
Abstract
The availability of different forages varies across landscapes, but little is known about how proportional consumption of different foods affects food intake and the use of landscapes. The relative amounts of nutrients and plant secondary metabolites (PSM) consumed by herbivores may influence forage intake and animal fitness in landscapes dominated by plants with PSM. Our objective was to determine if the relative availability of nutritious foods that contain PSM affected food preference and intake by lambs. Lambs in three treatments (eight lambs/treatment) were offered two PSM-containing foods at different relative availabilities: treatment 1—ad libitum terpene- and 100-g tannin-containing food, treatment 2—ad libitum tannin- and 100-g terpene-containing food, and treatment 3—ad libitum access to both tannin- and terpene-containing foods. We measured intake of individual foods and total intake of PSM-containing foods during conditioning and a preference test, where animals were offered both PSM-containing foods ad libitum. When lambs were fed 100 g of the tannin-containing food, they ingested more terpene and total PSM (P < 0.05) than when both PSM were offered ad libitum, but limiting terpenes did not affect intake of tannin or total intake of PSM (P > 0.10). During preference tests, all groups preferred tannins over terpenes, but lambs in the treatment fed 100 g of tannin had a higher preference for terpenes than did lambs in the other two treatments (P < 0.10). These results support the notion that the relative amounts of PSM consumed affects intake and preference for PSM-containing foods by herbivores.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
All plant species contain plant secondary metabolites (PSM) that limit herbivory and can be toxic at high doses (Cheeke and Schull 1985; Foley et al. 1999). To maintain health, grazing animals must regulate intake of PSM. The variable degree to which herbivores cope with PSM creates differential use of plant species, potentially shifting the balance of competition among plant species in ways that can change ecosystem structure (Council for Agriculture Science and Technology 2002; Villalba et al. 2004). As ecosystems become dominated by invasive plants high in PSM, alternative forages decline because of increased competition and grazing pressure. Ever-decreasing plant diversity further compromises the ability of animals to regulate intake of PSM and meet demands for nutrients (Dearing and Cork 1999; Burritt and Provenza 2000; Villalba et al. 2004), which further increases grazing pressure on palatable plants (Provenza et al. 2003a).
Liver biotransformation and rumen degradation both help herbivores ingest plants with terpenes and tannins. In the liver, PSM are processed through two different detoxification pathways (Freeland and Janzen 1974; Dearing and Cork 1999; Burritt and Provenza 2000; Villalba et al. 2004). In mammals, biotransformation of PSM is carried out in the liver and usually occurs in two steps. The first step (phase I) introduces a reactive group—such as OH, NH2, COOH, or SH—into the structure of the PSM such that the PSM is made more hydrophilic and polar. During the second step (phase II), the newly formed compound is conjugated with endogenous molecules or groups—glucuronic acid, amino acids, sulphates, or methyl groups—that are hydrophilic (Osweiler et al. 1985). PSM such as terpenes are processed primarily via the phase I pathway, whereas tannins are processed by the phase II pathway (Sipes and Gandolfi 1986). Condensed tannins are large molecules normally not absorbed in the gastrointestinal tract; some are degraded and absorbed to a degree, yielding postabsorptive (potentially toxic) effects (Mehansho et al. 1987; Clausen et al. 1990). Finally, diverse microbial populations in the foregut also are important for PSM biotransformation in ruminants. Terpenes (Johnson et al. 1976; Dizba et al. 2006) and tannins (Foley et al. 1999) both are detoxified by microbes in the rumen.
Previous research suggests that total intake of PSM is higher when tannins and terpenes are fed together rather than when fed in isolation (Villalba et al. 2004; Mote et al. 2007), which suggests that the relative amounts of PSM in a landscape play a key role in how herbivores use forages. Postingestive feedback responses likely are sensitive enough to regulate proportional intake of different PSM (Marsh et al. 2006), as they do with protein and energy (Villalba and Provenza 1999; Provenza and Villalba 2006).
Multiple food alternatives increase sheep intake of foods that contain PSM (Burritt and Provenza 2000; Villalba et al. 2004), perhaps because PSM have different effects in the body and are detoxified by different mechanisms (Freeland and Janzen 1974). Most PSM satiate herbivores (Provenza et al. 2003b); once a threshold of PSM intake is reached, an animal will stop eating the food with that PSM until detoxification is achieved (Foley et al. 1999; Wiggins et al. 2006a, b, c). Satiation on specific PSM in foods may create mild to strong food aversions that decrease subsequent intake and preference for those foods (Provenza 1996). Thus, animals exposed too frequently or to too large a quantity of a specific PSM will satiate on food with that PSM, decreasing their preference relative to an alternative PSM-containing food.
We compared consumption of two PSM-containing foods that varied in availability. We hypothesized that the relative abundance of the two PSM-containing foods—tannins and terpenes—would influence the total amount of food consumed as well as preference for each food. We predicted lambs would decrease their preference for the PSM available ad libitum and that PSM consumed below satiation thresholds would be preferred, as they were not eaten in amounts high enough to promote aversions.
Methods and Materials
The experiment was conducted with weaned commercial crossbred lambs, averaging 35 kg (SEM = 1.09), at the Mote Farm in Willard, UT (NAD 83 UTM 12 T 0413040 4583816). Lambs were individually penned with free access to trace mineralized salt blocks and fresh water. All procedures were approved by the Utah State University Institutional Animal Care and Use Committee.
Test Foods
Test foods were formulated to be similar in PSM and nutrient contents to plants found in the sagebrush steppe ecosystem. We used two isocaloric (2.9 Mcal digestible energy/kg dry matter [DM]) and isonitrogenous (126 g crude protein/kg DM; NRC 1985) test foods that contained either tannins or terpenes, similar to those described by Villalba et al. (2004). The tannin-containing food (dry weight) was 76% beet pulp, 9% soybean meal, and 15% quebracho tannin powder. The terpene-containing food (dry weight) was 52.8% beet pulp, 26% grape pomace, 14% soybean meal, and 7.2% of a terpene solution where camphor (18.2 mg/g diet), 1–8 cineole (12.0 mg/g diet), and p-cymene (0.7 mg/g diet) were dissolved in 4.1% of vegetable oil. Terpene volatilization was reduced by mixing terpenes with vegetable oil, which also improved terpene homogeneity in the diet (Kimball et al. 1998). All ingredients were ground to 2- to 3-mm particle sizes to facilitate mixing.
Conditioning
Twenty-four lambs were stratified by weight and randomly assigned to three treatments (eight lambs/treatment) that differed in the amounts of the tannin- and terpene-containing foods they received: treatment 1—terpene food ad libitum, 100 g of the tannin food; treatment 2—tannin food ad libitum, 100 g of the terpene food; treatment 3—tannin and terpene foods ad libitum. One lamb in treatment 1 was removed from the study because it consistently refused to eat any of the test foods. All lambs were offered both foods simultaneously for 4 hr, beginning at 06:00 each morning, for 12 d. Alfalfa pellets were offered at 10:00 for the first 3 d, at 500, 300, and 150 g lamb−1 d−1, respectively, to ensure lambs maintained ruminal health while being introduced to the novel foods. After day 3, all lambs were fed only their respective diets.
Preference Tests
After conditioning, all animals were fed alfalfa pellets ad libitum from 06:00 to 10:00 for five consecutive days to allow for toxin elimination. The retention time of quebracho tannin in the gastrointestinal tract ranges from 48 (free and soluble) to 72 hr (bound to protein and fiber; Silanikove et al. 1994, 1996). The average primary and terminal elimination half-lives of plasma terpene concentrations is on the order of minutes after terpenes are infused into the rumen or blood stream (Dizba et al. 2006).
We wanted to determine lambs’ preferences for tannin- and terpene-containing foods as a function of their prior experience during conditioning, so we offered all lambs a choice between the terpene- and tannin-containing foods for 30 min/d on three consecutive days immediately after the 5-d toxin elimination period. The diet fed during preference tests was the same diet lambs in treatment 3 were fed during conditioning. After preference tests, lambs were fed alfalfa pellets until 10:00, when refusals were collected and no other food was offered until the next day.
Statistical Analyses
Data were analyzed as a split-plot design with three treatments (ad libitum terpenes, restricted tannins; ad libitum tannins, restricted terpenes; ad libitum tannins and terpenes) as the whole-plot factor; lambs were nested within treatments. Day was the subplot. Intake of individual foods and total intake of PSM-containing foods were the response variables for both the conditioning and preference trials. Refusals were subtracted from the amount of food offered to calculate grams of food ingested per kilogram body weight (BW). Preference was computed as: [intake of a single-toxin containing food/total intake of toxin-containing foods] × 100. Analyses of variance (ANOVAs) were completed by using the MIXED procedure in SAS 9.1 for Windows (SAS, Littell et al. 1996). Data were analyzed with standard procedures for unbalanced ANOVA because of the removal of a lamb from treatment 1. Assumptions of normality and homogeneity of variance were met in normal probability plots and plots of residuals against predicted values.
Results
Conditioning
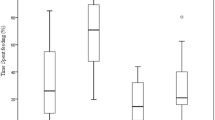
Averaged across the 12 d of conditioning, lambs in treatment 1 that were offered a restricted amount of tannins and ad libitum amounts of terpenes ate more terpene-containing food (17.4 g/kg BW; SEM = 2.1) than lambs in treatment 3 that were offered both the tannin- and the terpene-containing foods ad libitum (5.3 g/kg BW; SEM = 2.0; P < 0.05, Fig. 1). Intake of tannins by lambs in treatment 2 fed ad libitum amounts of tannins and restricted amounts of terpenes (8.7 g/kg BW; SEM = 1.14) did not differ from that of lambs in treatment 3 (9.4 g/kg BW; SEM = 1.14; P > 0.05, Fig. 1). Averaged across days, lambs in treatment 1 ate more total PSM-containing food (20.2 g/kg BW; SEM = 1.7) than lambs in treatments 2 (10.6 g/kg BW; SEM = 1.6) or 3 (14.7 g/kg BW; SEM = 1.6; treatment effect P < 0.001; Fig. 1). By day 9, ten lambs had diarrhea—five each in treatments 2 and 3. After lambs were offered alfalfa pellets for 5 d, all signs of diarrhea disappeared.

Daily intake of terpene- and tannin-containing foods by three groups of lambs during 12 d of conditioning. Lambs in treatment 1 (ad libitum terpenes, restricted tannins), were offered the terpene-containing food ad libitum and only 100 g of tannin-containing food. Lambs in treatment 2 (ad libitum tannins, restricted terpenes) were offered the tannin-containing food ad libitum and only 100 g of the terpene-containing food. Lambs in treatment 3 were offered both the tannin- and the terpene-containing foods ad libitum
Preference Tests
When offered a choice, lambs in all treatments preferred the tannin- to the terpene-containing food, but the differences were less pronounced for treatment 1 (ad libitum terpenes, restricted tannins: 59%, P < 0.15) than for treatments 2 (ad libitum tannins, restricted terpenes: 70%, P < 0.10) and 3 (ad libitum tannins and terpenes: 79%, P < 0.05). Although all lambs preferred tannins to terpenes, lambs in treatment 1 ate more terpene food than lambs in treatments 2 or 3 (P < 0.10; Fig. 2). Lambs in the three treatments did not differ in intake of the tannin food (P = 0.22), nor did intake of the terpene food differ for lambs in treatments 2 and 3. Total daily consumption of food with PSM was greater for lambs in treatment 1 (10.7 g/kg BW) than for lambs in treatments 2 (6.7 g/kg BW) or 3 (8.0 g/kg BW; P = 0.002; Fig. 2).

Intake of terpene- and tannin-containing foods by three groups of lambs during preference tests. During conditioning, lambs in treatment 1 (ad libitum terpenes, restricted tannins) were fed the terpene-containing food ad libitum and only 100 g of tannin-containing food. Lambs in treatment 2 (ad libitum tannins, restricted terpenes) were offered the tannin-containing food ad libitum and only 100 g of the terpene-containing food. Lambs in treatment 3 were offered both the tannin- and the terpene-containing foods ad libitum
Discussion
We determined whether the availability of two foods with different PSM affected total food intake and the food preferences of sheep. We hypothesized that the relative amount of tannins and terpenes ingested would influence the amount of food consumed as well as preference for each food. We predicted that lambs would decrease their preference for the PSM available ad libitum and that the PSM available in restricted amounts would be more preferred than the PSM available ad libitum, as the former were not eaten in amounts high enough to promote satiety and ensuing food aversions (Provenza 1996).
We found that lambs in all treatments preferred the tannin- to the terpene-containing food, but the preference was less pronounced for treatment 1 (restricted tannin, ad libitum terpenes) than for treatment 2 (ad libitum tannins, restricted terpenes) than for treatment 3 (ad libitum tannins and terpenes). The combination of restricted tannins/ad libitum terpenes enhanced overall food intake to a greater extent than ad libitum tannins/restricted terpenes or ad libitum tannins and terpenes. Our results suggest specific flavor-feedback characteristics of the foods, as well as the amount and possibly the order in which they were ingested, influenced intake by lambs of foods that contained tannins and terpenes.
During conditioning, the tannin-containing food induced satiety at a lower level of intake than did the terpene-containing food (Fig. 1). As tannin intake increased relative to terpene intake, total food consumption decreased indicating an antagonistic relationship between the tannin- and terpene-containing foods. While high intake of tannins (or terpenes) may cause rumen stasis and a general state of anorexia, this was not the case in this study, as lambs in all treatments ate alfalfa pellets readily after conditioning, nor was this the case in previous studies when lambs fed tannin- and terpene-containing foods ate comparable amounts to lambs fed the same nutritious basal foods without these PSM (Villalba et al. 2002a; Shaw et al. 2006a). Rather, Villalba et al. (2004) found that total intake of PSM was higher when tannin- and terpene-containing foods were fed in combination rather than alone. In that study (Villalba et al. 2004), intake of food with terpenes was similar whether terpenes were fed alone or fed with tannins, but the amount of tannin-containing food offered was not restricted; thus, higher tannin intake likely prevented further consumption of terpenes.
In the current study, intake of the terpene-containing food was three times higher when lambs were fed limited tannin and ad libitum terpene (treatment 1) than when they were fed ad libitum tannin and terpene (treatment 3). Conversely, when terpene was limited and tannin was fed ad libitum (treatment 2), tannin consumption was not enhanced relative to ad libitum tannin and terpene (treatment 3). Thus, by limiting the availability of the tannin-containing food, tannins were no longer the intake-limiting agent, and the high intake of the terpene-containing food likely was due to a neutralizing effect of a small amount of tannins on the negative effects of terpenes.
Lambs preferred tannins to terpenes, so they likely consumed tannins before terpenes in a meal. If so, tannins in the rumen may have ameliorated the negative effects of terpenes. This hypothesis, while requiring further testing, is consistent with findings of Mote et al. (2007), where tannins and terpenes were fed in different sequences. In that study, the highest PSM consumption occurred with the meal sequence tannin-containing food → terpene-containing food → nutritious food. Tannins likely bind to small amounts of terpenes reducing their effective concentration or increasing their rate of elimination, allowing sheep to eat more terpene-containing foods. Animals in treatment 3, fed ad libitum tannin, likely ingested more tannin than necessary to enhance terpene intake, explaining their lower total intake of PSM-containing food compared with lambs in treatment 1 fed with restricted tannin and ad libitum terpene.
Tannins are large molecules (molecular weights from 2,000 to 4,000) that remain in the rumen for many hours (Kumar and Singh 1984; Min and Hart 2003), whereas terpenes are small fat-soluble molecules (Foley and McArthur 1994) absorbed within minutes through the rumen epithelium (Dizba et al. 2006). Tannins are highly reactive molecules that once in the gut are likely to interact with terpenes. Tannins form complexes, mainly through hydrophobic/hydrogen interactions, with a wide variety of compounds including polysaccharides, nucleic acids, alkaloids, and minerals (Okuda et al. 1982; Min and Hart 2003). They also bind to and denature proteins (Jones and Mangan 1977; Burrows and Tyrl 2001). In so doing, they can reduce the production of volatile fatty acids (energy) and the supply of nitrogen (protein) adversely affecting the nutrition of herbivores (Foley et al. 1999).
At appropriate doses, however, tannins can improve the protein status of herbivores, thereby enhancing their ability to consume foods with PSM (Foley and McArthur 1994; Foley et al. 1999; Provenza et al. 2003b). Sheep and goats provided foods high in protein or given a choice between foods high in protein or energy, consume more tannins and terpenes than those fed only with high-energy foods (Villalba et al. 2002a, b). Moderate levels of tannins protect proteins from degrading in the rumen (Barry et al. 2001), which can improve the supply of high-quality amino acids to the abomasum and small intestines (Min and Hart 2003). In our study, tannin- and terpene-containing foods had soybean meal, a rich source of protein extensively degraded in the rumen (Nocek and Russell 1988). Rumen degradation of protein results in ammonia formation with subsequent loss of nitrogen in the urine (Satter et al. 1999). By binding to the protein in soybean meal, tannins may have protected proteins from degrading in the rumen and improved the supply of high-quality amino acids to the abomasum and small intestines, thus enhancing protein status and increasing terpene consumption by lambs in treatment 1.
Tannins were preferred to terpenes by all groups, but the differences were less pronounced for treatment 1 (59%) than for treatment 2 (70%) than for treatment 3 (79%). Flavor-feedback interactions likely initiated and then maintained the stronger preferences of lambs in treatments 2 and 3 for tannins over terpenes. The flavor of terpenes likely was sufficiently strong to cause lambs to prefer tannins over terpenes in treatments 2 and 3 where lambs were offered tannins ad libitum. Conversely, lambs in treatment 1 that were fed limited tannin and ad libitum terpene experienced greater exposure to the salient flavor of the terpenes in addition to the postingestive benefit from tannin–terpene interactions. These experiences collectively caused their higher preference for terpenes. This interpretation is consistent with two observations: (1) Several weeks are required for sheep to adapt to the flavor/postingestive consequences of eating foods high in terpenes (Banner et al. 2000) and (2) intake of foods with terpenes increases when alternative foods are restricted (Villalba et al. 2004; Shaw et al. 2006a, b).
Conversely, preference for tannins and avoidance of terpenes during conditioning reduced the ability of lambs in treatments 2 and 3 to maximize food intake. Indeed, their preference for tannins over terpenes caused diarrhea in over half of the lambs in those treatments, whereas no signs of diarrhea were observed in lambs under treatment 1. Fiber digestion in the rumen of sheep is depressed when condensed tannin concentrations reach 4% of DM intake (Roberts and Olson 1999), and tannins (Foley et al. 1999) and terpenes (Nagy and Tengerdy 1968) likely affect rumen bacteria selectively, influencing the ability of herbivores to consume the alternative PSM.
Our present and previous findings (Mote et al. 2007), and those of others (Wiggins et al. 2006a, b, c), suggest that the patchiness of plants with different secondary compounds is important in the foraging behavior of herbivores. Landscapes are mosaics of plant species with varied densities, diversities, and chemistries. Herbivores must move from patch to patch to maintain a balance of macro- and micronutrients. Meanwhile, they must cope with potentially toxic PSM. Preferences for various PSM can change dramatically when PSM are eaten in the right proportions and at the right times. Understanding both the spatial and the temporal contexts that lead to synergy may enable managers of ecological systems to create and maintain mixtures of plant species in a more even manner, creating greater biochemical diversity and reducing the competitive advantage of less-palatable plants that contain high concentrations of toxins.
There is growing appreciation of the importance of interactions between ecosystem chemistry and herbivores (Provenza et al. 2003a). Functional response models predict that food intake is a monotonically increasing function of plant density that asymptotes at maximum intake (Holling 1959). Thus, as the density of a chemotype in a plant community increases, the relative consumption of that chemotype decreases and vice-versa. The proportional disappearance of a plant species through herbivory is inversely related to its density (Provenza et al. 2003a). When the biomass of a chemotype reaches a certain critical amount (escape threshold), that species is increasingly protected from deleterious herbivory, leading to relatively high and ever-increasing use of less-abundant species (Provenza et al. 2003a). Herbivores become quickly satiated on the abundant chemotype and depend upon other complementary chemotypes to maintain appropriate nutrition (Provenza et al. 2003a, b). The preference for rare species does not necessarily increase, yet they may still be driven to extinction because of their rarity (Provenza et al. 2003a). These processes affect the structure and function of ecosystems (CAST 2002; Villalba et al. 2004), which in turn affect the ability of herbivores to meet their nutritional requirements.
Our study suggests that landscapes high in terpene-containing plants and low in tannin-containing plants may be used differently from ones high in tannins and low in terpenes. Herbivory in a high-terpene/low-tannin system would exert extreme pressure on the tannin components, perhaps to the point of extinction. Assuming plants are equally tolerant of herbivory, at some threshold of plant abundance for tannins and terpenes, as tannins become increasingly rare, the likelihood of tannin-containing plants disappearing from the system increases. A possible synergistic combination could be created in this situation such that herbivores offered limited amounts of tannin could increase pressure on terpene-containing plants helping maintain system stability. The alternative situation (high tannins/low terpenes) would be much more difficult to manage, as terpenes would likely have little influence on the use of tannin-containing plants, and terpene-containing plants may be used to the point of extinction. Thus, extinction rates for terpene-containing plants may be lower than for tannin-containing plants. This hypothesis could be tested with shrubs high in tannins (e.g., bitterbrush, Purshia tridentata, and oakbrush Quercus spp.) and high in terpenes (e.g., sagebrush, Artemisia tridentata). These shrubs codominate much of the sagebrush steppe ecosystem, which occupies nearly 1.1 million km2 of the western USA and Canada (Meyer and Karasov 1991). Consistent with the proposed interaction between terpenes and tannins, a previous study has shown that sheep manifesting a high preference for sagebrush (terpenes) consume more bitterbrush (tannins) than sheep manifesting a low preferences for sagebrush (Seefeldt 2005).
Finally, our results suggest that tannin-containing plants, such as bitterbrush and oakbrush, may be valuable components of landscapes in the development of grazing systems in the sagebrush steppe ecosystem. By combining the proper proportions of PSM and nutrients, in the right sequence, mangers can potentially optimize the use of dominant plant species with PSM. In our study, a small amount of tannins increased the ability of lambs to consume terpenes, whereas a small amount of terpenes had no such beneficial affect on tannin consumption. Therefore, some dietary mixtures, amounts, and sequences are likely to be more favorable than others nutritionally and physiologically. Thus, we hypothesize that grazing systems that combine limited tannins and large meals of terpenes will more evenly use landscapes high in terpene-containing plants, reducing their competitive advantage and increasing the plant diversity of those landscapes.
References
Banner, R. E., Rogosic, J., Burritt, E. A., and Provenza, F. D. 2000. Supplemental barley and activated charcoal increase intake of sagebrush by lambs. J. Range Manag. 53:415–420.
Barry, T. N., Mcneill, D. M., and Mcnabb, W. C. 2001. Plant secondary compounds: their impact on nutritive value and upon animal production, pp. 445–452, in Proc. XIX Int. Grass. Conf., Sao Paulo, Brazil.
Burritt, E. A. and Provenza, F. D. 2000. Role of toxins in intake of varied diets by sheep. J. Chem. Ecol. 26:1991–2005.
Burrows, G. E. and Tyrl, R. J. 2001. Toxic Plants of North America. Iowa State Press, Ames, Iowa.
Cheeke, P. and Schull, L. R. 1985. Natural Toxicants in Feeds and Poisonous Plants. Avi, Westport, CT.
Clausen, T. P., Provenza, F. D., Burritt, E. A., Reichardt, P. B., and Bryant, J. P. 1990. Ecological implication of condensed tannin structure: a case study. J. Chem. Ecol. 16:2381–2392.
Council for Agriculture Science and Technology (CAST) 2002, in W. C. Krueger, M. A. Sanderson, J. B. Cropper, M. Miller-Goodman, C. E. Kelley, R. D. Pieper, P. L. Shaver, and M. J. Trlica (eds.). Environmental Impacts of Livestock on U.S. Grazing Lands. Number 22.
Dearing, M. D. and Cork, S. 1999. Role of detoxification of plant secondary compounds on diet breadth in a mammalian herbivore, Trichosurus vulpecula. J. Chem. Ecol. 25:1205–1219.
Dizba, L. E., Hall, J. O., and Provenza, F. D. 2006. Feeding behavior of the lambs in relation to kinetics of 1.8-ceneole dosed intravenously or into the rumen. J. Chem. Ecol. 32:2 391–408.
Foley, W. J. and Mcarthur, C. 1994. The effects and costs of allelochemicals for mammalian herbivores: an ecological perspective, pp. 370–391, in D. J. Chivers and P. Langer (eds.). The Digestive System in Mammals: Food, Form and Function. Cambridge Univ. Press, Cambridge, UK.
Foley, W. J., Iason, G. R., and Mcarthur, C. 1999. Role of plant secondary metabolites in the nutritional ecology of mammalian herbivores: how far have we come in 25 years?, pp. 130–209, in H. G. Jung and G. C. Fahey, Jr. (eds.). Nutritional Ecology of Herbivores. Proc. Vth Int. Symp. Nutr. Herb. Am. Soc. Anim. Sci., Illinois.
Freeland, W. J. and Janzen, D. H. 1974. Strategies of herbivory by mammals: the role of plant secondary compounds. Am. Nat. 108:269–286.
Holling, C. S. 1959. Some characteristics of simple types of predation and parasitism. Can. Entemol. 91:385–398.
Johnson, A. E., James, L. F., and Spillet, J. 1976. The abortifacient and toxic effects of big sagebrush (Artemisia tridentata) and juniper (Juniperus osteosperma) on domestic sheep. J. Range Manag. 29:278–280.
Jones, W. T. and Mangan, J. L. 1977. Complexes of condensed tannins of sainfoin (Onobrychis viciifolia Scop.) with fraction-1 leaf protein and with submaxillary mucoprotein, and their reversal by polyethylene–glycol and pH. J. Sci. Food Agric. 28:126–136.
Kimball, B. A., Nolte, D. L., Engenman, R. M., Johnston, J. J., and Stermitz. F. R. 1998. Chemically mediated foraging preference of black bears (Ursus americanus). J. Mammal. 79:448–456.
Kumar, R. and Singh, M. 1984. Tannins: their adverse role in ruminant nutrition. J. Agric. Food Chem. 32:447–453.
Littell, R. C., Milliken G. A., Stroup W. W., and Wolfinger R. D. 1996. SAS System for Mixed Models. SAS Institute, Cary, NC.
Marsh, K. J., Wallis, I. R., Mclean, S., Sorensen, J. S., and Foley, W. J. 2006. Conflicting demands on detoxification pathways influence how common brushtail possums choose their diets. Ecology 87:2103–2112.
Mehansho, H., Butler, L. G., Rogler, J. C., and Carlson, D. M. 1987. Dietary tannins and salivary proline-rich proteins: interactions, induction, and defense mechanisms. Annu. Rev. Nutr. 7:423–440.
Meyer, M. W. and Karasov, W. H. 1991. Chemical aspects of herbivory in arid and semiarid habitats, p. 177, in R. T. Palo and C. T. Robbins (eds.). Plant Defenses Against Mammalian Herbivory. CRC, Boca Raton, FL.
Min, B. R. and Hart, S. P. 2003. Tannins for suppression of internal parasites. J. Anim. Sci. 81(E. Suppl. 2):E102–E109.
Mote, T. E., Villalba, J. J., and Provenza, F. D. 2007. Influence of foraging sequences on the ability of lambs to consume toxins when fed tannins, terpenes and a high-quality food. Appl. Anim. Behav. Sci. In press.
Nagy, J. G. and Tengerdy, R. P. 1968. Antibacterial action of essential oils of Artemisia as an ecological factor. II. Antibacterial action of the volatile oils of Artemisia tridentata (big sagebrush) on bacteria from the rumen of mule deer. Appl. Microbiol. 16:441–444.
Nocek, J. E. and Russell, J. B. 1988. Protein and energy as an integrated system. Relationship of ruminal protein and carbohydrate availability to microbial synthesis and milk production. J. Dairy Sci. 71:2070–2107.
NRC. 1985. Nutrient Requirements of Sheep (6th Ed.). National Academy, Washington, DC.
Okuda, T., Mori, K., and Shiota, M. 1982. Effects of interaction of tannins and coexisting substances. III. Formation and solubilization of precipitates with alkaloids. Yakugaku Zasshi 102:854–858.
Provenza, F. D. 1996. Aquired aversions as the basis for varied diets of ruminants foraging on rangelands. J. Anim. Sci. 74:2010–2020.
Provenza, F. D. and Villalba, J. J. 2006. Foraging in domestic herbivores: Linking the internal and external milieu, pp. 210–240, in V. L. Bels (ed.). Feeding in Domestic Vertebrates: from Structure to Function. CABI, Oxfordshire, UK.
Provenza, F. D., Villalba, J. J., and Bryant, J. P. 2003a. Foraging by herbivores: linking the biochemical diversity of plants to herbivore culture and landscape diversity, pp. 387–421, in J. A. Bissonette and I. Storch (eds.). Landscape Ecology and Resource Management: Linking Theory with Practice. Island Press, New York.
Provenza, F. D., Villalba, J. J., Dziba, L. E. Atwood, S. B. and Banner, R. E. 2003. Linking herbivore experience, varied diets, and plant biochemical diversity. Small Rum. Res. 49:257–274.
Roberts, J. L. and Olson B. E. 1999. Effect of Euphorbia esula on sheep rumen microbial activity and mass in vitro. J. Chem. Ecol. 25:297–314.
Satter, L. D., Jung, H. G., Van Vuuren, A. M., and Engels, F. M. 1999. Challenges in the nutrition of high-producing ruminants, pp. 609–646, in Jung, H.G. and G.C. Fahey, Jr. (eds.), Nutritional Ecology of Herbivores. Proc. 5th Int. Symp. Nutr. Herb. Am. Soc. Anim. Sci., Illinois.
Seefeldt, S. S. 2005. Consequences of selecting Rambouillet ewes for mountain big sagebrush (Artemisia tridentata ssp. vaseyana) dietary preference. Rangeland Ecol. Manage. 58:380–384.
Shaw, R. A., Villalba, J. J., and Provenza, F. D. 2006a. Resource availability and quality influence patterns of diet mixing with foods containing toxins by sheep. J. Chem. Ecol. 32:1267–1278.
Shaw, R. A.,Villalba, J. J., and Provenza, F. D. 2006b. Influence of stock density and rate and temporal patterns of forage allocation on the diet mixing behavior of sheep grazing sagebrush steppe. Appl. Anim. Behav. Sci. 100:207–218.
Sipes, I. G. and Gandolfi, A. J. 1986. Biotransformation of toxicants, pp. 609–646, in C. D. Klassen, M. O. Amdur, and J. Doull (eds.). Casarett and Doull’s Toxicology, The Basic Science of Poisons. Macmillian, New York, NY.
Villalba, J. J. and Provenza, F. D. 1999. Nutrient-specific preferences by lambs conditioned with intraruminal infusions of starch, cascein, and water. J. Anim. Sci. 79:2066–2074.
Villalba, J. J., Provenza, F. D., and Banner, R. E. 2002a. Influence of macronutrients and polyethylene glycol on intake of a quebracho tannin diet by sheep and goats. J. Anim Sci. 80:3154–3164.
Villalba, J. J., Provenza, F. D., and Banner, R. E. 2002b. Influence of macronutrients and activated charcoal on utilization of sagebrush by sheep and goats. J. Anim Sci. 80:2099–2109.
Villalba, J. J., Provenza, F. D., and Han, G. 2004. Experience influences diet mixing by herbivores: implications for plant biochemical diversity. Oikos 107:100–109.
Villalba, J. J., Provenza, F. D., and Olson, K. C. 2006. Terpenes and carbohydrate source influence rumen fermentation, digestibility, intake, and preference in sheep. J. Anim. Sci. 84:2463–2473.
Wiggins, N. L., Mcarthur, C., Davies, N. W., and Mclean, S. 2006a. Diet switching in a generalist mammalian folivore: fundamental to maximizing intake. Oecologia 147:650–657.
Wiggins, N. L., Mcarthur, C., Davies, N. W., and Mclean, S. 2006b. Behavioral responses of a generalist mammalian folivore to the physiological constraints of a chemically defended diet. J. Chem. Ecol. 32:1133–1147.
Wiggins, N. L., Mcarthur, C., Davies, N. W., and Mclean, S. 2006c. Spatial scale of the patchiness of plant poisons: a critical influence on foraging efficiency. Ecology 87:2236–2243.
Acknowledgements
This research was supported by grants from the Utah Agricultural Experiment Station and the Initiative for the Future of Agriculture and Food Systems, USDA (Agreement no. 2001-52103-11215). This paper is published with the approval of the Director, Utah Agricultural Experiment Station, and Utah State University, as paper number 7815.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Mote, T.E., Villalba, J.J. & Provenza, F.D. Relative Availability of Tannin- and Terpene-containing Foods Affects Food Intake and Preference by Lambs. J Chem Ecol 33, 1197–1206 (2007). https://doi.org/10.1007/s10886-007-9305-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10886-007-9305-2