
Abstract
Spiders are omnipresent, occurring in almost all terrestrial habitats. They are generalist predators and important for conservation biocontrol in agricultural fields, helping to reduce pesticide applications. In this study, we examined how spider families and spider web types in rice fields are related to local and landscape management. Samples were taken in differently managed rice fields adjoining either homegarden polycultures or banana monocultures. Furthermore, landscape structure, prey abundance, herb richness and cover were taken into account. The results showed that prey availability explained most of the variation in spiders and their web’s abundance, indicating that spiders in the rice fields can potentially contribute to pest control. Adjacent habitat had no effect on the spider populations, but maintaining fallow fields in the surrounding landscape seems to be a useful measure to promote Erigoninae in rice fields. There was no evidence that local management practices such as fertiliser and pesticide use had an impact on spider families, which appeared to be due to the low level of these inputs. Spider web sampling can complement spider sampling as it detects spiders hidden at the base of the rice tillers, which are likely to be missed by sweep netting. Additionally, tetragnathid webs are easy to observe and thus can be used as an indication for farmers not to spray pesticides as spiders are potentially controlling the pest species. Interviews with farmers made clear that many farmers in the study area showed their interest in management methods that promote biological pest control.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
In agricultural fields, spiders often occur in high abundance and richness (e.g. Marc et al. 1999; Sunderland and Samu 2000; Sebastian et al. 2005; Stenchly et al. 2012; Takada et al. 2012). Their broad food spectrum leads to stable population dynamics and allows them to remain in agricultural fields, especially in times of low abundances of preferred prey species (Maloney et al. 2003). Spiders as a group are generalist predators. However, different families, genera or species utilise various foraging strategies and exhibit specialisations with respect to microhabitat, prey items or activity periods. Spiders generally can be divided into web builders and free hunters, while Uetz et al. (1999) even distinguished between eight guilds in agricultural fields: sheet, tangle, orb and space web-building spiders and running, stalking, ambushing and foliage-dwelling free-hunting species. Hence, they can be very useful natural enemies for insect pests (Marc et al. 1999; Maloney et al. 2003). Thus, their conservation in agricultural fields is important for biological pest control (Settle et al. 1996; Lou et al. 2013) and contributes to environmentally friendly farming practices.
Rice (Oryza sativa) is the major food crop in Asia. To meet the demands of a growing population the ‘Green Revolution’, a most influential intervention in rice cultivation in all Asian countries starting in the mid-1960s, aimed to increase rice production through the introduction of agrochemicals, machinery and high yielding varieties. However, intensified cultivation practices may affect the conservation of prey-predator complexes in agricultural fields (Zhao et al. 2015; Lee et al. 2014). Fertiliser application can promote pest populations; this is especially the case with sap-feeding insects like plant- and leafhoppers, which respond positively to nitrogen-rich plants (Matson 1997; Lu and Heong 2009; Horgan and Crisol 2013). Pesticide application kills the target species but often it also kills useful natural enemies (Settle et al. 1996; Marc et al. 1999; Landis et al. 2000; Tilman et al. 2001). For instance, pesticides applied in the early growth of rice plants kill predators like spiders, which are then unable to control pests that peak later in the season, such as plant- and leafhoppers (hereafter referred to as leafhoppers), which are a major concern in rice cultivation (Settle et al. 1996; Lou et al. 2013; Lu et al. 2015). To avoid such secondary pest outbreaks, conservation biological pest control might be the better strategy instead of applying chemical pesticides. By conserving predators such as spiders in rice fields, the pest numbers can be held below the economic threshold (Settle et al. 1996). According to Balfour and Rypstra (1998) and Sunderland and Samu (2000), weed density provides higher structural diversity in a habitat, which benefits spider communities. Weeding therefore may reduce this structural diversity in the crop field and thus also can lead to a decline in spider abundance and richness.
Besides measures at a local scale, the features at the landscape scale can be equally important for the protection of predators in agricultural fields. Diverse natural or semi-natural habitats adjoining crop field as well as a more complex landscape structure containing comparatively undisturbed habitats, e.g. fallow fields, field margins or hedgerows, can provide refuges or source habitats for various species (Bianchi et al. 2006). Hence, these habitats are important for the colonisation of crop fields by predators such as spiders (Marc et al. 1999; Schmidt et al. 2008; Schoenly et al. 2010). However, expanding agricultural land, establishing monocultures and shifting to new crop types at the expenses of natural or semi-natural habitats are common phenomena in agriculturally used areas (Tilman et al. 2001; Laurance 2010).
In this study, we examined how different spider families as well as spider webs in rice fields are related to prey availability, adjacent habitat, landscape structure and cultivation practices. Understanding the relative role of local and landscape management for enhancing spider densities helps to develop strategies for conservation of these important biocontrol agents.
Materials and methods
Study site
The study took place in the Wayanad district, Kerala State, South India. Wayanad is an undulating plateau located in the Western Ghats between the Kerala plains in the west and the Mysore plateau in the east. The hilly terrain ranges between 700 and 2100 m a.s.l. The climate is classified as a tropical monsoon climate with a mean temperature range of 18–29 °C and an annual rainfall of 2322 mm. Agriculture is the major source of livelihood in Wayanad. Most of the agricultural land is maintained by subsistence farmers and small-holder plantations (Santhoshkumar and Ichikawa 2010). In diverse homegardens on the hill tops, surrounding the farm houses, farmers grow fruit trees, coffee (Coffea spp.), spices, vegetables, coconut (Cocos nucifera), etc. for self-sufficiency (Kumar et al. 2010). However, market pressure often leads to transformation of these homegardens into simplified systems such as rubber (Ficus elastica) or coffee plantations. Rice is cultivated in the plains surrounding the hills. The rice cultivation in this area is mainly rain-fed and therefore usually only one crop per year is possible. Cultivation starts in July after the monsoon rain and ends with the harvest in December. The ‘Green Revolution’ also affected this rather remote area of rice cultivation (Pandey et al. 2010). Furthermore, as a consequence of the commercialisation of agriculture in the late 1990s, the cultivation of cash crops such as banana (Musa spp.), arecanut (Areca catechu), ginger (Zingiber officinale) and turmeric (Curcuma longa) increased considerably and contributed largely to the foreign exchange earnings of the district (George and Krishnaprasad 2006). These cash crops are usually grown in transformed rice fields. Hence, the expansion of cash crops resulted in a reduction of the rice cultivation area from 30,000 ha (1980–1981) to 8995 ha (2011–2012) (GOI 2013). However, rice cultivation is still continued by many farmers and recently promoted by programmes to cultivate and conserve traditional rice varieties (Manoj 2012) and by prohibition of conversion of rice land for other purposes (Government of Kerala 2008).
Experimental design
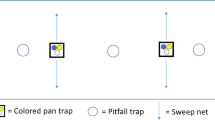
In total, 18 rice fields were selected for the study (Fig. S1). To incorporate cultivation practices and landscape structure, we selected rice fields with high-intensity and low-intensity management adjacent to homegardens (diverse polycultures) and banana fields (intensified monocultures). Samples were taken in 12 subplots per field. Each was 2 × 1 m in size and 2 m apart from each other. Four subplots each were located within three transects to cover the different microhabitats constituting a rice field: (1) at the edge of the field, closest to the adjacent habitat, (2) 10 m to the field and (3) at the earthen bunds in the midst of the rice fields (Fig. 1). The distance from the centre transect to the bund transects varied between 5 and 10 m, depending on the field size.

Rice fields beside homegarden polyculture (top left) and banana monoculture (top right) and sketch of the experimental design. In total 18 rice fields with high-intensity and low-intensity management were selected. Fields were located either adjacent to homegarden polycultures or banana monocultures. Samples were taken in four subplots (each 2 × 1 m, 2 m apart) within three transects (edge, centre, bund) in each field
To differentiate between low-intensity and high-intensity fields, we interviewed the farmers about their cultivation practices during the field site selection. Additionally, we regularly recorded their management steps during the whole sample period, as farmers were flexible in the management practices they applied. We focused on the three major practices of intensification: (1) the amount of applied fertiliser (min = 35.31 kg/ha; max = 705.98 kg/ha), (2) whether insecticides were applied and (3) whether weeding was done or not. No herbicides were used in the study region and weed control was done manually. Permission to work in their rice fields was provided by the farmers in the form of a written agreement.
Data collection
The survey took place during the cropping season after the South-West Monsoon, between August and December in 2011 and 2012. Samples were taken approximately once a month; one each during the tillering stage, during panicle development, flowering and milk grain stage to cover the different phenological stages of the rice crop (Reissig et al. 1986). In 2011, during the milk grain stage, most of the farmers did not permit sampling. Thus, only three sample rounds could be taken. Samples were taken on predominantly dry days between 8 am and 2 pm in an alternating order, so that each field was sampled in the morning and in the early afternoon as well. We collected spiders and insects by sweep netting using a triangular web with a side length of 35 cm and a rounded front. We conducted five sweeps per sampling in 2011, covering the 2 × 1 m of each subplot. Since the number of specimens which were collected by five sweeps was not as high as we had expected, the number of sweeps per subplot and sampling was increased to 15 in the following year. Sweep netting was always done by LB. Before the specimens were transferred into collecting bottles, they were killed. For several minutes we kept the net, with the living animals inside, in a collection bottle (1 l) containing a cotton ball with ethyl acetate. Spiders were then transferred to collecting bottles by hand. This allowed us to take photographs of the specimens without changes of colour or deformations caused by alcohol. Fallowing removal of spiders, the insects were transferred into bottles filled with 70% isopropyl alcohol. Samples were taken to the lab for identification. All specimens were preserved in 70% isopropyl alcohol and stored in the collection of the Zoological Survey of India, WGRC Calicut. Identification of spiders was based on Tikader (1987), Barrion and Litsinger (1995), Murphy and Murphy (2000), Proszynski (2003), Jocqué and Dippenaar-Schoeman (2007), Sebastian and Peter (2009) and the taxonomy followed the World Spider Catalog (2014). Leafhopper identification was based on the descriptions given by Kalshoven (1981) and names were crosschecked with the Encyclopedia of Life (EOL 2014).
Spider webs were surveyed in 2012 during the flowering and milk grain stages. Before the sweep netting was done, each subplot had been carefully searched for webs. A spray bottle filled with water was used to increase the webs’ visibility. We did not step into the subplots while surveying the webs. The rice leaves were moved as little as possible in order to prevent any disturbance of the spiders and the destruction of their webs. We counted and identified the webs. They were categorised into four sizes: tiny (5–10 cm diameter), small (10–15 cm), medium (15–20 cm) and large (≥20 cm). Their orientation (horizontal, vertical and diagonal) was noted as well as their location (top, middle, at base of the rice tillers, and on the ground/soil). These parameters are closely linked to the web types described by Jocqué and Dippenaar-Schoeman (2007) and Sebastian and Peter (2009), which were used for web identification.
A plant survey to assess weed richness and cover in the rice fields was conducted in 2012 during panicle development and milk grain stage. In each subplot the plant species were identified and the coverage for each species was estimated. Specimens that could not be identified in the field were collected and taken to the lab. Sasidharan (2011) and The Plant List (2013) were used as a reference of weed identification and recent taxonomy.
Furthermore, the current land-use in the surroundings of each plot was mapped within a 500 m radius based on Google Earth pictures (scale = 1.7:10,000), verified by a field survey and digitised to a GIS map using ArcGIS 10 (ESRI 2011). We selected a 500 m radius, which seemed to capture the landscape size relevant to spider dispersal (e.g. Schmidt and Tscharntke 2005). We used the Shannon Index (diversity function in the vegan package in R) to calculate the habitat diversity of each landscape, based on the number of different habitat types and the percentage of each habitat type of the landscape’s total area (Steffan-Dewenter et al. 2002).
Statistical analysis
We pooled the data per transect over the 2 years to increase the sample size. For the analysis of spider webs only data from 2012 were used, also pooled per transect. We conducted a multivariate analysis using the mvabund package (3.9.3) (Wang et al. 2016) using R (3.1.2) (R Core Team 2014) to analyse the response of different spider families and spider web types to several environmental and management factors. Separate models were computed for spider families and spider webs. The mvabund package builds on a model-based approach for the analysis of multivariate abundance data. The function manyglm of the mvabund package computes generalised linear models for each species (or, as in our case, families and web types respectively) in a dataset separately using a collective set of explanatory variables. Using the function anova.manyglm(), adjusted for multiple testing, p.uni = “adjusted”, multivariate and univariate results for each family were obtained. We used a negative binomial distribution to account for count data. An integrated inference tool in the model takes correlations between families into account (Wang et al. 2012). Checks for homoscedasticity, normality of errors and absence of outliers were done by diagnostic plots of each model. Overlapping landscapes (as it was the case in several locations due to a restricted number of suitable fields) does not necessarily lead to spatial autocorrelation (Zuckerberg et al. 2012). Spatial autocorrelation should only be taken into account in the model if it occurs. However, there is no option to check for spatial autocorrelation in manyglm residuals so far. A Spearman’s correlation test was conducted to check for correlations between the explanatory variables, namely adjacent habitats (homegarden or banana), the amount of applied fertiliser (kg/ha), insecticide application, weed operations, landscape diversity, abundance of leafhoppers, Lepidoptera and other insects, weed richness, percentage of weed cover and the percentages of landscape components (homegarden, banana, rice and fallow) in the landscape (Table S1). Due to correlations between landscape structure and percentage of homegarden, leafhopper abundance and abundance of Lepidoptera and other insects respectively, we fitted the following models:
-
(1)
y ~ adjacent habitat + amount of applied fertiliser + insecticide application + weed operations + landscape structure + abundance of all insects + weed richness + percentage of weed cover
-
(2)
y ~ % homegarden + % banana + % rice + % fallow
-
(3)
y ~ leafhopper abundance
-
(4)
y ~ Lepidoptera abundance
-
(5)
y ~ abundance of other insects.
Additionally, we conducted a Spearman’s correlation test between the abundances of leafhoppers, Lepidoptera, other insects and the application of fertilisers to check whether there is an indirect effect of fertiliser through prey abundance or not.
Results
In total, we collected 2073 spider individuals belonging to 15 families and 86 species. Only the seven most abundant spider families were included in the analysis (total abundance ≥40), i.e. Tetragnathidae, Oxyopidae, Salticidae, Araneidae, Lycosidae, Linyphiidae, Thomisidae (Table 1, Table S2). For the other families we had too few counts to obtain meaningful results. A total of 386 spider webs were counted and classified into five different web types, namely orb webs, space webs, ground webs, tetragnathid webs and reduced webs (Table 2). Reduced webs were excluded from the analysis because of insufficient records.
A total of 15,620 leafhoppers were counted over the 2 years with 10,367 individuals collected only in 2012. These individuals were identified to species level if possible. Leafhoppers were dominated by well known rice pest species, namely Sogatella furcifera (n = 2103), Nephotettix spp. (n = 1899) and Recilia dorsalis (n = 573) (Table S3). Furthermore, 7116 insects of ten orders other than leafhoppers were recorded, i.e. Coleoptera, Dermaptera, Hemiptera, Hymenoptera, Isoptera, Lepidoptera, Mantodea, Odonata, Orthoptera and Planipennia (Table S4). Among the 1396 Lepidoptera (adult and larvae) observed, we recorded high numbers of Nymphula depunctalis (rice leaf-roller) and Cnaphalocrocis medinalis (leaf-folder) (together constituting about 70% of the 2012 samples), both are known as rice pests.
The results of the multivariate analysis (Table 3, Supporting Information III) showed that adjacent habitat and landscape structure did not affect spider families or spider web types. Moreover, no effect could be recognised for the three different management practices, fertiliser and insecticide application and weeding operations. Weed richness did not affect spider families or web types. Also, the percentage of different landscape components had no effect on spider families. However, ground webs were positively related to the percentage of fallows.
The number of available insect prey explained a significant amount of variation in spider abundance and spider web number (Fig. 2; Table S5). A closer look at these numbers revealed different responses of spider families and web types to the abundance of leafhoppers, Lepidoptera and other insects. As the results of the univariate analysis have shown, all spider families were positively related to Lepidoptera abundance, while only Tetragnathidae, Oxyopidae and Linyphiidae positively responded to leafhopper abundance. The number of Tetragnathidae, Oxyopidae and Salticidae significantly increased with raising numbers of other insects (Fig. 3; Tables S7, S8 and S9). Furthermore, the increase in herb cover was negatively related to spider families, but this result was only significant for Oxyopidae (Fig. 2; Table S5).

Response of spider families to a percent of herb cover in the rice fields and b insect abundance. Significance codes: ***≤0.001, **≤0.01, *≤0.05, ·≤0.1. Ara = Araneidae, liny = Linyphiidae, lyco = Lycosidae, oxy = Oxyopidae, tetra = Tetragnathidae, thom = Thomisidae

Relationship between spider families and a leafhopper abundance, b Lepidoptera abundance and c other insects except leafhoppers and Lepidoptera. Significance codes: ***≤0.001, **≤0.01, *≤0.05, ·≤0.1. Ara = Araneidae, liny = Linyphiidae, lyco = Lycosidae, oxy = Oxyopidae, tetra = Tetragnathidae, thom = Thomisidae
The analysis of spider webs showed similar results as the analysis of spider families—the abundance of insect prey being the most important (Fig. 4a; Table S10). The number of Tetraganthid and orb webs increased with numbers of leafhoppers, Lepidoptera and other insects (Fig. 5; Table S12, S13 and S14). Moreover, spider web numbers were positively related to higher percentages of fallow fields in the surrounding landscape (Fig. 4b; Table S11) while herb cover had no effect. Fertiliser applications were not related to leafhoppers, Lepidoptera and other insects (Fig. S1).

Relationship between spider web types and a insect abundance and b percentage of fallow field in the surrounding landscape. Significance codes: ***≤0.001, **≤0.01, *≤0.05, ·≤0.1. Ground = ground webs, orb = orb webs, space = space webs, tetra = Tetragnathidae-webs

Relationship between spider web types and increasing a leafhopper abundance, b Lepidoptera leafhopper and c other insects except leafhoppers and Lepidoptera. Significance codes: ***≤0.001, **≤0.01, *≤0.05, ·≤ 0.1. Ground = ground webs, orb = orb webs, space = space webs, tetra = Tetragnathidae-webs
Discussion
Leafhoppers, Lepidoptera and other insect prey
The availability of prey explained most of the density of the seven most abundant spider families (>40 individuals) and four spider web types. However, in contrast to, e.g., Nyffeler and Benz (1987), Kiritani et al. (1972), Sebastian et al. (2005), Takada et al. (2012), Takada et al. (2013), Lou et al. (2013), who describe a top-down effect of spiders on pest insects, our findings suggest that spiders were driven by bottom-up effects. Marc et al. (1999) and Harwood et al. (2001) describe the aggregation of spiders in prey rich areas as a possible numerical response. Furthermore, prey switching can occur, when spiders change their prey preferences in favour of the most abundant prey (Nyffeler et al. 1994a; Riechert and Lawrence 1997). In the studied case, spiders follow the density of their prey. This indicates that spiders may have the potential to prevent severe pest outbreaks or even to control pest insects in the rice fields. However, to evaluate pest regulation delivery by spiders in the studied area, further investigations are necessary.
The number of leafhoppers and Lepidoptera was most important for the most abundant spider family, Tetragnathidae. Abundance of other insects also had a positive effect, but it was less significant. In case of tetragnathid webs, the response to leafhopper abundance was least significant, compared to Lepidoptera and other insects. Tetragnathidae build large, horizontal webs in the upper part of the rice vegetation or in its canopy. According to Tahir et al. (2009) the order of prey caught by Tetragnatha javana is Lepidoptera, followed by Diptera, Homoptera, Coleoptera, Hymenoptera and Orthoptera, which corresponds with our findings, showing highest increase of Tetragnathidae with increasing abundance of Lepidoptera and leafhoppers. Kiritani et al. (1972) also showed that Tetragnathidae prey efficiently on leafhoppers.
For Araneidae, only the increase of Lepidoptera abundance showed a slightly significant effect. However, the numbers of orb webs, which are primarily build by Araneidae, not only increased significantly with the numbers of Lepidoptera, but also with leafhoppers and other insects. Tahir et al. (2009) found that Lepidoptera were the preferred prey of the Araneidae Neoscona theisi, which corresponds with our findings for Araneidae and orb webs.
The low numbers of Linyphiidae in this study might be partly due to a sample bias. Linyphiidae are usually found at the base of the rice tillers where it was difficult to reach with the sweep net in tall rice varieties (up to 1.4 m high). However, Sebastian et al. (2005) recorded similarly low numbers of Linyphiidae. Nevertheless, the results showed a slightly positive relationship between Linyphiidae and Lepidoptera and leafhopper abundance respectively. These findings are in line with Lou et al. (2013), who reported that Linyphiidae do feed on leafhoppers but are more important predators of young Lepidoptera larvae and rice aphids. More than 60% of the captured Lepidoptera in our study were larvae, which may explain the positive relationship between Lepidoptera and Linyphiidae. The lack of rice aphids in the studied fields could be a reason why the abundance of other insects did not have a significant effect.
The second most abundant spider family, Oxyopidae, hunts by chasing or stalking. The numbers of Oxyopidae increased with the abundance of leafhoppers, Lepidoptera and other insects. Tahir and Butt (2009) showed in their experiment that Oxyopes javanus do feed on leafhoppers, Lepidoptera and grasshopper nymphs, but preferred Lepidoptera larvae over leafhopper nymphs and adults. Their least favourite prey were grasshopper nymphs. Noticeable were the high numbers of immature Oxyopes sp. individuals in our samples. However, Barrion et al. (2012) found such high number of immature Oxyopes sp. in rice fields in China as well and reported their efficiency in feeding on nymphs of leafhoppers. Although much smaller than the adults but high in numbers immature Oxyopes sp. may have the potential to reduce the number of leafhopper nymphs.
The number of Salticidae, also hunting spiders, increased with higher numbers of Lepidoptera and other insects. The frequently observed moths N. depuntalis and C. medinalis fit well into the prey spectrum of Carrhotus viduus (C. L. Koch) Sebastian and Peter (2009), which was the most abundant member of the Salticidae in our study fields. Additionally, Salticidae may prey on the larvae of N. depunctalis and C. medinalis, which were mostly observed in the middle layer of the rice vegetation. This is the preferred hunting ground of these spiders. Furthermore, Salticidae may also hunt less airworthy insects, such as beetles (Riechert and Bishop 1990; Nyffeler et al. 1994b), which could explain their positive response to other insects.
The numbers of Lycosidae found in this study was comparatively low, probably, because sweep netting is not the most efficient way to sample this group. Pitfall traps would have been much more appropriate to capture these ground running spiders but due to the standing water in the fields, they could not be installed. In contrast to studies by Kiritani et al. (1972), Nyffeler and Benz (1987) and Lou et al. (2013), who reported leafhoppers to be the preferred prey of Lycosidae, our results indicated an increase in Lycosidae abundance with higher numbers of Lepidoptera. However, the analysis showed a slightly positive response to leafhopper abundance as well, though it was not significant.
In the study area damage to the crop due to pest species do occur, but only occasionally and, according to the farmers, very locally. However, as the majority of the rice farmers in Wayanad are subsistence farmers, these locally restricted outbreaks can cause significant crop damage and substantially decrease an individual farmer’s yield. In consequence, those affected are forced to buy additional rice from the market to supply the need of rice for the year, which often results in serious financial hardship. Therefore, conserving the spider community in the rice fields that is potentially able to control pests and prevent pest outbreaks, is a useful measure for these farmers.
Weed cover and richness
Structural diversity provided by plant diversity was hypothesised to be important to conserve spider populations in agricultural fields. Plant diversity provides hiding places for hunting spiders and diverse options for web builders to fix their webs and also additional prey insects, which are related to non-crop plants (Balfour and Rypstra 1998; Sunderland and Samu 2000; Foelix 2011). However, we did not find any effects of plant diversity, on spider families or on spider webs. Moreover, our results indicated a trend towards a decline in abundance for all spider families with an increase of herb cover in the rice fields. However, only the effect on Oxyopidae was significant. Probably a too dense vegetation impedes foraging of these free hunting spiders and thus the prey have more options to hide (Butt and Xaaceph 2015). Web builders need enough structure to fix their webs but also require enough space to build them properly (Foelix 2011). Perhaps, denser herb vegetation in rice fields reduces the spaces for web building or the chance that prey is intercepted by spider webs. Therefore, mechanical weeding by hand, as it is done in the study area, apparently does not disturb spider population in the fields. Furthermore, it might also be possible that the efficiency of invertebrate capture is reduced in fields with denser weed cover.
Adjacent habitat, landscape structure and management
In contrast to the findings of Clough et al. (2005) and Schmidt et al. (2008), no effects were found for the type of adjacent habitat (homegarden polyculture vs. banana monoculture), but the percentage of fallow fields in the surrounding landscape showed a positive impact on the number of ground webs. Fallows serve as a retreat during the dry season for some spiders, especially for those living in small crevices in the soil and on the bunds (Arida and Heong 1994; Bambaradeniya et al. 2004). Thus, fallow fields within the rice landscape may help in conserving ground dwelling spiders in the rice fields. Homegarden polycultures and banana monocultures are dominated by woody plants and may possibly be inhabited by a spider assemblage that prefers woody habitats and does not spill-over into open habitats, such as the rice fields (Entling et al. 2007). Lemessa et al. (2015) and Schmidt et al. (2008) discussed that the spatial scale showing explanatory power is species specific. Thus, a 500 m radius is perhaps not the best spatial scale for all species and a possible reason for the low impact of landscape patterns found. Hence, the type of adjacent habitat and landscape structure seemed to be of less importance for the conservation of the spider families in rice fields compared to the findings of studies in different contexts (e.g. Marc et al. 1999; Schmidt et al. 2008; Schoenly et al. 2010).
Several studies showed a harmful effect of chemical insecticides and fertilisers on spiders (e.g. Marc et al. 1999; Settle et al. 1996; Amano et al. 2011). However, we did not find any effect of different levels of fertiliser or of insecticide applications. Similarly, insect abundance was not affected by fertiliser application, suggesting that there is no indirect effect through prey abundance. The reason for the lack of impact of the different management practices could be that intensification of rice cultivation in the studied region had not yet reached levels high enough to significantly damage the spider population. Compared to other rice cultivation regions, the intensification observed in Wayanad is very modest (Lu and Heong 2009). The interviews and discussions with the farmers showed that the opinions about how to cultivate rice are diverse and do range from the traditional way only using organic manure and manual pest management to very intensified management with high yielding varieties, chemical fertiliser and pesticides, larger fields, irrigation and machineries and all possible combinations in between. However, there also are those farmers (usually owning larger portions of land) who totally believe in intensified agriculture and who are trying to promote it within the district. Hence, where the future of Wayanad’s rice cultivation lies is hard to predict. It will depend on various factors such as the farmers’ personal decisions, the market structure, local politics, etc.
Spider web sampling
The identification of spider webs between the rice plants was not easy and perhaps is less precise, compared with studies by Gollan et al. (2010) and Stenchly et al. (2011), which were conducted in woody habitats. However, spider web sampling can complement the spider sampling. In fields where the installation of pitfall traps was impossible, the survey of spider webs might be a good addition to sweep netting, as it is rather difficult to catch spiders at the base of the rice tillers, especially in later cropping stages when the crop has grown high. For instance, Erigoninae (Linyphiidae), which build ground webs—tiny sheet webs, built over small holes in the soil, might be missed by sweep netting but could be detected by their webs. The same may apply for Araneidae, which hide in the middle layer of the vegetation. Although spider webs and spider abundances have been sampled in different years, the analyses suggest diverging results. For instance, the analysis of spider families only showed an increase of Araneidae with an increasing number of Lepidoptera, while the analysis of spider webs indicated that orb webs, which are mainly build by Araneidae, responded positively to increasing numbers of leafhoppers and also other insects. Ground webs were positively related to higher percentages of fallows in the surrounding, but such a relationship was not found for Linyphiidae.
In addition, the large horizontal webs in the rice canopy built by Tetragnathidae are easy to observe in the early morning when they are covered with dew drops. High numbers of tetragnathid webs could indicate to the farmers that insect pests are potentially being controlled by spiders, suggesting deciding against pesticide application. Interviews during the field site selection revealed that several farmers have a good understanding about the complexity of the rice agroecosystem and also explained the role of spiders within the fields; that they feed on the pest species and therefore are helpful. Moreover, some farmers, who have started with intensified management because they had hoped for higher yields and more security against pest outbreaks, were frustrated because these expectations did not become true. Pesticides had not shown an effect but the farmers simultaneously noticed a drop in the quality of the rice. Therefore, they asked us, and discussed among themselves during a workshop, about alternatives. Those farmers who did not use pesticides often followed the traditional way of pest control using Artemisia plants as pest repellents. Many farmers were interested in the concept of biological pest control and asked about how to promote it in practice, which might be a good sign towards sustainable and environmentally friendly smallholder rice cultivation.
Conclusion
In conclusion, the findings of this study showed that the major determining factor for overall spider and spider web abundance was the prey availability, indicating that the spider community in these rice fields was driven by bottom-up effects. This suggests that spiders may have the potential to control the number of pest insects in the rice fields. Hence, conservation biological control of spiders could be a measure to prevent pest outbreaks and thus losses in harvest as well as the environmentally problematic application of pesticides. The spider assemblage in rice fields and the adjacent, woody habitats (homegarden polyculture vs. banana monoculture) may be too different, thereby hindering important spill-over across habitat edges. However, maintaining fallow fields in the surrounding landscape of the rice fields appeared to be a useful measure to conserve predators in the rice fields, as fallows can serve as refuge habitats during the dry season for spiders such as Erigoninae. The missing evidence that fertiliser and insecticide application had an impact on spiders may be due to the low level of intensification. Herbicides were not used in the study region and the weeding by hand apparently does not disturb spider populations. Spider web sampling can be a useful complement to spider sampling. Spiders that occur in the lower levels of the rice vegetation or at the base of the tillers might be missed by sweep netting, but are likely to be detected by their webs. In addition, the large tetragnathid webs are easy to observe and indicate high numbers of Tetragnathidae, which could be a reason for farmers not to spray pesticides as spiders are potentially already controlling the number of pest insects. Although there are farmers in the study area who want to push the intensification of rice cultivation forward, there also are many farmers, who see the drawbacks of an intensified agriculture and either already follow organic farming or are very interested in how to apply environmentally friendly management methods.
References
Amano T, Kusumoto Y, Okamura H, Baba YG, Hamasaki K, Tanaka K, Yamamoto S (2011) A macro-scale perspective on within-farm management: how climate and topography alter the effect of farming practices. Ecol Lett 14:1263–1272
Arida GS, Heong KL (1994) Sampling spiders during the rice fallow period. Int Rice Res Notes 19:20
Balfour RA, Rypstra AL (1998) The influence of habitat structure on spider denisty in a no-till soybean agroecosystem. J Arachnol 26:221–226
Bambaradeniya C, Edirisinghe JP, deSilva DN, Gunatilleke CVS, Ranawana KB, Wijekoon S (2004) Biodiversity associated with an irrigated rice agro-ecosystem in Sri Lanka. Biodivers Conserv 13:1715–1753
Barrion AT, Litsinger JA (1995) Riceland spiders of South and Southeast Asia. CAB International, Wallingford
Barrion AT, Villareal SS, Catindig J, Cai D, Yuan QH, Heong KL (2012) The spider fauna in the rice agricultural landscape of Hainan Island, China: composition, abundnace and feeding structure. Asian Life Sci 21:625–651
Bianchi FJJA, Booij CJH, Tscharntke T (2006) Sustainable pest regulation in agricultural landscapes: a review on landscape composition, biodiversity and natural pest control. Proc R Soc B 273:1715–1727
Butt A, Xaaceph M (2015) Functional Response of Oxyopes javanus (Araneidae: Oxyopidae) to Sogatella furcifera (Hemiptera: Delphacidae) in Laboratory and Mesocosm. Pak J Zool 47:89–95
Clough Y, Kruess A, Kleijn D, Tscharntke T (2005) Spider diversity in cereal fields: comparing factors at local, landscape and regional scales. J Biogeogr 32:2007–2014
Entling W, Schmidt MH, Bacher S, Brandl R, Nentwig W (2007) Niche properties of Central European spiders: shading, moisture and the evolution of the habitat niche. Global Ecol Biogeogr 16:440–448
EOL (2014) Encyclopedia of Life. http://eol.org/. Accessed 15 Sep 2014
Foelix RF (2011) Biology of spiders, 3rd edn. Oxford University Press, Oxford, New York
George J, Krishnaprasad P (2006) Agrarian distress and farmers’ suicide in the tribal Distric of Wayanad. Social Sci 34:70–85
GOI (2013) Facts and Figures of Agriculture in Kerala. Department of Agriculture, Thiruvananthapuram, Kerala. http://www.keralaagriculture.gov.in/pdf/fact_figures_25062014.pdf. Accessed 16 Sep 2014
Gollan J, Smith H, Bulbert M, Donnelly A, Wilkie L (2010) Using spider web types as a substitute for assessing web-building spider biodiversity and the success of habitat restoration. Biodivers Conserv 19:3141–3155
Government of Kerala (2008) Kerala Conservation of Paddy Land and Wetland Act, 2008
Harwood JD, Sunderland KD, Symondson W (2001) Living where the food is: web location by linyphiid spiders in relation to prey availability in winter wheat. J Appl Ecol 38:88–99
Horgan FG, Crisol E (2013) Hybrid rice and insect herbivores in Asia. Entomol Exp Appl 148: 1–19
Jocqué R, Dippenaar-Schoeman AS (2007) Spider families of the world, 2nd edn. Musée royal de l’Afrique centrale; ARC-PPRI, Tervuren
Kalshoven LGE (1981) Pests of crops in Indonesia. P. T. Ichtiar Baru - Van Hoeve, Jakarta
Kiritani K, Kawahara S, Sasaba T, Nakasuji F (1972) Quantitative evaluation of predation by spiders on the green rice leafhopper, Nephotettix cincticeps Uhler, by a sight-count method. Res Popul Ecol 13:187–200
Kumar NA, Gopi G, Prajeesh P (2010) Genetic erosion and degradation of ecosystem services of wetland rice fields: a case study from Western Ghats, India. In: Lockie S, Carpenter D (eds) Agriculture, biodiversity and markets. Earthscan, London, pp 137–153
Landis DA, Wratten SD, Gurr GM (2000) Habitat management to conserve natural enemies of arthropod pests in agriculture. Annu Rev Entomol 45:175–201
Laurance WF (2010) Habitat destruction: death by a thousand cuts. In: Sodhi NS, Ehrlich PR (eds) Conservation biology for all. Oxford University Press, Oxford, pp 73–87
Lee SY, Kim ST, Jung JK, Lee J-H (2014) A comparison of spider communities in Bt and non-Bt rice fields. Environ Entomol 43: 819–827
Lemessa D, Hambäck PA, Hylander K (2015) The effect of local and landscape level land-use composition on predatory arthropods in a tropical agricultural landscape. Landscape Ecol 30:167–180
Lou Y-G, Zhang G-R, Zhang W-Q, Hu Y, Zhang J (2013) Biological control of rice insect pests in China. Biol Control 67:8–20
Lu Z, Heong KL (2009) Effects if nigtrogen-enriched rice plants on ecological fitness of planthoppers. In: Heong KL, Hardy B (eds) Planthoppers: new threats to the sustainability of intensive rice production systems in Asia. International Rice Research Institute (IRRI), Los Baños, pp 247–266
Lu Z, Zhu P, Gurr GM, Zheng X, Chen G, Heong KL (2015) Rice pest management by ecological engineering: a pioneering attemt in China. In: Heong KL, Cheng J, Escalada MM (eds) Rice planthoppers: ecology, management, socio economics and policy. Springer Science + Business Media, Dordrecht, pp 163–180
Maloney D, Drummond FA, Alford R (2003) Spider predation in aroecosystems: Can spiders effectively control pest populations? Department of Biological Sciences, University of Maine, Orono
Manoj EM (2012) Lifeline for Wayanad’s traditional rice varieties. The Hindu, August 27. http://www.thehindu.com/todays-paper/tp-national/tp-kerala/article3826145.ece#.UDsRyyGDRx0.email. Accessed 23 May 2015
Marc P, Canard A, Ysnel F (1999) Spiders (Araneae) useful for pest limitation and bioindication. Agric Ecosyst Environ 74:229–273
Matson PA (1997) Agricultural intensification and ecosystem properties. Science 277:504–509
Murphy F, Murphy JA (2000) An introduction to the spiders of South East Asia. With notes on all the genera. Malaysian Nature Society, Kuala Lumpur
Nyffeler M, Benz G (1987) Spiders in natural pest control: a review. J Appl Enthomol 103:321–339
Nyffeler M, Sterling WL, Dean DA (1994a) Insectivorous activities of spiders in United States field crops. J Appl Entomol 118:113–128
Nyffeler M, Sterling WL, Dean DA (1994b) How spiders make a living. Environ Entomol 23:1357–1367
Pandey S, Byerlee D, Dawe D, Dobermann A, Mohanty S, Rozelle S, Hardy B (eds) (2010) Rice in the global economy. Strategic research and policy issues for food security. International Rice Research Institute, Manila
Proszynski J (2003) Salticidae (Aranae) of the World. http://salticidae.org/salticid/diagnost/title-pg.htm. Accessed 18 May 2015
R Core Team (2014) R: a language and environment for statistical. R Foundation for Statistical Computing, Vienna
Reissig WH, Heinrichs EA, Litsinger JA, Moody K, Fiedler L, Mew TW, Barrion AT (1986) Illustrated guide to integrated pest management in rice in tropical Asia. International Rice Research Institute, Manila
Riechert SE, Bishop L (1990) Prey control by an assemblage of generalist predators: spiders in garden test systems. Ecology 71:1441–1450
Riechert SE, Lawrence K (1997) Test for predation effects of single versus multiple species of generalist predators: spiders and their insect prey. Entomol Exp Appl 84:147–155
Santhoshkumar A, Ichikawa K (2010) Homegardens: sustainable land use systems in Wayanad, Kerala, India. In: Bélair C, Ichikawa K, Wong BYL, Mulongoy KJ (eds) Sustainable use of biological diversity in socio-ecological production landscapes. Background to the ‘Satoyama Initiative for the benefit og biodiversity and human well-being’. Secretariat of the Convention on Biological Diversity, Montreal, pp 125–128
Sasidharan N (2011) Flowering plants of Kerala. 2nd edn. Kerala Forest Research Institute, Peechi, Kerala
Schmidt MH, Tscharntke T (2005) Landscape context of sheetweb spider (Araneae: Linyohiidae) abundance in cereal fields. J Biogeogr 32:467–473
Schmidt MH, Thies C, Nentwig W, Tscharntke T (2008) Contrasting responses of arable spiders to the landscape matrix at different spatial scales. J Biogeogr 35:157–166
Schoenly KG, Cohen JE, Heong KL, Litsinger JA, Barrion AT, Arida GS (2010) Fallowing did not disrupt invertebrate fauna in Philippine low-pesticide irrigated rice fields. J Appl Ecol 47:593–602
Sebastian PA, Peter KV (eds) (2009) Spiders of India. Universities Press, Hyderabad
Sebastian PA, Mathew MJ, PathummalBeevi S, Joseph J, Biju CR (2005) The spider fauna of irrigated rice ecosystem in central Kerala, India across different elevational ranges. J Arachnol 33:247–255
Settle WH, Ariawan H, Astuti ET, Cahyana W, Hakim AL, Hindayana D et al (1996) Managing tropical rice pests through conservation of generalist natural enemies and alternative prey. Ecology 77:1975–1988
Steffan-Dewenter I, Münzenberg U, Bürger C, Thies C, Tscharntke T (2002) Scale-depentent effects of landscape context on three pollinator guilds. Ecology 83:1421–1432
Stenchly K, Clough Y, Buchori D, Tscharntke T (2011) Spider web guilds in cacao agroforestry—comparing tree, plot and landscape-scale management. Divers Distrib 17:748–756
Stenchly K, Clough Y, Tscharntke T (2012) Spider species richness in cocoa agroforestry systems, comparing vertical strata, local management and distance to forest. Agric Ecosyst Environ 149:189–194
Sunderland K, Samu F (2000) Effects of agricultural diversification on the abundance, distribution, and pest control potential of spiders: a review. Entomol Exp Appl 95:1–13
Tahir HM, Butt A (2009) Predatory potential of three hunting spiders inhabiting the rice ecosystems. J Pest Sci 82:217–225
Tahir HM, Butt A, Sherawat SM (2009) Foraging strategies and diet composition of two orb web spiders in rice ecosystems. J Arachnol 37:357–362
Takada MB, Yoshioka A, Takagi S, Iwabuchi S, Washitani I (2012) Multiple spatial scale factors affecting mirid bug abundance and damage level in organic rice paddies. Biol Control 60:169–174
Takada MB, Kobayashi T, Yoshioka A, Takagi S, Washitani I (2013) Facilitation of ground-dwelling wolf spider predation on mirid bugs by horizontal webs built by Tetragnatha spiders in organic paddy fields. J Arachnol 41:31–35
The Plant List (2013) Version 1.1. http://www.theplantlist.org/. Accessed 30 Sep 2014
Tikader BK (1987) Handbook of Indian spiders. Zoological Survey of India, Calcutta
Tilman D, Fargione J, Wolff B, D’Antonio C, Dobson A, Howarth R et al (2001) Forecasting agriculturally driven global environmental change. Science 292:281–284
Uetz G, Halaj J, Cady AB (1999) Guild structure of spiders in major crops. J Arachnol 27:270–280
Wang Y, Naumann U, Wright ST, Warton DI (2012) mvabund—an R package for model-based analysis of multivariate abundance data. Methods Ecol Evol 3:471–474
Wang Y, Naumann U, Wright S, Eddelbuettel D, Warton D (2016) mvabund: statistical methods for analysing multivariate abundance data. R package version 3.11.9. https://CRAN.Rproject.org/package=mvabund
World Spider Catalog (2014) World Spider Catalog, Version 15.5. http://wsc.nmbe.ch. Accessed 15 Sep 2014
Zhao Z, Sandhu HS, Gao F, He D (2015) Shifts in natural enemy assemblages resulting from landscape simplification account for biocontrol loss in wheat fields. Ecol Res 30:493–498
Zuckerberg B, Desrochers A, Hochachka WM, Fink D, Koenig WD, Dickinson JL (2012) A persistent, but misdirected concern when collecting and analyzing ecological data. J Wildl Manag 76:1072–1080
Acknowledgements
We are grateful for the cooperation of 15 farmers of Wayanad: Achappan Palookkappu, Bharathan Kalluvayal, Gopalan Kalluvayal, Gopalan Chempakappatta, Hareendran Cherukkattoor, Jhon Paliyana, Manoj Paliyana, Mathai Paliyana, Radhakrishnan Paliyana, Rajan Paliyana, Raman Palliyara, Sudheesh Karingari, Sunil Karingari, Vijayan Thamarachira and Yamuna Paliyana. We are grateful to Parameshwaran Prajeesh who conducted the plant survey. We would like to thank Sabitha, Mathews, Kamarudheen, and Prashoob for field assistance, Dr. Sunil Jose for assistance with spider identification and Bastian Niemeyer who digitised the land-use maps. We duely acknowledge the support provided by the BioDIVA project and colleagues and the M.S. Swaminathan Research Foundation. We thank Stephan Rudolph, Arne Schulze and two anonymous reviewers for helpful comments on an earlier version of this manuscript. This study was conducted whithin the BioDIVA research group, funded by FONA—Social-Ecological Research, BMBF (Federal Ministry of Education and Research, Germany).
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Betz, L., Tscharntke, T. Enhancing spider families and spider webs in Indian rice fields for conservation biological control, considering local and landscape management. J Insect Conserv 21, 495–508 (2017). https://doi.org/10.1007/s10841-017-9990-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10841-017-9990-2