building platform (c Experiment C2). b Thin section showing the pelletal grain structure. d Granular structure, observed at the turret’s side facing the location of the sand source, indicated by the white arrow. Voids in blue
The dynamics of material rearrangement was evident in the experiments providing different materials consecutively, because of the final alternate composition of the turrets (Fig. 5, Experiments D1 and D2). The turret constructed in the first part of the experiment D1 only with clay (Fig. 5a) gradually included the sand grains offered the day 6th onwards, after removal of the available clay (Fig. 5b and c), and vice versa (Fig. 5e, f and g), i.e., workers included clay pellets in the sandy walls. The micromorphology of the inner wall of the turret showed the structure corresponding to the material initially available, i.e., either a “pelletal structure” or a “single grain structure” (as in Fig. 2), depending on whether clay or coarse sands were initially available. Interestingly, the turret initially built with clay (Experiment D1) also included some sand grains in the wall of its central shaft. Since sands were only available in the second part of the experiment, their incorporation in the shaft wall indicates that sand-carrying workers did not only drop and redistribute their loads on the turret surface, but also entered into the turret for unloading. The external turret walls, however, showed a similar “spongy grain structure” irrespective of the temporal sequence of material availability (Fig. 5d and h).

a Turrets built with both clay and coarse sand as materials, offered consecutively. In Experiment D1 (left), clay was offered singly for 5 days. At the 6th day, the remaining material was removed and replaced with coarse sand. The opposite schedule was used in Experiment D2 (right), i.e., first coarse sand and then clay were offered singly. d and h Spongy grain structure. Voids in blue
Most of the samples studied across the experiments showed a notable regular porosity around 50–60%. Comparisons with the porosity of the turret’s side artificially watered suggested that ants actively reworked the moist materials to keep porosity within this range. Watered walls exhibited a more compact structure in their interior, which corresponded to their previously-irrigated surface, as expected because of the breakdown of the porous structure. The original pelletal structure collapsed partially, showing porosities around 20%, as well as thin fissures or scattered micropores of less than 200 μ. However, the compact microstructure of the watered wall gradually changed outward to a porous microstructure, as workers reworked the moist material while continuing the construction.
In addition to its occurrence in watered walls, a porosity lower than 50–60% was also observed in walls of shafts and branching galleries, and in the infillings of several closed galleries. The infillings studied across the experiments showed a large micromorphological variation, even in different galleries of the same turret. Three types of infillings were observed in turrets composed of clay and sands: i) dense infillings displaying a similar structure to that of the adjacent wall but showing low porosity (20 to 40%), ii) loose and porous infillings composed of grains weakly adhered with thin clay bridges, or loose grains alternated with single clay pellets (porosity up to 60%), and iii) complex infillings composed of two distinct sectors displaying a massive structure at the perimeter, and a loose and porous structure centrally.
Forming the Building Units: Load Sizes and Loading Times
The relationship between the sizes of the loads collected from the different building materials ant the ant sizes is shown in Fig. 6. For clay and fine sands, there was a significant positive relationship between load wet mass (L) and ant mass (A), i.e., bigger ants collected larger loads, although the explanatory power of the equations was low (Clay: L = 0.762A + 0.543, r 2 = 0.21, F1,46 = 12.14, p < 0.01; Fine sand: L = 0.612A + 1.179, r 2 = 0.16, F1,62 = 11.98, p < 0.01). For coarse sands, there was no relationship between the variables (Coarse sand: L = 0.321A + 2.118, r 2 = 0.02, F1,65 = 2.28, p > 0.1, NS). To allow comparisons of load sizes, the median load masses were calculated for the body-size range 3.5–8 mg, i.e., for ants that were able to carry all three materials, resulting in 4.13 mg for clay, 4.15 mg for fine sand, and 3.29 mg for coarse sand, these values not being statistically different (Kruskal-Wallis H-test, H = 1.681, df = 2, p > 0.4, NS). Considering that the mean mass of a single dry grain was 0.048 ± 0.023 mg (N = 40) and 1.054 ± 0.499 mg (N = 119) for fine and coarse sands, respectively, it follows that a wet load (18% water content) of fine sand included in average 73 grains, whereas one of coarse sands consisted in average of 2.6, i.e., of 2 or 3 single grains.

Relationship between ant body mass and wet mass of the loads collected by workers from different sources of building materials: clay, fine sands, and coarse sands. For statistics see text
Interestingly, no small ants in the size range 1–3.5 mg were observed to collect loads of coarse sand (Fig. 6). Such small workers had difficulties in grasping coarse sands, and were unable to collect single sand grains. Based on data on body morphometry for this species: HW = 1.0085*BM0.3774, HW: Head width in mm, BM: Body mass in mg (Schelter 2009), it could be calculated that the head width of workers in the size class between 1 and 3.5 mg varied from 1.01 to 1.62 mm, which appear to limit the collection of coarse sand grains in the range 0.6–1.2 mm.
Regarding the time investment, workers in the range between 3.5 and 8 mg needed a median time of 72 s to form and collect a pellet of clay (n = 25), 23 s for a load of fine sand (n = 53), and only 8 s for a load of coarse sand (n = 67), these values being significantly different from each other (Kruskal-Wallis H-test, H = 87.5, df = 2, p < 0.001, after Dunn’s pairwise multiple comparisons).
Discussion
Turret Construction
Using a laboratory nest that reproduced the spatial arrangement of fungus chambers in relation to the openings leading to the nest surface, it was possible to stimulate workers to import building materials and to construct a ventilation turret on top of a nest opening. The combination of the experimental work with micromorphological studies revealed interesting details about the behavioral responses involved in turret construction. Several workers were observed to collect single loads from the material source, brought them to the turret being constructed and after dropping, ran back to the source for the next item. Other ants climbed the turret structure with their loads, and apparently searched for irregularities on the turret surface. They were observed to gently push and press the carried load so as to fit it into an available empty space or pore on the turret wall, a behavior also observed in field nests.
Considering the import of the three available materials as observed, it is tempting to ask whether workers showed preferences for any of them. Our experiments were not conceived as choice experiments, so that the question about material selection remains elusive. However, our measurements of individual loading times and total material consumption for turret building allow some inferences. The size of the collected loads was similar irrespective of the building materials, but the time needed to collect one markedly varied among them. Comparisons between the total amount of clay and coarse sands used to build a turret, in relation to the time invested in the collection of a single load, suggest that the workers’ decision to engage in load collection did not depend on the encountered material. This can be illustrated by comparing the ratios between the times needed to collect a single load of coarse sand and clay, and the ratio between the total sand and clay amounts collected over 10 days for turret building, as follows. It took a worker nine times longer to form and collect a clay pellet than one load of coarse sand (72 s for clay, 8 s for coarse sand). Under the realistic assumption that single workers need ca. 1 min for running to the material source and back, it follows that a collecting cycle for a clay pellet lasts 132 s, and one for a sand load 68 s in average. As a consequence, turrets constructed with clay and coarse sands offered simultaneously should consist of roughly the double amount of sand than of clay, assuming that both materials have the same probability to be found by workers, and that workers transport them at the same rate. Deviations from this ratio may indicate that the probability of a worker to collect a load is influenced by the material itself. The ratio between coarse sand and clay incorporation in the final turret measured in our experiments averaged 2.1, which suggests no preference for any of the materials offered. In Temnothorax ants dwelling in rock crevices, experiments suggest the existence of preference for larger sand grains for the construction of nest walls, although workers also collect smaller grains to build mixed structures (Aleksiev et al. 2007a, b). Termites do indeed prefer particles from particular soil types for the construction of specific parts of the nest structure (Jouquet et al. 2002).
How strong the behavioral response of importing materials for turret construction was became evident even in between experiments, when no materials were offered. Workers were observed to accumulate leaf fragments and twigs around the nest openings, which they cut and carried from the foraging arena, i.e., they needed to walk loaded through the fungus chambers to reach the nest surface on the building platforms.
During the construction of the turret, most workers were observed to deposit the collected building materials at the shortest distance from the source, initially piling them on the surface of the emerging turret. However, a marked posterior movement and redistribution of the materials occurred, indicating that the turret was a fairly dynamic structure. This fact was evident in those experiments in which clay and sands were offered at the opposite sides of the opening. While the composition of the initial turret was remarkably asymmetric, sandy in front of the sand source and clayish in front of the clay source, the asymmetry disappeared as the turret grew, because of the redistribution and combination of materials as mentioned above. A similar phenomenon occurred when clay and sands were offered consecutively. When clay was available at the beginning, and later replaced with sands, the clay walls deeply included sands among the pellets at the end of the experiments. Both materials were combined in the turret structure, showing a new, mixed surface. In a similar way, sandy walls deeply included clay pellets among the grains when the offered sand was replaced with clay. Likewise, turrets built with clay and sands offered at opposite sides were initially strong asymmetric, and a gradual movement and redistribution of clay and sands was observed over time, leading to a final, rather homogeneous or marble-like surface. Only in shorts periods of time during the development of the turret constructed with clay and fine sands, a distinct sand layer almost covered the whole clay side of the turret, but it finally disappeared because sands were moved and redistributed forming a new mixed fabric combined with clay.
What variables trigger the collection of building materials and spatially guide their deposition around nest openings? Workers may respond to the step gradients of humidity and respiratory gases that are expected to occur at nest entrances, and cluster pellets and objects there as known for several ant species (McCook 1879; Jonkman 1980a; Smith and Tschinkel 2005; Bollazzi and Roces 2007; Ribeiro and Navas 2008). In fact, it has been experimentally demonstrated that leaf-cutting ant workers counteract humidity losses from the nest by collecting building materials and by depositing them around the nest openings (Bollazzi and Roces 2010b). In Acromyrmex heyeri, the observation of an active search for building materials indicate that humidity losses trigger in fact a building response, and not merely the dropping of objects being transported as a direct reaction to the perceived stimulus (Bollazzi and Roces 2010a). Soil moisture may also stimulate turret building, likely because excavation is facilitated, and also because the damped soil pellets easily adhere on each other. In fact, Atta vollenweideri workers build and enlarge their nest turrets particularly after rains (Jonkman 1980a), but turret construction also occur in dry periods. Whether workers’ building responses are directly influenced by airflow velocities around the opening or through the tunnels, as known for termites (Howse 1966), remains to be investigated.
As already mentioned, the ventilation turrets constructed by Atta vollenweideri are particularly large, and no other Atta species is known to construct such structures that improve ventilation, although several species cluster materials around the nest openings or even close them during rains, colony inactivity, or after the swarming flights (Autuori 1947; Weber 1972). Turret construction for nest ventilation in A. vollenweideri may have been favoured during evolution because of the marked low porosity of the clay soils where the nests occur, which strongly reduces, or may even preclude, diffusive air exchanges between the nest and the atmosphere through the soil’s structure (Currie 1984). Small turrets upon nest entrances are known in few other fungus-growing ant species, but in these cases, they are unlikely to promote nest ventilation because of their size. Nests of Acromyrmex landolti have a single small turret that has been shown to protect the colony against flooding (Navarro and Jaffé 1985; LeBrun et al. 2011). Even the small colonies of Trachymyrmex turrifex, consisting of only two or three dozen individuals, build a single cylindrical turret with a height of 1 to 4 cm (Wheeler 1907), the function of which is unknown.
Building Materials and Turret Micromorphology
By comparatively characterizing the turret’s micromorphology across the experiments, we distinguished different types of microstructures depending on the material employed. When only coarse sands were offered, the turret showed a “single grain structure” and evinced a striking stability. It was only composed of piled sand grains fitted by their faces like the rock blocks used in the pre-Columbian Peruvian constructions of Machu Picchu, in which no cement was employed.
When only moist clay was offered, the turret showed a “pelletal structure”, composed of an intricate porous net of clay pellets that almost entirely fitted one each other because of two reasons: first, the plasticity of the moist clay, and second, the presence of mammillated surfaces that tightly fastened upon each other even without cement. A similar shape of the pellets was described for Formica mound-building ants. Pellets were described as “accumulations of small particles of soil, united in various irregular shapes, by the pressure of the mandibles” (McCook 1877). The author observed that “the irregular faces of the pellets fit into and fasten upon each other”, and concluded that “whether or not these pellets be wrought into their peculiar shapes with deliberate and intelligent purpose; or are only an accident of their preparation or handling, it is obvious that their form must greatly facilitate the work of the ant in fastening them together”. In nests of Solenopsis invicta fire ants, soil pellets were described as curled aggregates, suggesting that rain-dampened soil might be excavated by the worker’s mandible in much the same way that ice-cream is excavated by an ice-cream scoop (Cassill et al. 2002). Agreeing with this description, the pellets of Atta vollenweideri are also hollow and voluminous aggregates, light and very easy to be carried by workers (Cosarinsky and Roces 2007). Thus, a pelletal construction seems to be very adequate to build dynamic structures, allowing a posterior reorganization of the employed materials.
The micromorphology of the turrets constructed with clay and sands offered simultaneously revealed two types of microstructures, depending on the sand granulometry. In walls constructed with coarse sands and clay, the sand grains were cemented by small clay masses forming a porous fabric (spongy grain structure), whereas in walls constructed with fine sands and clay, clusters of fine sands adhered to the clay or filled pores of a pelletal fabric (pelletal grain structure). These microstructural types suggest the involvement of two different building behaviors when clay was combined with coarse or fine sands. When coarse sands were available, workers constructed the turret mostly linking one or two coarse sand grains with the clay masses of pelletal origin, whereas when fine sands were available, they constructed a pelletal fabric and then placed packs of fine sands on the pelletal surface and in existing pores. Our measurements of individual load sizes and number of sand grains per single load are consistent with these arguments.
The Dynamics of Turret Building: Maintenance of a Porous Yet Mechanically-Stable Structure?
A notable tendency to attain the microstructural types mentioned above was evident after the micromorphological analysis of turrets constructed with clay and sands that were offered consecutively. In the sandy turret constructed only with coarse sands, the initial single grain structure deeply modified to a new spongy grain structure composed of sand grains cemented with small clay masses, after the offered sand was replaced with clay. That is, workers mobilized clay and included the pellets in the less-porous, sandy walls. On the contrary, in turrets constructed only with clay at the beginning, the original pelletal structure deeply modified to a new spongy grain structure after the sand replaced the clay offered for building, in which grains were cemented with clay masses of clay removed from the pelletal fabric. These results indicate that workers do not simply pile clay over sands or sands over clay after replacement of the available materials, and evince some complexity in construction behavior. The micromorphological analysis of the final wall demonstrated that the imported materials were distributed and combined tending toward a kind of microstructural balance that may be related to the maintenance of a porous but mechanically-stable structure.
The turret porosity was notably invariant in all the experiments. It ranged from 50 to 60%, independent of the building materials used, and it was very similar to that observed in samples taken from turrets of field nests (Cosarinsky and Roces 2007). The ants’ building responses aimed at attaining this porosity range were in addition revealed by comparing the porosity of the initially-irrigated turret surface with that of the following construction above the watered surface. The compact microstructure of the watered wall gradually changed to a porous microstructure in the reworked turret surface, returning to porosity values in the range 50 to 60%.
The view that the observed porosity range is not simply the necessary consequence of the spatial arrangement of the pellets is additionally supported by the examination of more unstable structures. The wall of narrow shafts and branching galleries showed lower porosity values than the rest of the turret, revealing an active behavior aimed at reinforcing those structurally-unstable regions, protecting them from collapsing. In addition, the surface of many galleries in turrets composed of clay and coarse sands were reinforced and smoothed with compact clay coatings, while no coatings were observed at the surface of the main, more larger shafts, as reported for field nests (Cosarinsky and Roces 2007). This indicate that ant workers are able to respond to local variations in the mechanical stability of the nest structure, as also suggested in a recent study on tunneling behavior of ants (Espinoza and Santamarina 2010), although the question of how workers sense it remains elusive. Responses to mechanical instabilities appear to be a widespread phenomenon in ants, since galleries of Camponotus punctulatus nests are plastered with a distinct soil coating (Cosarinsky 2006), and the compactness of the gallery walls appears to be necessary to stabilise the structure and to support the high mass of the nest mound (Cosarinsky 2006; Gorosito et al. 2006). Coating of nest walls has also been reported for other ant species inhabiting sandy soils (Dejean and Lachaud 1994; Wang et al. 1995).
Taken together, our study demonstrates that ventilation turrets in grass-cutting ant nests are built structures that do not simply result from a passive deposition of the excavated soil. The building effort is large: for a turret of average size constructed over 10 days, workers imported ca. 600 g of material, which corresponds to a total of 15,000 loads (4 mg each) transported per day. Turrets are dynamic structures that change in shape and arrangement of their constitutive materials: building materials are imported by workers and initially piled around the nest entrance, being re-distributed to form particular types of microstructures. Workers do not appear to select particular materials to be imported for turret building, but are selective in their spatial distribution and assembly into the turret structure. The substrate properties are known to influence ant digging behavior and so the shape of the excavated structures (Espinoza and Santamarina 2010; Toffin et al. 2010). Our results go beyond by showing that the substrate properties spatially guide workers’ responses in the context of building, leading to material re-arrangement and to the construction of structures with a particular morphology. The observed dynamics in material arrangement appears to be aimed at the maintenance of a porous yet mechanically-stable structure. The attained turret porosity may represent a compromise between high structural stability and low construction costs because of savings in material import. In Atta vollenweideri, a structurally-stable turret is needed to improve nest ventilation, and also to prevent the inflow of runoff during rainy periods, as known for other ant species (Navarro and Jaffé 1985; LeBrun et al. 2011). A porous structure may be advantageous to allow its rapid deconstruction as soon as a nest opening needs to be closed during rains, and to allow gas exchanges with the outside when the turret remains closed after heavy rains or in winter (Jonkman 1980a; Kleineidam and Roces 2000).
References
Aleksiev AS, Longdon B, Christmas MJ, Sendova-Franks AB, Franks NR (2007a) Individual choice of building material for the nest construction by worker ants and the collective outcome for the colony. Anim Behav 74:559–566
Aleksiev AS, Sendova-Franks AB, Franks NR (2007b) The selection of building material for wall construction by ants. Anim Behav 73:779–788
Autuori M (1947) Contribuição para o conhecimento da saúva (Atta spp. Hymenoptera–Formicidae). IV. O saúveiro depois da primeira revoada (Atta sexdens rubropilosa Forel, 1908). Inst Biol 18:39–70
Bahadori MN (1978) Passive cooling systems in Iranian architecture. Sci Am 238:144–154
Bollazzi M, Kronenbitter J, Roces F (2008) Soil temperature, digging behaviour, and the adaptive value of nest depth in South American species of Acromyrmex leaf-cutting ants. Oecologia 158:165–175
Bollazzi M, Roces F (2007) To build or not to build: circulating dry air organizes collective building for climate control in the leaf-cutting ant Acromyrmex ambiguus. Anim Behav 74:1349–1355
Bollazzi M, Roces F (2010a) Control of nest water losses through building behavior in leaf-cutting ants (Acromyrmex heyeri). Insectes Soc 57:267–273
Bollazzi M, Roces F (2010b) Leaf-cutting ant workers (Acromyrmex heyeri) trade off nest thermoregulation for humidity control. J Ethol 28:399–403
Bollazzi M, Roces F (2010c) The thermoregulatory function of thatched nests in the South American grass-cutting ant Acromyrmex heyeri. J Insect Sci 10:137, available at: insectscience.org/10.137
Bullock PN, Fedoroff A, Jongerius G, Stoops G, Tursina T (1985) Handbook for soil thin section description. Waine Research Publications, Albrighton
Cassill DL, Tschinkel WR, Vinson SB (2002) Nest complexity, group size and brood rearing in the fire ant, Solenopsis invicta. Insectes Soc 49:158–163
Cosarinsky M (2006) Nest micromorphology of the neotropical mound building ants Camponotus punctulatus and Solenopsis sp. Sociobiol 47:329–344
Cosarinsky M, Roces F (2007) Neighbor leaf-cutting ants and mound-building termites: comparative nest micromorphology. Geoderma 141:224–234
Currie JA (1984) Gas diffusion through soil crumbs: the effects of compaction and wetting. J Soil Sci 35:1–10
Dejean A, Lachaud J-P (1994) Ecology and behavior of the seed-eating ponerine ant Brachyponera senaarensis (Mayr). Insectes Soc 41:191–210
Espinoza DN, Santamarina JC (2010) Ant tunneling—a granular media perspective. Granular Matter 12:607–616
Fitzpatrick EA (1984) Micromorphology of soils. Chapman and Hall, New York
Gorosito NB, Curmi P, Hallaire V, Folgarait PJ, Lavelle PM (2006) Morphological changes in Camponotus punctulatus (Mayr) anthills of different ages. Geoderma 132:249–260
Hansell MH (2005) Animal architecture. Oxford University Press, Oxford
Howse PE (1966) Air movement and termite behaviour. Nature 210:967–968
Jonkman JCM (1980a) The external and internal structure and growth of nests of the leaf-cutting ant Atta vollenweideri Forel, 1893 (Hym.: Formicidae). Part I. Z Ang Ent 89:158–173
Jonkman JCM (1980b) The external and internal structure and growth of nests of the leaf-cutting ant Atta vollenweideri Forel, 1893 (Hym.: Formicidae). Part II. The internal nest structure and growth. Z Ang Ent 89:217–246
Jouquet P, Lepage M, Velde B (2002) Termite soil preferences and particle selections: strategies related to ecological requirements. Insectes Soc 49:1–7
Kleineidam C, Ernst R, Roces F (2001) Wind-induced ventilation of the giant nests of the leaf-cutting ant Atta vollenweideri. Naturwissenschaften 88:301–305
Kleineidam C, Roces F (2000) Carbon dioxide concentrations and nest ventilation in nests of the leaf-cutting ant Atta vollenweideri. Insectes Soc 47:241–248
Korb J (2003) Thermoregulation and ventilation of termite mounds. Naturwissenschaften 90:212–219
LeBrun EG, Moffett M, Holway DA (2011) Convergent evolution of levee building behavior among distantly related ant species in a floodplain ant assemblage. Insectes Soc 58:263–269
MacKay WP, MacKay EE (1985) Temperature modifications of the nest of Pogonomyrmex montanus (hymenoptera: formicidae). Southwest Nat 30:307–310
McCook HC (1877) Mound-making ants of the Alleghenies, their architecture and habits. Trans Amer Ent Soc 6:253–296
McCook HC (1879) On the architecture and habits of the cutting ant of Texas (Atta fervens). Ann Mag Nat Hist 3:442–449
Moreira AA, Forti LC, de Andrade APP, Boaretto MAC, Lopes JFS (2004) Nest architecture of Atta laevigata (F. Smith, 1858) (Hymenoptera: Formicidae). Stud Neotrop Fauna Environ 39:109–116
Murphy CP (1986) Thin section preparation of soils and sediments. A. B. Academic Publishers, London
Navarro JG, Jaffé K (1985) On the adaptive value of nest features in the grass-cutting ant Acromyrmex landolti. Biotropica 17:347–348
Ribeiro PL, Navas CA (2008) Colony dehydration and water collection by specialized caste in the leaf-cutting ant Atta sexdens rubropilosa. J Insect Behav 21:549–558
Robinson EJH, Holcombe M, Ratnieks FLW (2008) The organization of soil disposal by ants. Anim Behav 75:1389–1399
Schelter M (2009) Energetik des Grabverhaltens von Blattschneiderameisen. In: Diploma-Thesis, Fakultät für Biologie, Julius-Maximilians-Universität Würzburg, Germany, pp 48
Scherba G (1962) Mound temperatures of the ant Formica ulkei emery. Am Midl Nat 67:373–385
Sleeman JR, Brewer R (1972) Micro-structure of some Australian termite nests. Pedobiol 12:347–373
Smith CR, Tschinkel WR (2005) Object depots in the genus Pogonomyrmex: exploring the “who”, what, and where. J Insect Behav 18:859–879
Stoops G (1964) Application of some pedological methods to the analysis of termite mounds. In: Bouillon A (ed) Études sur les termites Africains. Edition de l’Université, University of Leopoldville, Leopoldville, pp 379–398
Stoops G (2003) Guidelines for analysis and description of soil and regolith thin sections. Soil Science Society of America, Madison
Sudd JH (1982) Ants: foraging, nesting, brood behavior, and polyethism. In: Hermann HR (ed) Social insects, vol IV. Academic, New York, pp 107–155
Toffin E, Kindekens J, Deneubourg JL (2010) Excavated substrate modulates growth instability during nest building in ants. Proc R Soc Lond B 277:2617–2625
Tofilski A, Ratnieks FLW (2005) Sand pile formation in Dorymyrmex ants. J Insect Behav 18:505–512
Tschinkel WR, Bhatkar A (1974) Oriented mound building in the ant, Trachymyrmex septentrionalis. Environ Entomol 3:667–673
Wang D, McSweeney K, Lowery B, Norman JM (1995) Nest structure of ant Lasius neoniger emery and its implications to soil modification. Geoderma 66:259–272
Weber NA (1972) Gardening ants—the attines. The American Philosophical Society, Philadelphia
Wehner R (1970) Ètudes sur la construction des cratères au-dessus des nids de la fourmi Cataglyphis bicolor (Hymenoptera, Formicidae). Insectes Soc 17:83–94
Wheeler WM (1907) The fungus-growing ants of North America. Bull Am Mus Nat Hist 23:669–807
Whitford WG (2003) The functional significance of cemented nest caps of the harvester ant, Pogonomyrmex maricopa. J Arid Environ 53:281–284
Acknowledgements
Special thanks are due to Annette Laudahn, Martin Bollazzi and Oliver Geissler for valuable help during the experiments. We are also indebted to Alejandro G. Di Giacomo and the Götz family for providing facilities at the Reserva Ecológica El Bagual (Alparamis SA—Aves Argentinas) in Formosa, Argentina, where preliminary field experiments were performed. This study was performed at the Univ. of Würzburg during a research visit of the first author, supported by funds from the German Research Foundation (DFG, grant SFB 554/TP E1).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Cosarinsky, M.I., Roces, F. The Construction of Turrets for Nest Ventilation in the Grass-Cutting Ant Atta vollenweideri: Import and Assembly of Building Materials. J Insect Behav 25, 222–241 (2012). https://doi.org/10.1007/s10905-011-9290-8
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10905-011-9290-8