
Abstract
The Brazilian Cerrado harbors great floristic and structural diversity composed of a mosaic of natural vegetation types and anthropogenic environments such as introduced pastures. Here, our goal was to evaluate the patterns of taxonomic and functional diversity of dung beetles in a human-modified landscape in Brazilian Cerrado. For this, we evaluated the species richness, species composition and abundance (non-weighted by abundance) and species diversity (Shannon index—weighted by abundance) and three functional indexes (functional richness, functional evenness and functional dispersion). We collected the insects in fragments of Cerrado (Cerrado stricto sensu), riparian forests under pressure of timber removal and livestock and exotic pastures (Urochloa spp.) in Anastácio and Aquidauana, Mato Grosso do Sul, Brazil. We used pitfall traps baited with human feces and fresh capybara dung, a large native rodent, as a representative of the mammal regional fauna. Dung beetle richness was higher in Cerrado fragments, while the abundance was higher in exotic pastures and riparian forests. Species composition differed among vegetation types. Finally, the species diversity and functional diversity did not differ among vegetation types. Thus, we demonstrated the information generated by conceptually similar indexes (e.g. functional richness, species richness and species diversity) may not provide similar information on dung beetles responses to disturbance due to differences in the weighting on species abundance. Finally, the use of taxonomic and functional metrics generate complementary information that can helps us to reach more efficient conclusions in terms of biodiversity conservation and ecosystem functionality in human-modified variegated landscape.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Agriculture and livestock farming has been the one of the major causes of the transformation of natural ecosystems, resulting in landscapes with an environmental mosaic, characterized by fragments of natural areas mixed with anthropized areas (e.g. pastures, agriculture, agroforestry) (Barlow et al. 2016). Human-modified landscapes often result in the loss of native species (Tabarelli et al. 2010; DeClerck et al. 2010). Despite over one quarter of planet terrestrial surface being covered by ecosystems modified by agricultural activities (Foley et al. 2005), some types of anthropized systems may serve as complementary habitats for native species in human-modified landscapes, thus contributing to biodiversity conservation (Costa et al. 2017) and ecosystem function (Santos-Heredia et al. 2018).
The Brazilian Cerrado is the largest, richest and possibly the most threatened tropical savannah on the Planet, being an important hotspot (Myers et al. 2000; Silva and Bates 2002). However, only 7% of this biome is preserved in protected areas (Soares-Filho et al. 2014). This biome harbors great floristic and structural diversity composed of a mosaic of vegetation types (e.g. grasslands, savannas, and forests) (Oliveira-Filho and Ratter 2002), providing high levels of species richness due to the interchange of species among vegetation types (Almeida and Louzada 2009; Gries et al. 2012). However, the deforestation and land use intensification in Brazilian Cerrado are producing variegated landscapes with modified natural environments and anthropogenic environments, such as exotic pastures, which are expanding and negatively impacting the biodiversity of this biome (Marris 2005; Almeida et al. 2011; Ferreira et al. 2014; Queiroz et al. 2017).
Dung beetles (Coleoptera: Scarabaeinae) perform important ecological functions in the habitats in which they live, acting on nutrient cycling and improving soil fertility and control of livestock parasites (Nichols et al. 2008). They are also considered efficient indicators of environmental changes because they undergo changes in community metrics (e.g. abundance, species richness, composition and/or functional diversity) when subjected to environmental changes (Nichols et al. 2007; Barrágan et al. 2011; Bicknell et al. 2014; França et al. 2016; Beiroz et al. 2018) and their collection methods are relatively inexpensive (Gardner et al. 2008). In addition, dung beetles have a high degree of habitat specificity in the Cerrado (Almeida and Louzada 2009; Gries et al. 2012), and can be useful as focal organisms to assess anthropic impacts (Braga et al. 2013; Korasaki et al. 2013; Correa et al. 2016a; França et al. 2016) in this biome.
Different metrics have been used to assess the impacts of land use change on biodiversity. Within the taxonomic diversity, there are metrics that give weight to rare and abundant species differently (Gotelli and Colwell 2001; Chao and Shen 2003). For example, there is no weight for rare and abundant species to calculate species richness (number of species), abundance (number of individuals) and composition (which species are present in a certain area) (Gotelli and Colwell 2001). In contrast, the Shannon-Wiener index is weighted by species abundance to assess species diversity, being based on the contribution of all individuals to their respective species. However, the information would not be dependent only on the total number of individuals and the total number of species, but also on the proportion of the number of individuals of each species, implying that species are not equally likely (Shannon and Wiener 1949). It can reflect species richness and species evenness, being most sensitive to land use change when there is a high number of rare species in the community (Chao and Shen 2003).
Functional diversity is a measure of biodiversity that quantifies the diversity of characters; capturing differences in species morphology and physiology, life history traits, and ecological niches that affect community responses to the disturbance, and consequently, changes in ecosystem functions (Cadotte et al. 2011; Gerisch et al. 2012; Audino et al. 2014; Derhé et al. 2016). Recently, functional diversity is being used to evaluate responses of dung beetle communities to land use change in order to obtain more efficient information in terms ecosystem functionality (Barragán et al. 2011; Audino et al. 2014; Derhé et al. 2016; Gómez-Cifuentes et al. 2017; Beiroz et al. 2018). Although, there is research that has evaluated the taxonomic response (e.g. species richness, composition and abundance) of dung beetle community to land use change (e.g. Almeida et al. 2011; Gries et al. 2012; Martello et al. 2016) and species diversity in different vegetation types (e.g. Almeida and Louzada 2009) in Brazilian Cerrado, the functional patterns of dung beetle communities in this biome has yet to be investigated. In this sense, using only the taxonomic metrics (species composition, species richness, abundance and diversity) of the community to predict changes in the ecosystem could leave important questions on the maintenance of ecosystem functions and services unanswered (Mouillot et al. 2013a, b).
Due to this, our objective was to evaluate the taxonomic and functional patterns of dung beetle communities in a human-modified landscape in the Brazilian Cerrado. We investigated how abundance, species richness, species composition, species diversity and functional diversity are affected by different vegetation types. For this, we collected the insects in Cerrado fragments (Cerrado stricto sensu), riparian forests under pressure of timber removal and livestock and in exotic pastures (Urochloa spp.) to answer the following questions: (1) Do exotic pastures have a higher dung beetle abundance but lower dung beetle richness and species diversity than natural environments (Cerrado and riparian forests)? We expect higher dung beetle abundance in exotic pastures due to increased cattle dung availability, since this resource is abundant, predictable and frequent in Neotropical pastures (Louzada and Carvalho e Silva 2009), and higher dung beetle richness and diversity in natural environments due to greatest resource variability (Driscoll and Weir 2005); (2) Does the species composition differ among the three vegetation types? We expect that species composition is different among the vegetation types due to clear differences in vegetation structure (Halffter and Arellano 2002; Costa et al. 2017) and (3) Does the lower species diversity translate into a reduced functional diversity in exotic pastures? We expect that the reduction of functionally specialized species in exotic pastures results in lower functional diversity. In addition, we discuss the results found using the two metrics; taxonomic (weighted and non-weighted by abundance) and functional diversity, and how it can useful to understand the possible impacts of land use change in the Brazilian Cerrado.
Materials and methods
Study sites and experimental protocol
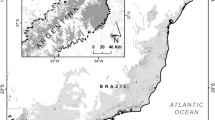
The study was carried out in a region of Cerrado, in the Brazilian municipalities of Anastácio (20°45′45″S, 55°43′49″W) and Aquidauana (19°54′36″S, 55°50′16″W), that are neighbors and are separated by the Aquidauana river. The climate of the region, according to the Köppen classification is Aw, tropical hot-wet, with a rainy summer and a dry winter (Alvares et al. 2014). Annual precipitation ranges from 1200 to 1300 mm and average annual temperature is 26 °C.
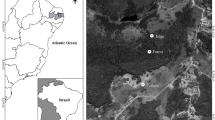
The studied area has been historically modified by livestock farming activities, which generated the variegated landscapes. Overall, the landscape is composed of fragments of Cerrado (stricto sensu), riparian forest under pressure of timber removal and livestock and exotic pastures (Urochloa spp.) (Fig. 1).

Localization of the studied area in Brazil, highlighting the 30 sites of sampling in the Brazilian Cerrado
Cerrado fragments (Cerrado stricto sensu) are characterized by a mixture of plants of two distinct strata: (often 3–8 m; canopy covering 50–90%), and a ground layer composed of grasses, herbs, and small shrubs (Oliveira-Filho and Ratter 2002), with typical wild animals of the Cerrado biome (anteaters, deer, manned wolf, peccaries, rodents, and others). The Cerrado fragments, ranging in size from 3 to 18 ha, are located in a Legal Reserve Area, being conserved with little sign of anthropogenic action.
Riparian forests were degraded initially for timber use and later by the entry of domestic animals (cattle) under pressure from timber removal and livestock. The areas are characterized by a woody layer composed of trees and large shrubs, and there are also wild animals typical of the region. We used a degraded riparian forests because in the study region there were no conserved riparian forests (without high sign of anthropogenic action).
Exotic pasture areas are large open areas, ranging in size from 5 to 20 ha, cultivated with African grasses (Urochloa ssp.), without intensive management (no use of fertilization, and no application of herbicides), with stocking rates between 0.8 and 1.3 livestock per hectare. They are managed in a rotational system where the cattle remain for ~ 45 days before being moved to other pastures and return after ~ 60 days.
We sampled dung beetles in 30 areas, 10 Cerrado areas, 10 areas of degraded riparian forests and 10 exotic pasture areas. The areas within the same system were separated by, at a minimum, 300 m to insure independence of the samples (Silva and Hernandez 2015), while areas of different systems were separated by approximately 1 km. Regions surrounding the sampling sites are dominated by extensive exotic pastures and patches of the Brazilian Cerrado (Puker et al. 2013).
We considered each area as replicates (n = 10 areas) for each system. In each area, we delimited a transect of 400 m located approximately 50 m from the edge habitat, with five sample points separated by 100 m. At each point we installed a set with two traps, one baited with fresh capybara dung (~ 40 g) and the other baited with human feces (~ 40 g), each set of traps from the same point was separated by 20 m, totaling 10 traps per area. Thus, the sample effort was 300 traps, with 100 traps installed in each land use system. Human feces is more appropriate when the target is to sample greater abundance and richness of dung beetles independent of the vegetation structure (Correa et al. 2016b). We also used dung of capybara, a large native rodent, as representative of mammalian fauna that occur frequently in the three studied habitats in order to better sample the community (Puker et al. 2013).
Dung beetle sampling and identification
We sampled dung beetles in December 2011 and January 2012 (middle of the rainy season). In tropical ecosystems, the rainy season is the period of greatest activity and dung beetle richness (Halffter 1991). The traps were composed of a plastic pot (15 cm in diameter, 9 cm high) covered with a plastic cap supported by bamboo sticks. They were installed at ground level, and contained approximately 250 ml of a 1.5% liquid detergent saline solution. The baits were placed in plastic containers (50 mL) in the center of each trap using a wire as a bait holder. Traps remained in the field for 48 h.
The beetles were identified by Dr. Fernando Zagury Vaz-de-Mello. The voucher specimens are deposited in the Entomology Section of the Zoological Collection of the Federal University of Mato Grosso (UFMT, Cuiabá, Mato Grosso, Brazil).
Functional traits
Three functional traits were analyzed that could directly influence the ecological functions performed by dung beetles: habit resource allocation (tunnelers, rollers or dwellers), diet preference (human feces, capybara dung or generalist feces) and body size (Supplementary Material).
Data analysis
Pattern of species richness, abundance and diversity
To assess the completeness of our dung beetle survey in the three habitats, we calculated the number of observed species (Sobs) as a percentage of the total species richness, which was estimated based on the average of four abundance based nonparametric estimators: ACE, CHAO1, JACK1 and BOOTSTRAP, using the formula: Sampling efficiency = [Sobs × 100/((ACE + CHAO1 + JACK1 + BOOTS)/4)]. The richness estimates were calculated with the software EstimateS v. 9.1.0, with 999 randomizations (Colwell 2013).
To evaluate differences in dung beetle abundance, species richness and species diversity (Shannon index) among habitats, we used Generalized Linear Models (GLMs). All GLMs were submitted to residue analysis, in order to evaluate the adequacy of the distribution of errors (Crawley 2002). We used Poisson errors corrected for over-dispersion (quasi-Poisson) for dung beetle abundance and richness. All analyzes were performed using the R v3.3.1 (R Development Core Team 2016).
Species composition and dispersion patterns
To graphically express the changes in dung beetle community composition among habitats we used a non-metric multidimensional scaling analysis (NMDS) based on Bray–Curtis dissimilarity matrix with standardized and square-root transformed abundance data (Anderson and Willis 2003). To verify these differences we used a permutational multivariate analysis of variance (PERMANOVA) (Anderson 2001). To test heterogeneity of multivariate dispersions between different habitat types, we used permutational multivariate analysis of dispersion (PERMDISP) (Anderson 2001). NMDS, PERMANOVA and PERMDISP were performed using the Primer v.6 software with PERMANOVA+ (Clarke and Gorley 2006).
Functional diversity
To calculate three functional diversity indexes, we used the dbFD function of the ‘FD’ package (R Development Core Team 2016): (1) Functional richness (FRic) represents the range of traits in a community quantified by the volume of functional trait space occupied; (2) Functional evenness (FEve) summarizes how species abundances are distributed along the occupied functional space (Villéger et al. 2008); and (3) Functional dispersion (FDis) which indicates the distribution of abundances in the space of functional traces in relation to a weighted centroid in abundance and the volume of space occupied (Laliberté and Legendre 2010).
To evaluate differences in the three functional diversity indexes among habitats, we also used GLMs with gaussian errors. All GLMs were submitted to residual analysis, in order to evaluate the adequacy of the distribution of errors (Crawley 2002). All analyzes were performed using the R v3.3.1 (R Development Core Team 2016).
Results
Patterns of species richness, abundance and species diversity
In the Cerrado, we captured 30 species and 2577 individuals; 26 species and 6020 individuals in the riparian forests; and 26 species and 6224 individuals in the exotic pastures (Table 1). The Cerrado shared 24 species with riparian forest, and 23 species with exotic pastures. Riparian forest pastures shared 19 species with exotic pastures.
The four species richness estimators indicate high sampling efficiency, with 91% in the Cerrado, 87% in the riparian forests and 67% in the exotic pastures (Table S1). The species accumulation curves suggest that the sampling effort was adequate to characterize the local dung beetle community (Fig. 2).

Individual-based species accumulation curves for dung beetles sampled in two native vegetation and exotic pastures in Brazilian Cerrado. Shaded areas indicate 95% confidence intervals
In the Cerrado, of the 2577 individuals sampled, the dominant species were: Uroxys sp. (22.82%), Onthophagus aff. hirculus (16.41%) and Canthon aff. tristis (16.06%) (Table 1). In riparian forests, of the 6020 individuals sampled, the dominant species were: Trichillum externepunctatum Preudhomme de Borre (59.87%), Dichotomius opacipennis (Luederwaldt) (7.66%) and Onthophagus aff. hircullus (7.49%). In exotic pastures, of the 6224 individuals sampled, the dominant species were: Uroxys sp. (34.77%), Dichotomius bos (Blanchard) (16.96%) and Ontherus appendiculatus (Manerrheim) (19.84%) (Table 1).
The species richness was higher in the Cerrado, and similar in the riparian forests and exotic pastures (F(2,27) = 4.01, p = 0.03) (Fig. 3). Dung beetle abundance was lower in the Cerrado and similar in exotic pastures and riparian forests (F(2,27) = 3.77, p = 0.03) (Fig. 3). Dung beetle diversity did not differ among the systems (F(2,27) = 1.76, p = 0.19) (Fig. 4).

Mean abundance and species richness of dung beetle communities in three different habitats in the Brazilian Cerrado. Different letters above the bars indicate statistically significant differences among habitats (p < 0.05)

Mean diversity (Shannon index) of dung beetle communities in three different habitats in the Brazilian Cerrado. Identical letters above the bars indicate that there were no significant differences among the habitats (p > 0.05)
Species composition
The dung beetle species composition differed among the three vegetation types, each habitat forming distinct groups (PERMANOVA, pseudo-F = 8.08, p = 0.001) (Fig. 5; Table 2; Table S3). Vegetation types showed no differences in the multivariate dispersion of points (PERMDISP, F = 0.45, p = 0.68) (Table 2).

Non-metric multidimensional scaling results (NMDS) constructed from Bray–Curtis matrices, for dung beetle communities in three different habitats in the Brazilian Cerrado
Functional diversity
The functional richness (F(2,27) = 2.28, p = 0.09), functional evenness (F(2,27) = 0.67, p = 0.52) and functional dispersion (F(2,27) = 0.44, p = 0.64) did not differ among the three habitats (Fig. 6).

Mean ± SE of functional dispersion (FDis), functional evenness (FEve) and functional richness (FRic) of dung beetle communities in three different habitats in the Brazilian Cerrado: identical letters above the bars indicate that there were no significant differences among the habitats (p > 0.05)
Discussion
Patterns of species richness, abundance and diversity
This study evaluated, for the first time, the patterns of taxonomic and functional diversity of dung beetles among different vegetation types in Brazilian Cerrado. Despite the differences in sample effort and sampling protocols, we recorded a dung beetle community in Brazilian Cerrado which presented a larger number of species in Cerrado fragments and exotic pastures, when compared to previous studies in the same region (i.e. Correa et al. 2016a, b; Cerrado: S = 38 and exotic pastures: S = 24 species). Although our sampling efficiency was high in Cerrado and Riparian forest, above 85%, in exotic pastures it was low. This can be related the high number of rare species sampled in exotic pastures (e.g. singletons or doubletons) (Gotelli and Colwell 2001).
The species richness was higher in Cerrado fragments. Dung beetle richness is influenced by the vegetation structure (Hanski and Cambefort 1991); in particular the richness of the land use system (Lobo and Martín-Piera 2002) and also of the vertebrate fauna (Estrada et al. 1999), especially richness of large mammals (Nichols et al. 2013). Anthropic actions (e.g. land use change, selective logging, grazing and others) alter environmental conditions that directly affect species biology, such as luminosity, temperature and humidity, resulting in biodiversity loss (Hanski and Cambefort 1991). Studies have demonstrate that vegetation type is a key factor in the organization of dung beetle communities in tropical landscapes, where more simplified environments, such as exotic pastures, have lower species richness (Braga et al. 2013; Korasaki et al. 2013; Correa et al. 2016a). This can explain the lower dung beetle richness in riparian forests of our study. Although riparian forests are a natural system, the riparian forests of our study have been suffering anthropogenic actions, which may be negatively affecting the dung beetle community (Viegas et al. 2014). Riparian forests show very different characteristics from more open areas such as pastures and Cerrado, this was corroborated by the differentiated composition among these three systems, furthermore, these areas help to maintain biodiversity and their associated ecological functions in agricultural landscapes (Gray et al. 2014).
The riparian forest shared 24 species with Cerrado, indicating that, despite presenting very different vegetation characteristics, the Cerrado fragments can be used as refuge for dung beetles species from riparian forest. The Exotic pastures shared 23 species with Cerrado fragments and 19 with riparian forests, evidencing that dung beetle richness in exotic pastures may be increased by species of the natural systems. Although, in our study, there was the high sharing between natural habitats and pastures, many species are with low abundance in pastures, which may mean that they are just using these pastures as transitional areas (e.g. ecological corridors or step-stones; Almeida et al. 2011). However, the species that could colonize the pastures in the Brazilian Cerrado could provide important ecosystem functions and services for livestock development, such as dung removal (Nichols et al. 2008), fly parasites control (Braga et al. 2012) and gastrointestinal worms (Nichols and Gómez 2014). These findings suggest that the conservation of natural systems may be a primary source of biodiversity for dung beetles in exotic pastures in Brazilian Cerrado. In this context, the management of pasture areas together with Cerrado and riparian forests areas, or agroforestry systems, may be an alternative to avoid the loss of dung beetle biodiversity. In addition, some actions to increase the biological value of pastures, such as diversification of use, with horse and cattle (Louzada and Carvalho e Silva 2009) or other domestic animals such as sheep (Correa et al. 2013) can aid in the dung beetles conservation.
The greatest abundance of dung beetles in exotic pastures compared with Cerrado fragments was due to the dominance of Uroxys sp. and D. bos in exotic pastures. These two dominant species are probably favored by forage capacity in exotic pastures for cattle dung, resulting in a competitive advantage for some species in this new food source (Louzada and Carvalho e Silva 2009). In addition, dominant species in pastures can develop under high temperatures, intense sun exposure (Navarrete and Halffter 2008), and cattle compacted soils (Halffter et al. 1992). However, dung beetle abundance was similar between exotic pastures and riparian forests, due to the dominance T. externepuctatum in riparian forests. Although pastures are simple and homogeneous systems (Louzada and Carvalho e Silva 2009), and the forests show modifications due to anthropogenic action, they can offer advantages to opportunistic species and species more tolerant to the changes in the environmental dynamics that result in an increase of its population, surpassing the communities of the most conserved natural system (Escobar et al. 2007).
Dung beetle diversity did not differ among systems. Despite there being the highest dung beetle species richness in Cerrado, there was highest abundance in Riparian forest and exotic pastures. This result demonstrates that, besides species richness, species evenness is also very important to maintain a high diversity (Gorelick 2006). Rodrigues et al. (2013) comparing dung beetle diversity among three land uses (pastures, agriculture and forest) found the highest diversity in pastures. Thus, the increased resource supply and distribution during a longer and constant time for the dung beetles (cattle dung) in pastures and the adaptability of dung beetles to the modified environments (Riparian forest) can explain the high diversity of dung beetles in these systems.
Species composition and dispersion patterns
The dung beetle species composition differed among the three vegetation types, while the dispersion patterns are similar among them. Our results demonstrate well-defined communities in each vegetation type, highlighting the importance of each of them for conserving diversity of dung beetles in the landscape studied. We recorded a segregation pattern of species composition between natural environments and exotic pastures; and between different natural vegetation types which also has been documented in other studies on dung beetle communities in Brazilian Cerrado (e.g. Almeida and Louzada 2009; Almeida et al. 2011; Gries et al. 2012), evidencing the high degree of habitat specificity of dung beetles in the Cerrado biome (Almeida and Louzada 2009; Gries et al. 2012). The difference in species composition may be associated with microclimatic factors (biotic and/or abiotic factors) that each environment provides for dung beetles (Hanski and Cambefort 1991). The maintenance of dung beetles in a habitat is regulated by the sensitivity of adults and larvae to factors such as temperature and humidity (Larsen 2012). Thus, microclimatic changes are considered key factors in the impoverishment and loss of dung beetle biodiversity, limiting the species dispersal and colonization capacity, resulting in the eventual extinction of habitat specialists (Gardner et al. 2008).
Functional diversity
We did not find differences for the functional diversity indexes FRic, FEve and FDis among the vegetation types. Differences in taxonomic and functional patterns may be a result of functional redundancy between species in native vegetation and exotic pastures; or replacement by functionally different species that could maintain similar FRic values (Rosenfeld 2002; Magnago et al. 2014). Thus, despite the higher dung beetle richness in the Cerrado fragments, the high rates of species sharing among vegetation types may be a good predictor of the maintenance of functional diversity in the landscape, causing possible functional redundancy of species. For dung beetles is common for species to be grouped in the extremes (high originality and low uniqueness), losing species without decreasing FRic (Beiroz et al. 2018). Therefore, even with the species richness reduction in pastures, it may not have occurred the loss of functionally specialized species, not leading to reduction of functional diversity (Villeger et al. 2008). As for the FEve, an explanation for this pattern is that the dung beetle species of the Brazilian Cerrado can present characteristics that allow them to tolerate the environmental conditions created by the disturbance. Similar FEve values between natural and anthropized sites may suggest that niche space occupied in anthropogenic areas is really being exploited by species and not only underutilized (Audino et al. 2014). Finally, similar values of FD is among habitats may indicate a greater dispersion of functional traits in anthropic areas, corresponding to a gain in the variability of responses to disturbances between species that contribute in a similar way to the ecosystem function (Laliberté et al. 2010).
Complementarity between the results evaluating taxonomic (weighted and non-weighted by abundance) and functional diversity
In order to predict common or divergent changes in functional and taxonomic diversity, it is necessary to use different weights for rare and abundant species. However, many common species make unique and persistent contributions to functional diversity (Chapman et al. 2018). We found different patterns of taxonomic (non-weighted by abundance) and functional diversity. Recently, Beiroz et al. (2018) demonstrated that taxonomic metrics (species richness) are more sensitive than functional metrics in detecting the effects of strong habitat modification in dung beetle communities (e.g. eucalyptus plantation). However, the responses depend mainly on the intensity of the disturbed and the functional characteristics chosen (Mlambo 2014; Beiroz et al. 2018). A low intensity disturbance in highly diversified communities does not modify functional structure, but may alter species composition. On the other hand, high intensity disturbances tend to negatively affect both taxonomic and functional components of the local biodiversity (Mlambo 2014; Magnago et al. 2014). In contrast, we found similar patterns of taxonomic (weighted by abundance) and functional diversity. In ecological applications, the true number of species is often unknown and some rare species may not be discovered in a sample of individuals (Gotelli and Colwell 2001; Chao and Shen 2003). Since rare species usually contribute more to functional diversity than common species (Chapman et al. 2018), metrics that ignore the importance of species abundance may underestimate the impacts (Moullit et al. 2013a, b). In this sense, in contrast to species richness, species diversity is most sensitive to land use change when there is a high number of rare species in the community (Chao and Shen 2003). Thus, we demonstrated the information generated by conceptually similar indexes (e.g. FRic, species richness and species diversity) may influenced by differences in the weighting on species abundance, providing different information on dung beetles responses to disturbance.
Studies on the response animal groups that provide important services to the ecosystem are important to supply baselines for conservation policies, which may help protect tropical savannas around the world. Thus, we demonstrate the importance of the presence of native vegetation areas, besides conserving high taxonomic diversity, also favors species sharing by maintaining a large number of dung beetles in the exotic pastures. This may help to maintain the taxonomic (Barlow et al. 2007; Rös et al. 2012; Costa et al. 2017; Beiroz et al. 2018) and functional diversity of dung beetles in the human-modified variegated landscape (Beiroz et al. 2018), mitigating the possible effects of land use change on the Brazilian Cerrado. Although, in our study, the patterns of taxonomic (non-weighted by abundance) and functional diversity of dung beetles are different, these two metrics generate complementary information that can helps us to reach more efficient conclusions in terms of biodiversity conservation, as well as ecosystem functionality in human-modified variegated landscape (Beiroz et al. 2018).
References
Almeida SSP, Louzada JNC (2009) Estrutura da comunidade de Scarabaeinae (Scarabaeidae: Coleoptera) em fitofisionomias do Cerrado e sua importância para a conservação. Neotrop Entomol 38:32–43
Almeida S, Louzada J, Sperber C, Barlow J (2011) Subtle-use change and tropical biodiversity: dung beetle communities in Cerrado grasslands and exotic pastures. Biotropica 43:704–771
Alvares CA, Stape JL, Sentelhas PC, Gonçalves JLM, Sparovek G (2014) Köppen’s climate classification map for Brazil. Meteorol Z 22:711–728
Anderson MJ (2001) A new method for non-parametric multivariate analysis of variance. Austral Ecol 26:32–46
Anderson MJ, Willis TJ (2003) Canonical analysis of principal coordinates: a useful method of constrained ordination for ecology. Ecology 84:511–525
Audino LD, Louzada J, Comita L (2014) Dung beetles as indicators of tropical forest restoration success: is it possible to recover species and functional diversity? Biol Conserv 169:248–257
Barlow J, Gardner TA, Araujo IS et al (2007) Quantifying the biodiversity value of tropical primary, secondary and plantation forests. PNAS 104:18555–18560
Barlow J, Lennox GD, Ferreira J et al (2016) Anthropogenic disturbance in tropical forests can double biodiversity loss from deforestation. Nature 7535:144–147
Barragán F, Moreno CE, Escobar F, Halffter G, Navarrete D (2011) Negative impacts of human land use on dung beetle functional diversity. PLoS ONE 6:e17976
Beiroz W, Sayer E, Slade SM, Audino L, Braga RF, Louzada J, Barlow J (2018) Spatial and temporal shifts in functional and taxonomic diversity of dung beetle in a human-modified tropical forest landscape. Ecol Ind 95:418–526
Bicknell JE, Phelps SP, Davies RG, Mann DJ, Struebig MJ, Davies GD (2014) Dung beetles as indicators for rapid impact assessments: evaluating best practice forestry in the neotropics. Ecol Ind 43:154–161
Braga RF, Korasaki V, Audino LD, Louzada J (2012) Are dung beetles driving dung-fly abundance in traditional agricultural areas in the Amazon? Ecosystems 15:1173–1181
Braga RF, Korasaki V, Andresen E, Louzada J (2013) Dung beetle community and functions along a habitat-disturbance gradient in the Amazon: a rapid assessment of ecological functions associated to biodiversity. PLoS ONE 8:e5778
Cadotte MW, Carscadden K, Mirotchnick N (2011) Beyond species: functional diversity and the maintenance of ecological processes and services. J Appl Ecol 48:1079–1087
Chao A, Shen T-J (2003) Nonparametric estimation of Shannon’s index of diversity when there are unseen species in sample. Environ Ecol Stat 10:429–443
Chapman ASA, Tunnicliffe V, Bates AE (2018) Both rare and common species make unique contributions to functional diversity in an ecosystem unaffected by human activities. Divers Distrib 24:568–578
Clarke KR, Gorley RN (2006) Primer v6 Permanova+. Primer-E Ltd., Plymouth
Colwell RK (2013) EstimateS: Statistical estimation of species richness and shared species from samples
Correa CMA, Puker A, Korasaki V, Oliveira NG (2013) Dung beetles (Coleoptera, Scarabaeinae) attracted to sheep dung in exotic pastures. Rev Bras Entomol 57:113–116
Correa CMA, Puker A, Ferreira KR, Cristaldo CM, Ferreira FNF, Abot AR, Korasaki V (2016a) Using dung beetles to evaluate the conversion effects from native to introduced pasture in the Brazilian Pantanal. J Insect Conserv 20:447–456
Correa CMA, Puker A, Korasaki V, Ferreira KR, Abot AR (2016b) The attractiveness of baits to dung beetles in Brazilian Cerrado and exotic pasturelands. Entomol Sci 19:112–123
Costa C, Oliveira VHF, Maciel R, Beiroz W, Korasaki V, Louzada J (2017) Variegated tropical lndscapes conserve diverse dung betle communities. PeerJ 5:e3125
Crawley MJ (2002) Statistical computing—an introduction to data analysis using s-plus. Wiley, London
DeClerck FAJ, Chazdon R, Holl KD et al (2010) Biodiversity conservation in human-modified landscapes of Mesoamerica: present, past and future. Biol Conserv 143:2301–2313
Derhé MA, Murphy H, Monteith G, Menéndez R (2016) Measuring the sucess of restoration for restoring biodiversity and ecosystem functioning. J Appl Ecol 53:1714–1724
Driscoll DA, Weir T (2005) Beetle responses to habitat fragmentation depend on ecological traits, habitat condition and remnant size. Conserv Biol 19:182–194
Escobar F, Hallfter G, Arellano L (2007) From forest to pasture: an evaluation of the influence of environment and biogeography on the structure of dung beetle (Sscarabaeinae) assemblages along three altitudinal gradients in the Neotropical region. Ecography 30:193–208
Estrada A, Anzures DA, Coates-Estrada R (1999) Tropical rain forest fragmentation, howler monkeys (Alouatta palliata), and dung beetles at Los Tuxtlas, Mexico. Am J Primatol 48:253–262
Ferreira J, Aragao LEOC, Barlow J et al (2014) Brazil’s environmental leadership at risk. Science 346:706–707
Foley JA, DeFries R, Asner GP et al (2005) Global consequences of land use. Science 309:570–574
França F, Louzada J, Korasaki V, Griffiths H, Silveira JM, Barlow J (2016) Do space-for-time assessments underestimate the impacts of logging on tropical biodiversity? An Amazonian case study using dung beetles. J Appl Ecol 53:1098–1105
Gardner TA, Barlow J, Araujo IS et al (2008) The cost-effectiveness of biodiversity surveys in tropical forests. Ecol Lett 11:139–150
Gerisch M, Agostinelli V, Henle K, Dziock F (2012) More species, but all do the same: contrasting effects of flood disturbance on ground beetle functional and species diversity. Oikos 121:508–515
Gómez-Cifuentes A, Munevar A, Gimenez VC, Gatti MG, Zurita GA (2017) Influence of land use on the taxonomic and functional diversity of dung beetles (Coleoptera: Scarabaeinae) in the southern Atlantic forest of Argentina. J Insect Conserv 21:147–156
Gorelick R (2006) Combining richness and abundance into a single diversity index using matrix analogues of Shannon’s and Simpson’s indices. Ecography 29:525–530
Gotelli NJ, Colwell RK (2001) Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecol Lett 4:379–391
Gray CL, Slade EM, Mann DJ, Lewis TO (2014) Do riparian reserves support dung beetle biodiversity and ecosystem services in oil palm-dominated tropical landscape? Ecol Evol 4:1049–1060
Gries R, Louzada J, Almeida S, Macedo R, Barlow J (2012) Evaluating the impacts and conservation value of exotic and native tree afforestation in Cerrado grasslands using dung beetles. Insect Conserv Divers 5:175–185
Halffter G (1991) Historical and ecological factors determining the geographical distribution of beetles (Coleoptera. Scarabaeidae. Scarabaeinae). Folia Entomol Mex 82:195–238
Halffter G, Arellano L (2002) Response of dung beetle diversity to human-induced changes in a tropical landscape. Biotropica 34:144–154
Halffter G, Favila ME, Halffter V (1992) A comparative study of the structure of the scarab guild in Mexican tropical rain forest and derived ecosystem. Folia Entomol Mex 84:131–156
Hanski I, Cambefort Y (1991) Dung beetle ecology. Princeton University Press, Princeton
Korasaki V, Braga RF, Zanetti R, Moreira FMS, Vaz-de-Mello FZ, Louzada J (2013) Conservation value of alternative land-use systems for dung beetles in Amazon: valuing traditional farming practices. Biodivers Conserv 22:1485–1499
Laliberté E, Legendre P (2010) A distance-based framework for measuring functional diversity from multiple traits. Ecology 91:299–305
Laliberté E, Wells JA, Declerck F et al (2010) Land-use intensification reduces functional redundancy and response diversity in plant communities. Ecol Lett 13:76–86
Larsen TH (2012) Upslope range shifts of Andean dung beetles in response to deforestation: compounding and confounding effects of microclimatic change. Biotropica 44:82–89
Lobo JM, Martín-Piera F (2002) Searching for a predictive model for species richness of Iberian dung beetle based on spatial and environmental variables. Conserv Biol 16:158–173
Louzada JNC, Carvalho e Silva PR (2009) Utilisation of introduced Brazilian pastures ecosystems by native dung beetles: diversity patterns and resource use. Insect Conserv Divers 2:45–52
Magnago LFS, Edwards DP, Edwards FA, Magrach A, Martins SV, Laurence WF (2014) Functional attributes change but functional richness is unchanged after fragmentation of Brazilian Atlantic forests. J Ecol 102:465–485
Marris E (2005) Conservation in Brazil: the forgotten ecosystem. Nature 437:944–945
Martello F, Andriolli F, Souza TB, Dodonov P, Ribeiro MC (2016) Edge and land use effects on dung beetles (Coleoptera: Scarabaeidae: Scarabaeinae) in Brazilian cerrado vegetation. J Insect Conserv 20:967–970
Mlambo MC (2014) Not all traits are ‘functional’: insights from taxonomic and biodiversity-ecosystem functioning research. Biodivers Conserv 23:781–790
Mouillot D, Bellwood DR, Baraloto C et al (2013a) Rare species support vulnerable functions in high-diversity ecosystems. PLoS Biol 11:e1001569
Mouillot D, Graham NAJ, Villéger S, Mason NWH, Bellwood DR (2013b) A functional approach reveals community responses to disturbances. Trends Ecol Evol 28:167–177
Myers N, Mittermeier RA, Mittermeier CG, Fonseca GAB, Kent J (2000) Biodiversity hotspots for conservation priorities. Nature 403:853–858
Navarrete D, Halffter G (2008) Dung beetle (Coleoptera: Scarabaeidae: Scarabaeinae) diversity in continuous forest, forest fragments and cattle pastures in a landscape of Chiapas, Mexico: the effects of anthropogenic changes. Biodivers Conserv 17:2869–2898
Nichols E, Gómez A (2014) Dung beetles and the epidemiology of parasitic nematodes: patterns, mechanisms and questions. Parasitology 141:614–623
Nichols E, Larsen T, Spector S, Davis AL, Escobar F, Favila M, Vulinec K (2007) Global dung beetle response to tropical forest modification and fragmentation: a quantitative literature review and meta-analysis. Biol Conserv 137:1–19
Nichols E, Spector S, Louzada JNC, Larsen TS, Favila M, The Scarabaeinae Research Network (2008) Ecological functions and services provided by Scarabaeinae dung beetles. Biol Conserv 141:1461–1474
Nichols E, Uriarte M, Peres CA, Louzada J, Braga RF, Schiffler G et al (2013) Human-induced trophic cascades along the fecal detritus pathway. PLoS ONE 8:e75819
Oliveira-Filho T, Ratter JA (2002) Vegetation physiognomies and woody flora of the Cerrado biome. In: Oliveira PS, Marquis RJ (eds) The Cerrados of Brazil—ecology and natural history of a neotropical Savanna. Columbia University Press, New York, pp 91–120
Puker A, Correa CMA, Korasaki V, Ferreira KR, Oliveira NG (2013) Dung beetles (Coleoptera: Scarabaeidae) attracted to dung of the largest herbivorous rodent on Earth: a comparison with human feces. Environ Entomol 42:1218–1225
Queiroz ACM, Rabello AM, Braga DL et al (2017) Cerrado vegetation types determine how land use impacts ant biodiversity. Biodivers Conserv. https://doi.org/10.1007/s10531-017-1379-8
R Development Core Team (2016) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. http://www.RProject.org
Rodrigues MM, Uchôa MA, Ide S (2013) Dung beetle (Coleoptera: Scarabaeoidea) in three landscapes in Mato Grosso do Sul, Brazil. Braz J Biol 73:211–220
Rös M, Escobar F, Halffter G (2012) How dung beetles respond to a human-modified variegated landscape in Mexican cloud forest: a study of biodiversity integrating ecological and biogeographical perspectives. Divers Distrib 18:377–384
Rosenfeld JS (2002) Functional redundancy in ecology and conservation. Oikos 98:156–162
Santos-Heredia C, Andresen E, Zárate DA, Escobar F (2018) Dung beetles and their ecological functions in three agroforestry systems in Lacandona rainforest of México. Biodivers Conserv 27:2379–2394
Shannon CE, Wiener W (1949) The mathematical theory of communication. Illinois Press, Urbana
Silva JMC, Bates JM (2002) Biogeographic patterns and conservation in the South American Cerrado: a tropical savanna hotspot. Bioscience 52:225–233
Silva PG, Hernandez MIM (2015) Spatial patterns of movement of dung beetle species in a tropical forest suggest a new trap spacing for dung beetle biodiversity studies. PLoS ONE 10(5):e012611
Soares-Filho B, Rajão R, Macedo M, Carneiro A, Costa W, Coe M, Rodrigues H, Alencar A (2014) Cracking Brazil’s forest code. Science 344:363–364
Tabarelli M, Aguiar AV, Ribeiro MC, Metzeger JP, Peres AC (2010) Prospects for biodiversity conservation in the Atlantic forest: lessons from aging human-modified landscapes. Biol Conserv 143:2328–2340
Viegas G, Stenert C, Schulz UH, Maltchik L (2014) Dung beetle communities as biological indicators of riparian forest widths in southern Brazil. Ecol Ind 36:703–710
Villéger S, Mason NWH, Mouillot D (2008) New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 89:2290–2301
Acknowledgements
We thank the father of the first author, Agenor Martinho Correa, for the encouragement and logistical support, and FAPEMIG (APQ—02696-15) for the financial support. We also thank Fernando Z. Vaz-de-Mello (UFMT) for identification of the dung beetle species; Cezar F. Borges for the map confection, Cleilsom M. Cristaldo, Naiara G. Oliveira and Kleyton R. Ferreira, for the field support; Mrs. Marcos Crestani, Gélio P. Brum and Jorge de Deus Ricardo for access to their properties, Wallace Beiroz and three anonymous reviewers for the fruitful comments on the manuscript. CMAC received a PhD scholarschip from the Conselho Nacional de Desenvolvimento Científico Tecnológico (CNPq, Brazil) (140741/2015-1) from the Entomology Graduate Program, Universidade Federal de Lavras, and PhD sandwich scholarship from the Coordenação de Aperfeiçoamento de Pessoa de Nível Superior (CAPES, Brazil) (88881.134292/2016-01).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
There is no conflict of interest among authors.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Correa, C.M.A., Braga, R.F., Puker, A. et al. Patterns of taxonomic and functional diversity of dung beetles in a human-modified variegated landscape in Brazilian Cerrado. J Insect Conserv 23, 89–99 (2019). https://doi.org/10.1007/s10841-018-00118-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10841-018-00118-6