
Abstract
CD40 ligand (CD40L) deficiency or X-linked hyper-IgM syndrome (X-HIGM) is a well-described primary immunodeficiency in which Pneumocystis jiroveci pneumonia is a common clinical feature. We have identified an unusual high incidence of fungal infections and other not yet described infections in a cohort of 11 X-HIGM patients from nine unrelated Brazilian families. Among these, we describe the first case of paracoccidioidomycosis (PCM) in X-HIGM. The molecular genetic analysis of CD40L was performed by gene sequencing and evaluation of CD40L protein expression. Nine of these 11 patients (82%) had fungal infections. These included fungal species common to CD40L deficiency (P. jiroveci and Candida albicans) as well as Paracoccidioides brasiliensis. One patient presented with PCM at age 11 years and is now doing well at 18 years of age. Additionally, one patient presented with a simultaneous infection with Klebsiella and Acinetobacter, and one with condyloma caused by human papilloma virus. Molecular analysis revealed four previously described CD40L mutations, two novel missense mutations (c.433 T > G and c.476 G > C) resulting in the absence of CD40L protein expression by activated CD4+ cells and one novel insertion (c.484_485insAA) within the TNFH domain leading to a frame shift and premature stop codon. These observations demonstrated that the susceptibility to fungal infections in X-HIGM extends beyond those typically associated with X-HIGM (P. jiroveci and C. albicans) and that these patients need to be monitored for those pathogens.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Congenital susceptibility to fungal infections is most frequently observed in primary immunodeficiency disorders associated with deficiency or dysfunction of T cells or phagocytes (APECED, STAT1, IL-17R, and STAT3 deficiencies) [1–4]. Mutations in CD40 ligand (CD40L; CD154, MIM #300386), a surface protein expressed by activated CD4+ T cell deficiency, is a well-defined T cell defect that causes the X-linked hyper-IgM syndrome (X-HIGM) [5–9]. In addition to extracellular and intracellular bacterial infections, patients with X-HIGM are susceptible to fungal infections, particularly Pneumocystis jiroveci pneumonia, and infections with Candida albicans, Cryptococcus and Histoplasma [10–12]. Interestingly, the majority of these infections occur within the first 5 years of life and are much less common thereafter [11–13].
In humans, the effective immune response against fungi is dependent on both innate and adaptive responses. Appropriate activation of phagocytic cells and particularly the production of reactive oxygen intermediate by nicotinamide adenine dinucleotide phosphate oxidase are important for the control of fungal infections [14]. The activation of this system is coordinated by the interleukin (IL)-12/interferon-γ (IFN-γ) axis, which is triggered by CD40/CD40L interaction [15] and mediated by Th1 cells. On the other hand, the humoral immune response orchestrated by antibodies plays an essential role in the defense against extracellular bacteria as illustrated by the increased susceptibility to these infections by patients with predominantly antibody deficiencies [16]. In this context, the CD40–CD40L interaction plays an important role by facilitating the expression of activation-induced cytidine deaminase and other proteins involved in the class switch recombination and somatic hypermutation of immunoglobulin (Ig) variable region genes [17].
Here we report the clinical features and molecular defects of a cohort of X-HIGM patients identified in Brazil, with a high incidence of fungal infections (82%) compared to the incidence described in Europe and North America (34% to 57%) [11, 13]. The spectrum of fungal infections in our patients includes species that have never been described in X-HIGM such as paracoccidioidomycosis (PCM). In addition, we observed a patient who developed sepsis by being simultaneously infected with Klebsiella pneumonia and Acinetobacter sp. and a case of condyloma caused by human papilloma virus (HPV) and extended the number of known CD40L mutations by four.
Methods
Subjects
Between 2006 and 2011, we identified 11 patients from nine unrelated families with CD40L deficiency in our clinic. The criteria for diagnosing CD40L deficiency complied with the International Union of Immunological Societies Expert Committee on Primary Immunodeficiencies [16], based on characteristic clinical criteria and confirmed by the lack of CD40L expression by activated T cells and the presence of mutation in the CD40L gene. Informed consent was obtained from all patients or their parents, and blood was collected under institutional guidelines. The study was approved by the Ethics Committee at the Institute of Biomedical Sciences, University of São Paulo, according to the Helsinki Convention and the Brazilian Department of Health.
Cell Culture and Activation
Peripheral blood mononuclear cells were isolated from heparinized blood after Ficoll-Hypaque sedimentation and cultured (1 × 106 cells/well) for 5 h at 37°C in 5% CO2 in flat-bottomed microtiter plates in a final volume of 200 μl of RPMI 1640 (Gibco Laboratories), supplemented with 10% fetal calf serum, 1% glutamine, and antibiotics, with or without phorbol myristate acetate (15 ng/ml, Sigma Laboratories, St. Louis, MO, USA) and calcium ionophore (ionomycin, 300 ng/ml, Sigma Laboratories).
Lymphocyte Counts and CD40L Expression
T (CD3+CD4+, CD3+CD8+) and B (CD19+CD40+) cell counts were determined by flow cytometry analysis using specific monoclonal antibodies. CD40L expression was assessed on the surface of resting and activated CD3+CD4+ T cells using the monoclonal antibody TRAP1 (Mouse IgG1) as previously described [18]. Activation of cells was confirmed by staining CD3+CD4+ T cells with specific antibodies to CD69 (Becton Dickinson, BD) (data not shown). Prior to flow cytometry, samples were washed and fixed in 1% paraformaldehyde. The results obtained were analyzed using FlowJo software (Treestar Inc., Ashlan, OR, USA).
Sequence Analysis of CD40L Gene
Genomic DNA (gDNA) was isolated from EDTA blood using the Wizard® Genomic DNA Purification kit (Promega). All five exons of the CD40L gene, including the exon–intron boundaries, were individually amplified by polymerase chain reaction (PCR), as previously described [18]. Primers are available upon request. PCR products were purified using the GFXTM PCR DNA and Gel Band Purification Kit (GE Healthcare), according to the manufacturer’s instructions. PCR products were sequenced on a MegaBACE 1000 sequencer using the DYEnamic ET Dye Terminator kit (Thermo Sequenase™ II DNA Polymerase). The observed sequences were compared with the CD40L mutation registry (http://bioinf.uta.fi/CD40Lbase/), and structural analysis of missense mutations was performed based on the crystal structure of the protein (PDB 1ALY). CD40L complementary DNA (cDNA) was also sequenced as previously described [18]. To provide evidence that the two novel missense mutations identified are not single-nucleotide polymorphisms (SNPs), we evaluated both mutations using MutationTaster, a bioinformatic method to evaluate disease-causing potential of sequence alterations [19] and sequenced gDNA from 100 healthy Brazilian controls.
Results
Patient Baseline Characteristics
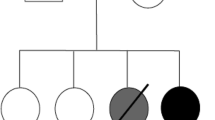
We enrolled 11 male patients ranging in age from 1 to 23 years in whom a diagnosis of X-HIGM syndrome was suspected. All patients came from nonconsanguineous Brazilian families (Fig. 1). Median age at time of disease onset was 6 months (ranging from 1 to 7 months). The median age at the time of X-HIGM diagnosis was 2 years (ranging from 5 months to 6 years).

Patients’ pedigrees. Black squares represent males with recurrent, severe infections. Black squares with slashes indicate deceased individuals. All female carriers, represented by circles with dots, were clinically and immunologically healthy
High Incidence of Fungal Infections and the Occurrence of Paracoccidioidomycosis
Nine of 11 (82%) patients in this cohort presented with fungal infections, many of which were being unusual or severe (Table I). Six patients had P. jiroveci pneumonia, an infection that typically occurs in young (under the age of 5 years) CD40L-deficient patients [11]; three patients had infections caused by C. albicans which were either of an unusual type or particularly severe: P5 was diagnosed with severe candida esophagitis, P6 with oral and perianal candidiasis, and P9 with persistent cutaneous candidiasis on the scalp (Fig. 2a). It is important to note that at the time of the candidiasis, P5 and P6 but not P9 were treated prophylactically with trimethoprim–sulfamethoxazole to prevent P. jiroveci pneumonia, but no other antibiotics were used. None presented with neutropenia at the time the Candida diagnosis was established.

Paracoccidioidomycosis and cutaneous candidiasis in CD40L-deficient patients. a Picture of persistent candidiasis on the scalp of P9. b Histopathologic characteristics of a lymph node biopsy of patient P10. Hematoxylin–eosin-stained specimen showing the tuberculoid granulomatous inflammation with Paracoccidioides brasiliensis within a multinucleated cell
At the age of 6 months, patient P10, born and living in Sao Paulo city, was diagnosed with X-HIGM while presenting with P. jiroveci pneumonia, recurrent otitis media, and sinusitis. At the age of 11 years while on intravenous immunoglobulin (IVIG) treatment, he presented with mild prolonged fever and cough. Chest CT scans showed mediastinal lymphadenopathy and a lymph node biopsy confirmed tuberculoid granulomatous inflammation as a consequence of Paracoccidioides brasiliensis within multinuclear cells (Fig. 2b). In addition, bone marrow biopsy revealed hypoplasia and tuberculoid granuloma, a clinical feature compatible with the acute form of PCM. It is important to note that P10 did not present with neutropenia 6 months before or during PCM. He was treated with an 8-month course of itraconazol and recovered.
Extracellular Bacterial Infections
One patient (P9) presented with local pyoderma and oropharyngitis beginning at 3 months of age followed by recurrent otitis and mastoiditis. One of the pathogens causing otitis was Pseudomonas aeruginosa. At the age of 3 years, he developed severe mastoiditis followed by sepsis caused by K. pneumonia and Acinetobacter sp., leading to the diagnosis of X-HIGM syndrome. Under treatment with cefepime and vancomycin, he fully recovered and IVIG therapy was started. P9 died at 8 years of age due to complications resulting from progressive multifocal leukoencephalopathy.
Other Isolated Etiologic Agents
Other etiologic agents causing infections were isolated in P3 and P6 while on adequate IVIG prophylaxis. The protozoan Cryptosporidium parvum, a relatively common pathogen observed in CD40L-deficient patients [11], caused chronic diarrhea in P3 at 4 years of age. P6 presented with a herpes simplex virus infection, which has been previously reported in X-HIGM patients [13]. In addition, he had perianal condyloma caused by HPV.
Laboratory and Immunologic Findings
Patient lymphocyte counts (CD3+CD4+, CD3+CD8+, CD19+CD40+) were similar to healthy controls (Table II). Neutropenia was identified in six patients (P2, P3, P4, P5, P6, and P10). In P3, P4, and P11, neutropenia was associated with recurrent oral or esophageal ulcers but not with other clinical features. Neutropenia was effectively treated with recombinant human granulocyte colony-stimulating factor in all patients except P4. Despite presenting persistent neutropenia, P4 did not have fungal infections.
All patients had low serum levels of IgG and IgA (Table II). Serum IgM was elevated in patients P1, P2, P7, P9, P10, and P11, but was normal in patients P3, P4, and P6 and reduced in patients P5 and P8.
Molecular Genetic Analysis of CD40L
Expression of the CD40L protein on the surface of activated CD3+CD4+ T cells was impaired in all patients except for P3, who had an expression level similar to the healthy control (Fig. 4). Seyama et al. [18] used a CD40–Ig fusion protein to demonstrate that the c.496 C > A mutation identified in P3 affects binding of CD40 to CD40L on activated T cells.
Sequence analysis of the CD40L gene revealed three novel mutations. These are two missense mutations, one in exon 4 (c.433 T > G, p.V126G, P1), the other in exon 5 (c.476 G > C, p.W140C, P2), and an insertion of 2 base pairs (c.551_552insAA, p.R165X190) within the tumor necrosis factor homologous (TNFH) domain found in one patient (P11) (Table III, Fig. 3b). MutationTaster analysis indicated no SNPs for these altered regions and neither of these two mutations was found in 100 healthy Brazilian control subjects. Furthermore, comparative analysis of the CD40L protein from different species showed that the two novel base pair substitutions affect conserved amino acids within the CD40L TNFH domain (Fig. 3a). These amino acid residues are essential for the CD40L structure, thereby abolishing expression of the CD40L protein on the surface of activated CD3+CD4+ T cells (Fig. 3a).

Novel mutations in CD40L-deficient patients. a The missense mutations affect conserved amino acids in the CD40L TNF homology domain (TNFH) of different species and are located at the binding sites with CD40 receptor. b The insertion of two adenines (p.R165X190) found in the CD40L gene of P11 results in a frame shift mutation creating a premature stop codon
The insertion found in P11 leads to a frame shift mutation with the creation of a premature stop codon resulting in an unstable truncated protein (Fig. 3b). The other four unique mutations identified in the remaining eight patients have been described previously (Table III) and their consequences in CD40L expression are demonstrated in Fig. 4.

Defective CD40L protein expression. Flow cytometry histograms showing CD40L expression on the surface of activated CD3+CD4+ T cells from patients (P1–P11) demonstrating a lack of protein expression in all but one patient (P3) who expresses a nonfunctional protein as previously demonstrated [18]. Histogram C shows data from a healthy control
Discussion
Common clinical features of CD40L-deficient patients include infections caused by extracellular and intracellular bacterial and fungal pathogens. Pneumonia caused by the commensal fungus P. jiroveci is a well-recognized complication in CD40L deficiency with incidence rates ranging from 32% to 42% [11, 13]. However, reports of other fungal infections are rare: Candida in two patients [12, 20], Cryptococcus in one patient [21], and Histoplasma in one patient [10]. While candidiasis and P. jiroveci infections usually occur during the first 5 years of life, the patients developing cryptococcosis and histoplasmosis and the Brazilian patient developing P. brasiliensis where 12, 19, and 11 years, respectively.
In this study, we identified a high rate of fungal infections (82%) in a cohort of 11 CD40L-deficient patients from Brazil, none of which had neutropenia at the time of presentation. These infections were caused by a broad range of fungal pathogens including P. jirovecii, C. albicans, and, not previously reported, one patient with P. brasiliensis infection. These observations suggest a possible underlying mechanism responsible for the occurrence of fungal infections which remains to be investigated. The reason for the high incidence of fungal infections and the occurrence of PCM described in this cohort of X-HIGM patients agrees with distinct epidemiological features and unique etiologic agents associated with the geographic and socioeconomic conditions in Latin America [22]. For example, in Brazil, the incidence of Candida infections is almost ten times higher than that reported in North America and Europe [22]. Accordingly, we identified three X-HIGM patients with severe Candida infections. P. brasiliensis is a fungus that causes the most important endemic systemic mycosis in South America [23]. Interestingly, both patients with primary immunodeficiency presenting with PCM are Brazilian. One is a patient with a mutation in the beta 1 subunit of the IL-12/IL-23 receptor [24] and the second is the patient (P10) described here. Both patients live in São Paulo, a huge urban area where the incidence of PCM is much lower than in rural regions. The actual incidence and prevalence of PCM is poorly known because reporting this disease is not mandatory in most South American countries, and its distribution is highly variable even in endemic areas [25]. However, some isolated studies reported the PCM average incidence from one to four new cases per 100,000 habitants per year in Brazil [26], whereas in other South American countries, it has been estimated an average incidence of 0.5 cases per 100,000 habitants per year with lower rates being described in Colombia [27, 28].
PCM is an infection acquired by inhalation of airborne propagules produced by fungal mycelium, which then transform into pathogenic yeast [29]. After P. brasiliensis invades the host via the respiratory tract, it persists in macrophages causing granuloma formation and disseminates through the reticuloendothelial system [30]. This pathogenesis of persistent P. brasiliensis inside macrophages and granuloma formation in lymph nodes and bone marrow was observed in our patient. Two different clinical forms of PCM can be distinguished: an acute, juvenile form and a chronic, adult form [31, 32]. The clinical picture of the acute form reveals a severe disease with reticuloendothelial system organ hypertrophy, bone marrow dysfunction, and septic episodes. The lungs are seldom the primary focus of infection but fungal pathogens can be proven in pulmonary secretions assuming some lung involvement [33]. The disease develops within a few weeks or months and is more frequently reported in children and young adults below 25 years of age [31, 34]. On the other hand, the chronic form results from infection usually acquired during the first two decades of the life staying clinically silent and causing disease in adults, mostly men of 30 to 50 years of age, by reactivation of a latent endogenous focus [32]. The disease progresses slowly with symptoms of cough, expectoration, weight loss, and fever. A characteristic of the chronic form is pulmonary manifestation; however, it can also occur as a multifocal form with fungal spread to more than one organ or tissue such as the oral mucosa, skin, lymph nodes [35–37], central nervous system, and gastrointestinal and ocular manifestations [38–40].
To eradicate PCM, an efficient Th1 immune response is required, characterized by the production of IFN-γ and subsequent activation of phagocytic cells [41]. By enhancing cytochrome b expression by phagocytes, IFN-γ is believed to potentiate the oxidative respiratory burst in macrophages and polymorphonuclear leukocytes, a process responsible for many of the microbicidal, tumoricidal, and inflammatory activities of these cells [27, 28]. In accordance, P. brasiliensis-pulsed mDCs from CD40L-deficient patients induce a Th2-skewed T cell response characterized by lower IFN-γ and higher IL-4 and IL-5 production compared to healthy subjects [42], an immunological milieu associated with increased susceptibility to fungal infections in humans [43].
The inability to eliminate extracellular bacterial and viral pathogens is also a common clinical feature of patients with X-HIGM and other well-defined humoral immunodeficiencies [11, 44]. However, only one CD40L-deficient patient has been reported with infections caused by P. aeruginosa [45], and this is the first report of sepsis caused by K. pneumonia and Acinetobacter sp. and of condyloma associated with HPV infections in patients with CD40L deficiency. K. pneumonia and Acinetobacter sp. are two gram-negative bacteria present in the gastrointestinal flora of immunocompetent individuals, causing worldwide nosocomial infections in newborns and in patients with AIDS [46, 47]. Among primary immunodeficient patients, Klebsiella sp. infections were only reported in chronic granulomatous disease [48] and X-linked agammaglobulinemia [49]. HPV infections were described in patients with hypogammaglobulinemia, highlighting the role of the antibody in controlling some viral pathogens. The success of vaccines against HPV in protecting women against HPV infection and cervical cancer [50] underlines the importance of adaptive immunity in the protection against this virus. HPV infections were also observed in patients with autosomal-dominant mutations in the chemokine receptor gene CXCR4 [51], but the underlying mechanism involved in this susceptibility still needs to be clarified.
In conclusion, our data highlight the key role of the CD40/CD40L signaling pathway in mediating immunity against fungal and bacterial infections. It demonstrates that defects in this pathway cause susceptibility to a much broader array of pathogens than those traditionally associated with X-HIGM (i.e., P. jirovecii). Therefore, we suggest that patients with CD40L deficiency especially if living in tropical areas should be monitored for these pathogens.
References
Liu L, Okada S, Kong XF, Kreins AY, Cypowyj S, Abhyankar A, Toubiana J, Itan Y, Audry M, Nitschke P, Masson C, Toth B, Flatot J, Migaud M, Chrabieh M, Kochetkov T, Bolze A, Borghesi A, Toulon A, Hiller J, Eyerich S, Eyerich K, Gulacsy V, Chernyshova L, Chernyshov V, Bondarenko A, Maria Cortes Grimaldo R, Blancas-Galicia L, Madrigal Beas IM, Roesler J, Magdorf K, Engelhard D, Thumerelle C, Burgel PR, Hoernes M, Drexel B, Seger R, Kusuma T, Jansson AF, Sawalle-Belohradsky J, Belohradsky B, Jouanguy E, Bustamante J, Bue M, Karin N, Wildbaum G, Bodemer C, Lortholary O, Fischer A, Blanche S, Al-Muhsen S, Reichenbach J, Kobayashi M, Rosales FE, Lozano CT, Kilic SS, Oleastro M, Etzioni A, Traidl-Hoffmann C, Renner ED, Abel L, Picard C, Marodi L, Boisson-Dupuis S, Puel A, Casanova JL. Gain-of-function human STAT1 mutations impair IL-17 immunity and underlie chronic mucocutaneous candidiasis. J Exp Med. 2011;208:1635–48.
Puel A, Cypowyj S, Bustamante J, Wright JF, Liu L, Lim HK, Migaud M, Israel L, Chrabieh M, Audry M, Gumbleton M, Toulon A, Bodemer C, El-Baghdadi J, Whitters M, Paradis T, Brooks J, Collins M, Wolfman NM, Al-Muhsen S, Galicchio M, Abel L, Picard C, Casanova JL. Chronic mucocutaneous candidiasis in humans with inborn errors of interleukin-17 immunity. Science. 2011;332:65–8.
Ryan KR, Hong M, Arkwright PD, Gennery AR, Costigan C, Dominguez M, Denning D, McConnell V, Cant AJ, Abinun M, Spickett GP, Lilic D. Impaired dendritic cell maturation and cytokine production in patients with chronic mucocutaneous candidiasis with or without APECED. Clin Exp Immunol. 2008;154:406–14.
Schimke K, Chubb SA, Davis WA, Davis TM. Helicobacter pylori cytotoxin-associated gene-A antibodies do not predict complications or death in type 2 diabetes: the Fremantle Diabetes Study. Atherosclerosis. 2010;212:321–6.
Fuleihan R, Ramesh N, Geha RS. Role of CD40–CD40-ligand interaction in Ig-isotype switching. Curr Opin Immunol. 1993;5:963–7.
Allen RC, Armitage RJ, Conley ME, Rosenblatt H, Jenkins NA, Copeland NG, Bedell MA, Edelhoff S, Disteche CM, Simoneaux DK, et al. CD40 ligand gene defects responsible for X-linked hyper-IgM syndrome. Science. 1993;259:990–3.
Aruffo A, Farrington M, Hollenbaugh D, Li X, Milatovich A, Nonoyama S, Bajorath J, Grosmaire LS, Stenkamp R, Neubauer M, et al. The CD40 ligand, gp39, is defective in activated T cells from patients with X-linked hyper-IgM syndrome. Cell. 1993;72:291–300.
DiSanto JP, Bonnefoy JY, Gauchat JF, Fischer A, de Saint Basile G. CD40 ligand mutations in X-linked immunodeficiency with hyper-IgM. Nature. 1993;361:541–3.
Fuleihan R, Ramesh N, Loh R, Jabara H, Rosen RS, Chatila T, Fu SM, Stamenkovic I, Geha RS. Defective expression of the CD40 ligand in X chromosome-linked immunoglobulin deficiency with normal or elevated IgM. Proc Natl Acad Sci U S A. 1993;90:2170–3.
Hostoffer RW, Berger M, Clark HT, Schreiber JR. Disseminated Histoplasma capsulatum in a patient with hyper IgM immunodeficiency. Pediatrics. 1994;94:234–6.
Levy J, Espanol-Boren T, Thomas C, Fischer A, Tovo P, Bordigoni P, Resnick I, Fasth A, Baer M, Gomez L, Sanders EA, Tabone MD, Plantaz D, Etzioni A, Monafo V, Abinun M, Hammarstrom L, Abrahamsen T, Jones A, Finn A, Klemola T, DeVries E, Sanal O, Peitsch MC, Notarangelo LD. Clinical spectrum of X-linked hyper-IgM syndrome. J Pediatr. 1997;131:47–54.
Nonoyama S, Shimadzu M, Toru H, Seyama K, Nunoi H, Neubauer M, Yata J, Och HD. Mutations of the CD40 ligand gene in 13 Japanese patients with X-linked hyper-IgM syndrome. Hum Genet. 1997;99:624–7.
Winkelstein JA, Marino MC, Ochs H, Fuleihan R, Scholl PR, Geha R, Stiehm ER, Conley ME. The X-linked hyper-IgM syndrome: clinical and immunologic features of 79 patients. Medicine (Baltimore). 2003;82:373–84.
Romani L. Immunity to fungal infections. Nat Rev Immunol. 2004;4:1–23.
Al-Muhsen S, Casanova JL. The genetic heterogeneity of mendelian susceptibility to mycobacterial diseases. J Allergy Clin Immunol. 2008;122:1043–51. quiz 1052–1043.
Notarangelo LD, Fischer A, Geha RS, Casanova JL, Chapel H, Conley ME, Cunningham-Rundles C, Etzioni A, Hammartrom L, Nonoyama S, Ochs HD, Puck J, Roifman C, Seger R, Wedgwood J. Primary immunodeficiencies: 2009 update. J Allergy Clin Immunol. 2009;124:1161–78.
Stavnezer J. Complex regulation and function of activation-induced cytidine deaminase. Trends Immunol. 2011;32:194–201.
Seyama K, Nonoyama S, Gangsaas I, Hollenbaugh D, Pabst HF, Aruffo A, Ochs HD. Mutations of the CD40 ligand gene and its effect on CD40 ligand expression in patients with X-linked hyper IgM syndrome. Blood. 1998;92:2421–34.
Schwarz JM, Rodelsperger C, Schuelke M, Seelow D. MutationTaster evaluates disease-causing potential of sequence alterations. Nat Methods. 2010;7:575–6.
Kraakman EM, Bontrop RE, Groenestein R, Jonker M, Haaijman JJ, t Hart BA. Characterization of the natural immune response of rhesus monkey CD4 + ve T cells to the bacterial antigen streptolysin O (SLO). J Med Primatol. 1995;24:306–12.
Jo EK, Kim HS, Lee MY, Iseki M, Lee JH, Song CH, Park JK, Hwang TJ, Kook H. X-linked hyper-IgM syndrome associated with Cryptosporidium parvum and Cryptococcus neoformans infections: the first case with molecular diagnosis in Korea. J Korean Med Sci. 2002;17:116–20.
Nucci M, Queiroz-Telles F, Tobon AM, Restrepo A, Colombo AL. Epidemiology of opportunistic fungal infections in Latin America. Clin Infect Dis. 2010;51:561–70.
Benard G, Duarte AJ. Paracoccidioidomycosis: a model for evaluation of the effects of human immunodeficiency virus infection on the natural history of endemic tropical diseases. Clin Infect Dis. 2000;31:1032–9.
Moraes-Vasconcelos D, Grumach AS, Yamaguti A, Andrade ME, Fieschi C, de Beaucoudrey L, Casanova JL, Duarte AJ. Paracoccidioides brasiliensis disseminated disease in a patient with inherited deficiency in the beta1 subunit of the interleukin (IL)-12/IL-23 receptor. Clin Infect Dis. 2005;41:e31–37.
Barrozo LV, Mendes RP, Marques SA, Benard G, Silva ME, Bagagli E. Climate and acute/subacute paracoccidioidomycosis in a hyper-endemic area in Brazil. Int J Epidemiol. 2009;38:1642–9.
Shikanai-Yasuda MA, Telles Filho Fde Q, Mendes RP, Colombo AL, Moretti ML. Guidelines in paracoccidioidomycosis. Rev Soc Bras Med Trop. 2006;39:297–310.
Bellissimo-Rodrigues F, Machado AA, Martinez R. Paracoccidioidomycosis epidemiological features of a 1,000-cases series from a hyperendemic area on the southeast of Brazil. Am J Trop Med Hyg. 2011;85:546–50.
Rolon PA. Paracoccidioidomycosis: an epidemic in the Republic of Paraguay, the center of South America. Mycopathologia. 1976;59:67–80.
McEwen JG, Bedoya V, Patino MM, Salazar ME, Restrepo A. Experimental murine paracoccidioidomycosis induced by the inhalation of conidia. J Med Vet Mycol. 1987;25:165–75.
Morejon KM, Machado AA, Martinez R. Paracoccidioidomycosis in patients infected with and not infected with human immunodeficiency virus: a case–control study. Am J Trop Med Hyg. 2009;80:359–66.
Bethlem EP, Capone D, Maranhao B, Carvalho CR, Wanke B. Paracoccidioidomycosis. Curr Opin Pulm Med. 1999;5:319–25.
Ferreira MS. Paracoccidioidomycosis. Paediatr Respir Rev. 2009;10:161–5.
Brummer E, Castaneda E, Restrepo A. Paracoccidioidomycosis: an update. Clin Microbiol Rev. 1993;6:89–117.
Fonseca ER, Pardal PP, Severo LC. Paracoccidioidomycosis in children in Belem, Para. Rev Soc Bras Med Trop. 1999;32:31–3.
Pereira PM, Akel PB, Lima LL, Kimura EN, Jalkh AP. Multifocal paracoccidioidomycosis: a diagnostic challenge due to late cutaneous manifestation. An Bras Dermatol. 2011;86:149–52.
Brazao-Silva MT, Andrade MF, Franco T, Ribeiro RI, Silva Wdos S, Faria G, Faria PR, Cardoso SV, Loyola AM. Paracoccidioidomycosis: a series of 66 patients with oral lesions from an endemic area. Mycoses. 2011;54:e189–195.
Marques SA. Fungal infections of the mucous membrane. Dermatol Ther. 2010;23:243–50.
Gasparetto EL, Liu CB, de Carvalho Neto A, Rogacheski E. Central nervous system paracoccidioidomycosis: imaging findings in 17 cases. J Comput Assist Tomogr. 2003;27:12–7.
Goldani LZ. Gastrointestinal paracoccidioidomycosis: an overview. J Clin Gastroenterol. 2011;45:87–91.
Odashiro AN, Odashiro PR, Fernandes PI, Leite LV, Odashiro M, Maloney S, Fernandes BF, Di Cesare S, Burnier Jr MN. Eyelid and conjunctival paracoccidioidomycosis simulating carcinoma. Int Ophthalmol. 2011;31:63–7.
Costa DL, Dias-Melicio LA, Acorci MJ, Bordon AP, Tavian EG, Peracoli MT, Soares AM. Effect of interleukin-10 on the Paracoccidioides brasiliensis killing by gamma-interferon activated human neutrophils. Microbiol Immunol. 2007;51:73–80.
Cabral Marques, O., C. Arslanian, R. Nalio Ramos, M. Morato, Paulo Vitor Soeiro, Sonia Jancar MD, Janaíra Fernandes Ferreira, Cristina Worm Weber, Gisele Kuntze, Nelson Augusto Rosario, Beatriz Tavares Costa Carvalho, Patricia Cruz Bergami-Santos, Mary J. Hackett, Hans D. Ochs, Troy R. Torgerson, Jose Alexandre Marzagão Barbuto, and Antonio Condino-Neto. Dendritic cells from X-linked Hyper IgM patients demonstrate defective responses to the fungal pathogens Candida albicans and Paracoccidioides brasiliensis. J Allergy Clin Immunol. 2011 (in press).
Romani L. The T cell response against fungal infections. Curr Opin Immunol. 1997;9:484–90.
Rezaei N, Hedayat M, Aghamohammadi A, Nichols KE. Primary immunodeficiency diseases associated with increased susceptibility to viral infections and malignancies. J Allergy Clin Immunol. 2011;127:1329–41. e1322; quiz 1342–1323.
Banatvala N, Davies J, Kanariou M, Strobel S, Levinsky R, Morgan G. Hypogammaglobulinaemia associated with normal or increased IgM (the hyper IgM syndrome): a case series review. Arch Dis Child. 1994;71:150–2.
Jeena P, Thompson E, Nchabeleng M, Sturm A. Emergence of multi-drug-resistant Acinetobacter anitratus species in neonatal and paediatric intensive care units in a developing country: concern about antimicrobial policies. Ann Trop Paediatr. 2001;21:245–51.
Omwandho CO, Gruessner SE, Tinneberg HR. Early pregnancy loss and neonatal deaths associated with Klebsiella pneumonia infection: a mini review of possible occupational health risk. Arch Gynecol Obstet. 2006;273:258–60.
Soler-Palacin P, Margareto C, Llobet P, Asensio O, Hernandez M, Caragol I, Espanol T. Chronic granulomatous disease in pediatric patients: 25 years of experience. Allergol Immunopathol (Madr). 2007;35:83–9.
Jacobs ZD, Guajardo JR, Anderson KM. XLA-associated neutropenia treatment: a case report and review of the literature. J Pediatr Hematol Oncol. 2008;30:631–4.
Jain N, Euler GL, Shefer A, Lu P, Yankey D, Markowitz L. Human papillomavirus (HPV) awareness and vaccination initiation among women in the United States, National Immunization Survey-Adult 2007. Prev Med. 2009;48:426–31.
Hernandez PA, Gorlin RJ, Lukens JN, Taniuchi S, Bohinjec J, Francois F, Klotman ME, Diaz GA. Mutations in the chemokine receptor gene CXCR4 are associated with WHIM syndrome, a combined immunodeficiency disease. Nat Genet. 2003;34:70–4.
Guerra-Maranhao MC, Costa-Carvalho BT, Nudelman V, Barros-Nunes P, Carneiro-Sampaio MM, Arslanian C, et al. Response to polysaccharide antigens in patients with ataxia-telangiectasia. J Pediatr (Rio J). 2006;82(2):132–6.
Acknowledgments
The authors thank FAPESP (grant 2008/06635-0 to OCM and grant 2008/55700-9 to ACN), CNPq, and Jeffrey Modell Foundation for financial support and the patients and their families for their participation in this study.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Cabral-Marques, O., Schimke, LF., Pereira, P.V.S. et al. Expanding the Clinical and Genetic Spectrum of Human CD40L Deficiency: The Occurrence of Paracoccidioidomycosis and Other Unusual Infections in Brazilian Patients. J Clin Immunol 32, 212–220 (2012). https://doi.org/10.1007/s10875-011-9623-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10875-011-9623-6