
Abstract
Abiotic disturbances often totally change the spatial arrangement of the forest environment and potentially reduce the proceeds from the forest. Nature conservationists face considerable pressure to log the wood, or at least to reduce the risk of forthcoming insect outbreaks. The aim of this study was to determine which environmental factors were the most significant for the species composition of subcorticoulous saproxylic beetles in post-disturbance areas. Beetles were studied in Primorsky Region woodlands (Far Eastern Russia) in 2008, in the second year following a heavy snow-break. The composition of saproxylic beetles was influenced significantly, and nearly in the same way, by microhabitat and tree level characteristics. The effect of interaction between microhabitat and tree level was low. Discrimination between coniferous and deciduous trees as the most important factor was followed by the stage of bark peeling, wetness of bast, presence of wood-inhabiting fungi, consistency of bast and sun exposure, all of which were significant predictors. Several taxa showed different demands between adults and larvae (e.g. Cucujus haematodes) or between species within a particular family (e.g. Pyrochroidae). The studied area was definitely not pristine woodland, but the reforestation was non-commercial in nature, with maximum use of natural regeneration and the presence of green trees in disparate life-stages was dominant. The results indicated that natural regeneration with retention of dead wood following a disturbance could be the goal in woodland areas with high conservation interest.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Abiotic disturbances such as wind, snow or ice-breaks, fires or floods often change the spatial arrangements in a forest completely (Svoboda et al. 2012). Moreover, they are often followed by insect outbreaks and, in terms of harvested forests, potential insect pests usually lack sufficient numbers of natural enemies to control them (Wermelinger 2004). It is also an indisputable fact that the afore-mentioned disturbances, whether caused by abiotic or biotic factors, reduce the proceeds from the forest (Carnus et al. 2006). In the case of abiotic disturbances in harvested forests, one risk (especially in Europe) is that the area requiring salvage loggings will expand due to attack by bark beetles (Wermelinger 2004). This can lead to a serious problem, not only in terms of lost productivity and the accompanying financial loss, but also for foresters who often face problems caused by the public—because the public use and access to the forest can be restricted.
In places where harvested forests predominate, the majority of the timber is logged and what would otherwise be a paradise for saproxylic organisms usually disappears within a few weeks or months due to advanced and effective forestry technologies. The only chances for saproxylics are usually in protected or less accessible areas where all or most of the wood is left to natural decomposition (Pouska et al. 2010; Svoboda et al. 2012). From the perspective of the forest manager, this situation often risks subsequent insect outbreaks with hotspots originating in protected areas (Wermelinger 2004; Modlinger et al. 2009). Furthermore, there is often a public outcry to restore (plant-up) such areas as they are perceived to be dead and of little value. Nature conservationists face considerable pressure to log the wood, or at least to reduce the risk of forthcoming outbreaks—e.g. by debarking the logs (Modlinger et al. 2009).
This is the real-world situation in terms of central Europe. On the other hand, disturbance to forest ecosystems typically results in the creation of wider, and more diversified, range of habitats for potential colonisation by biota—such as lying dead trees, snags and canopy openness. Furthermore, diversity in succession of these habitat types is higher when disturbances are repeated (Svoboda et al. 2012). Relatively little is known about the situation in areas where, although there is commercial logging, large areas are left to semi-natural succession (Christiansen and Bakke 1988). Such a situation prevails, for example, in Asia, where a significant portion of the forests were clear-cut in the past and the presence of veteran trees is rather unusual. There is still relatively little data from this area, but some studies suggest it may be, in comparison with Europe, the territory of many threatened and rare species (Horak et al. 2011; Muller et al. 2013).
From the perspective of this paper, the main assumption for the study of such forest ecosystems was that abiotic disturbances will naturally be followed by growing population densities of early arriving insects (Jonasova and Prach 2008). The second assumption was that, due to the semi-natural composition of the stands, early arriving saproxylics will not achieve such high population densities as in Europe (Wermelinger 2004). In this study, because the abiotic disturbance was caused by snow-breaks, in the growing season, and to broadleaved species, there was an assumption that altitude (broadleaves are replaced by conifers towards higher altitudes in the study area) could have a significant impact on the species composition of saproxylic beetles (Amman 1973).
The main aims of this study were to establish how the species composition of saproxylic beetles in a post-disturbance area of Primorsk (Far East Russia) was influenced by the spatial distribution of sampling sites, including environmental factors at the tree and microhabitat levels and also by interactions between them. An accompanying interest was in the effect of particular predictors and their influence on individual saproxylic beetles. Finally, the potential influence of altitude on the composition of saproxylic beetles was studied.
Materials and methods
Area of study
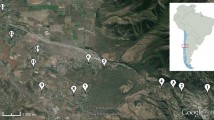
Beetle communities were studied in Far Eastern Russia in 2008 in an area covering more than 30,000 km2—namely, in the southern part of the Primorsky Region between 42.70N and 44.20N and between 131.50E and 135.50E, centered at 43.4506N and 133.4741E. This region borders China and North Korea in the west, the Sea of Japan in the south and east and the Khabarovsky Region (Russia) in the north. The mean altitude of the study area was ≈530 m a.s.l., ranging from 10 to 1,200 m a.s.l. The study region was highly diverse—from the tundra in the Sikhote-Alin Mountains, to coniferous and coniferous-deciduous forests to forest-steppe. However, the transition from boreal to temperate climate forests dominated. Relatively large area of the forests are harvested, although dead wood of damaged trees and also mature trees are often left for succession (Fig. 1). This area was disturbed by a heavy snowfall in the autumn of 2007, which caused considerable damage to still leafy trees in most parts of the study area.

Forestry in Primorsky kray (Far Eastern Russia). a Large clear-cuts with standing dead and snagged trees, high stumps and lying dead trees, living trees are mostly damaged or of low commercial importance; b High accumulations of logs of low commercial importance left after timber hauling; c Diversity of tree species composition as a result of semi-natural succession after large clear-cuts; d An example of heavy snow-break disturbance in forest of semi-natural origin. These forests are home of Cucujus haematodes (Cucujidae), Peltis giganteum (Trogossitidae), Pytho kolwensis (Pythidae) or Boros schneideri (Boridae), saproxylic beetles of high nature conservation importance
Sampling method
Bark peeling was used as one of the most suitable sampling methods for studying subcorticoulous species and communities of saproxylic beetles (Horak et al. 2012). Each sample was taken approximately at the center of the length of a log or at 1.3 m (breast height) in the case of a snagged tree. Equal random-stratified sampling was used to ensure the same sampling intensity (Hedgren and Weslien 2008): an equal number and sample dimension was chosen in each random piece of dead wood (Hirzel and Guisan 2002). Bark samples peeled with a screwdriver had a standardized surface area of 30 × 30 cm (Horak et al. 2011). In total, 167 samples were peeled during early spring (April) and autumn (October and November) 2008. Those periods are best suited to searching for saproxylic beetles before they start to disperse (spring) and after they stop dispersing (autumn); in addition, the bark is not stuck to the wood by frost as it is in the winter. Each individual adult or larva was then identified by myself and other specialists at least to the family level (for details see Horak et al. 2011).
The same method of bark peeling was used along two altitudinal transects under the Oblachnaya Mt. and through the Gornolyzhnaya Baza near Anisimovka. Both transects were laid out along irregular water streams. Each transect was divided into 50 m a.s.l. sampling locations (Oblachnaya Mt. with ten: 550–1,000 m and Gornolyzhnaya Baza with six: 350–600 m). At each 50 m location, ten pieces of dead wood were inspected that were distributed along a line approximately at a right angle to the transect. The species composition was relatively uniform in the Gornolyzhnaya Baza (nearly 100 % of deciduous trees with occasional occurrence (under 10 %) of fir, spruce and yew). In the transect under Oblatchnaya Mt., 100 % of trees were deciduous at 550, 600, 750, 800 and 900 m a.s.l., and 90 % at 650, 700 and 850 m a.s.l.. 100 % of trees were coniferous at 950, and 90 % at 1,000 m a.s.l.
Predictors studied
The coordinates of each dead wood sample were recorded using a Garmin Colorado 300. The coordinates (x, y) and their squared (x2, y2) and crossed (xy) products were used as a Space category for control of spatial autocorrelation (Horak 2013). In addition to Space, predictors at the Tree level were (1) tree (Table S1), (2) fruiting bodies of fungi, (3) sun exposure, (4) altitude, (5) disposition in the forest, (6) volume and (7) position. Predictors at the Microhabitat level were (1) wetness of the subcorticoulous substrate, (2) bark peeling, (3) bast consistency, (4) bark coverage, (5) bark thickness, (6) bast colour, (7) mycelia and (8) wood consistency (Table 1). All categorical predictors were recoded to the ordinal scale prior to the analyses (Buse et al. 2007).
Altitude was used as the studied predictor in the case of transects. The median value of sun exposure (full, medium and none) and total sampled dead wood volume (m3) were used as copredictors. The Oblachnaya Mt. transect was composed of a mixture of coniferous and deciduous trees; thus, the percentage of coniferous trees was also used as another copredictor.
Statistical analyses
All analyses were done in CANOCO 4.5. The length of the gradient for the total design in detrended correspondence analysis was 7.51 and for the Oblachnaya Mt. transect it was 5.87; thus, canonical correspondence analysis (CCA) was used for further analyses. In the case of Gornolyzhnaya Baza the length was 2.59; redundancy analysis (RDA) instead CCA is often indicated as an appropriate method with shorter length of gradient (Leps and Smilauer 2003)—however, RDA is inappropriate for species abundance data involving null abundances (Legendre and Legendre 1998) hence CCA was used as the most appropriate method. The first step for the total design was the use of all environmental predictors and spatial terms and their squares and cross products as copredictors with 9,999 permutations. Only predictors that had a variance inflation factor <10 were carried forward and used in further analysis due to potential multi-collinearity bias. The CCA with all predictors included was significant (total explained variance = 21.35 %; F = 2.08; P < 0.001). Thus, the subsequent forward selection was not statistically biased (Blanchet et al. 2008). Only predictors with a significant effect (P < 0.05) on the composition of saproxylic beetles were left in the analyses. The explained variance was computed for all significant predictors under 9,999 permutations. Variance partitioning was used to discriminate among the effects of spatial autocorrelation (i.e. Space category) and the two studied categories of Tree and Microhabitat levels. The CCA results were visualized after forward selection using CANODraw 4.12. The individual responses of abundant taxa (with more than five occurrences and more than ten individuals) were computed using species response curves in CANODraw.
Results
In total, 34 beetles and their developmental stages (502 individuals) were found and analyzed (Table S2; Fig S1).
The overall analysis explained nearly 18 % of the variance in the data. The species composition of saproxylic beetles was significantly influenced by Microhabitat and Tree level characteristics, while the effect of Space was not significant and explained the smallest percentage of explained variance. This showed that the results were not biased by spatial autocorrelation. The effects of Tree and Microhabitat characteristics on species composition were nearly identical and shared a relatively low proportion of variance (<1 %), indicating their independence in studies on saproxylic beetles (Fig. 2).

Venn diagram visualization of percentage values of variance explained by the studied categories (Space, Tree and Microhabitat) individually and jointly (circle overlaps) using CCA. ** is for the P < 0.01 and *** is for the P < 0.001 significance level. The negative fraction is given by the correlation structure of the three predictor sets (Legendre and Legendre 1998)
Here (Fig. 3) it can be seen that the composition of saproxylic beetles was, regarding the explained variance, most influenced by discrimination between coniferous and deciduous trees. The other two Tree level characteristics (sun exposure and the presence of fungal fruiting bodies) had lower, though significant, effects on the species composition of the studied taxa. The two Microhabitat factors—the wetness of the bast and bark peeling—were the predictors with the second and third highest and most significant influence on the species composition of saproxylic beetles. The consistency of the bast had the smallest effect on the species composition among the Microhabitat characteristics. The diagram also illustrates that, after the selection of collinear and non-significant predictors, there was only low collinearity among predictors in the first order space (i.e. the first and second axes) and that only sun exposure as the Tree characteristic and bark peeling at the Microhabitat level had relatively the same direct influence on the species composition of saproxylic beetles (Fig. 3).

Environmental plot as a visualization of CCA with six significant predictors derived from forward selection and the species composition of saproxylic beetles as dependent variable. Values in parentheses are the percentages of variance explained by a particular predictor; * is for P < 0.05, ** is for P < 0.01 and *** is for the P < 0.001. CCA Axes 1 and 2 are shown with their values of explained variance
The responses of individual taxa reveal that six of seven abundant taxa were significantly responding to some of the selected predictors (Table 2). Larvae of long-horned beetles preferred early stages of subcorticoulous succession. Flat bark beetle, C. haematodes, indicated a surprising contrast between adults and larvae regarding their preference for coniferous and deciduous tree species, respectively, and the same contrasting situation was evident in the preference for dry underbark substrate by adults and wet subcorticoulous conditions for larvae, while the negative association with a bad bark peeling was the same. Click beetle larvae did not show any significant associations. The larvae of the fire-colored beetle, P. lateralia, showed a preference for dead wood attacked by bracket fungi and, furthermore, for crumbled bast. Larvae of the other fire-colored beetle, S. fuscicollis, had the opposite demands from P. lateralia in terms of the presence of fungal fruiting bodies and preferred sunny habitats—the same as adults of the predaceous carabid, T. nana.
In Oblachnaya Mt., there were observed 19 beetles and their developmental stages (177 individuals) and in Gornolyznaya Baza 21 beetles and their developmental stages with 117 individuals. The response of saproxylic beetle communities to the altitudinal gradients was not significant under the Oblachnaya Mt. (F = 0.99; P = 0.40) and also in Gornolyznaya Baza (F = 0.63; P = 0.66).
Discussion
Most known Eurasian forest ecosystems disturbed by abiotic factors such as windstorms, fire and snow-breakage are immediately attacked by early arriving insects (Jonasova and Prach 2008). Bark beetles in particular are one of the first potential pests that affect revenues from timber production. On the other hand, most of the evidence is from the territory of Europe and North America (Paine et al. 1997). Furthermore, forests in Europe are known to be abused (Grove 2002) and thus they are more sensitive to outbreaks of potential insect pests. Nevertheless, abiotic disturbances are highly important for biodiversity (Bogusch et al. 2014) and some bark beetles are assigned as potential keystone species for ecosystems (Muller et al. 2008).
Management possibilities
The studied area is definitely not a pristine forest (Viitanen et al. 2005), but the non-commercial nature of the reforestation (with maximum use of natural regeneration) most likely leads to a much greater resistance to potentially harmful biotic agents. This is illustrated by the results in this paper, where bark beetles were nearly absent and the most abundant representative of early phloeoxylophages were larvae of long-horned beetles. From the points of view of forest management and conservation of saproxylic taxa, this result indicates what could be targeted in woodland areas with high conservation value—natural regeneration after disturbances is a promising conservation practice (Jonasova et al. 2010). At this point it is necessary to correct the hypothesis that abiotic disturbances must be followed by increasing population densities of early arriving bark beetles (e.g. Mezei et al. 2014). Even allowing for the semi-natural composition of the stands, early arriving saproxylics did not achieve such high population densities—galleries of bark beetles were found in fewer than 4 % of samples.
Response of saproxylic beetles
This study also revealed several results of potential importance with respect to the ecology of subcorticoulous saproxylic beetles. For instance, microhabitat and tree level characteristics seem to be interconnected at a low level as their interaction was very weak. This indicates that modeling on these levels should be manually separated or statistically partitioned (e.g. Horak et al. 2012). The large effect of discrimination between coniferous and deciduous tree species was not so surprising, because it is well known that saproxylic beetles (especially early xylophages) discriminate between these two types of trees rather than being mono- or oligophagous on a particular tree species or genus, while late arriving beetles tend to be more opportunistic (e.g. Slama 1998). The other significant tree level factors indicated that the response of saproxylic beetles is relatively consistent in Europe (i.e. sun exposure and decay stage expressed by the presence of fruiting bodies of fungi; Buse et al. 2007; Horak et al. 2012). The response to the microhabitat factors indicated that subcorticoulous beetles are highly influenced by the diverse condition of phloem together with bark peeling.
Individual species
The responses of individual taxa revealed relatively surprising results. One of the major findings is that adults and larvae of the same species or larvae in the same family might have contrasting demands on particular ecological characteristics of their environment. Larvae of the flat bark beetle, C. haematodes, which is one of the three most threatened saproxylic beetles in Europe (Nieto and Alexander 2010), showed a preference for deciduous trees with a wet subcorticoulous environment, while adults had the opposite demands. This might explain why this beetle is indicated in historical and recent literature as a species preferring conifer forests (e.g. Erichson 1845; Gutowski et al. 2006)—because most records of beetles are connected with adults, and larvae are highly underrepresented. One more point could also be noted—the overlap of larvae and adults of saproxylic beetles in the same habitats of subcorticoulous environment was relatively low (except for C. haematodes). As is indicated, but is generally not well known, in other saproxylic beetles (Park et al. 1931)—since only one incidence of an adult long-horned beetle, no adult fire-colored beetles, and only three incidences of adult click beetles and no larvae of ground beetles were observed for the most abundant taxa.
The responses of individual taxa may also reflect some ecological patterns of saproxylic beetles. As long-horned larvae were significantly associated with badly peeling bark, this might reflect the fact that their type of gallery (with leavings of undigested wood and bast pieces) does not create a suitable environment for secondary underbark associates such as C. haematodes. It is not very probable that this was due to an association of longhorns with early stages of dead wood decay, because most of the dead wood was of the same age. As mentioned above, bark beetles did not arrive and thus there could be no comparison with the underbark environment prepared by scolytids. One prediction could be that some secondary saproxylics (including threatened species) rather prefer bast naturally infested with filamentous fungi, which is the main part of their diet (Prikryl et al. 2012). The lack of response of click beetles to the studied predictors could be explained by their high diversification with respect to their ecological requirements (Horak and Rebl 2013). The main reason may be that for the purposes of this study, some click beetle larvae of more than one species were pooled into a single taxon due to the difficulty of identifying the larvae. Contrasting responses of the larvae of two fire-colored beetles revealed that these two species with morphologically similar larvae have, presumably for reasons of competition, divided their subcorticoulous environment. The ecology of this saproxylic beetle family is still relatively unknown, except for the fact that adults are not saproxylic (e.g. Park et al. 1931). The preference of T. nana for sun-exposed habitats is not surprising, because this predator is highly active under the bark of sun-exposed logs.
Conclusion
Natural regeneration following commercial harvest or natural disturbance of the forest with retention of dead wood, including living trees with microhabitats, could be the goal in sustainable managed woodlands—especially those with high conservation value.
References
Amman GD (1973) Population changes of the mountain pine beetle in relation to elevation. Environ Entomol 2:541–548
Blanchet FG, Legendre P, Borcard D (2008) Forward selection of explanatory variables. Ecology 89:2623–2632
Bogusch P, Blazej L, Tryzna M, Heneberg P (2014) Forgotten role of fires in Central European forests: critical importance of early post-fire successional stages for bees and wasps (Hymenoptera: Aculeata). Eur J Forest Res. doi:10.1007/s10342-014-0840-4
Buse J, Schroder B, Assmann T (2007) Modelling habitat and spatial distribution of an endangered longhorn beetle—a case study for saproxylic insect conservation. Biol Conserv 137:372–381
Carnus JM, Parrotta J, Brockerhoff E, Arbez M, Jactel H, Kremer A, Walters B (2006) Planted forests and biodiversity. J Forest 104:65–77
Christiansen E, Bakke A (1988) The spruce bark beetle of Eurasia. In: Berryman AA (ed) Dynamics of forest insect populations: patterns, causes, and implications. Springer, USA, pp 480–504
Erichson WF (1845) Naturgeschichte der Insecten Deutschlands. Erste Abtheilung. Coleoptera. Dritter band. Nicolaischen Buchhandlung, Berlin
Grove SJ (2002) Saproxylic insect ecology and the sustainable management of forests. Annu Rev Ecol Syst 33:1–23
Gutowski JM, Buchholz L, Kubisz D, Ossowska M, Sucko K (2006) Chrzaszcze saproksyliczne jako wskaznik odksztalcen ekosystemow lesnych borow sosnowych. Lesne Prace Badawcze 4:101–144
Hedgren O, Weslien J (2008) Detecting rare species with random or subjective sampling: a case study of red-listed saproxylic beetles in boreal Sweden. Conserv Biol 22:212–215
Hirzel A, Guisan A (2002) Which is the optimal sampling strategy for habitat suitability modelling. Ecol Model 157:331–341
Horak J (2013) Effect of site level environmental variables, spatial autocorrelation and sampling intensity on arthropod communities in an ancient temperate lowland woodland area. PLoS One 8:e81541
Horak J, Rebl K (2013) The species richness of click beetles in ancient pasture woodland benefits from a high level of sun exposure. J Insect Conserv 17:307–318
Horak J, Zaitsev AA, Vavrova E (2011) Ecological requirements of a rare saproxylic beetle Cucujus haematodes (Coleoptera: Cucujidae)—the beetles’ stronghold on the edge of its distribution area. Insect Conserv Divers 4:81–88
Horak J, Chumanova E, Hilszczanski J (2012) Saproxylic beetle thrives on the openness in management: a case study on the ecological requirements of Cucujus cinnaberinus from Central Europe. Insect Conserv Divers 5:403–413
Jonasova M, Prach K (2008) The influence of bark beetles outbreak vs. salvage logging on ground layer vegetation in Central European mountain spruce forests. Biol Conserv 141:1525–1535
Jonasova M, Vavrova E, Cudlin P (2010) Western Carpathian mountain spruce forest after a windthrow: natural regeneration in cleared and uncleared areas. For Ecol Manag 259:1127–1134
Legendre P, Legendre L (1998) Numerical Ecology, second English ed. Elsevier, Netherlands
Leps J, Smilauer P (2003) Multivariate analysis of ecological data using CANOCO. Cambridge University Press, Cambridge
Mezei P, Grodzki W, Blazenec M, Jakus R (2014) Factors influencing the wind/bark beetles’ disturbance system in the course of an Ips typographus outbreak in the Tatra Mountains. For Ecol Manag 312:67–77
Modlinger R, Holusa J, Liska J, Knizek M (2009) Stav populace lykozrouta smrkoveho Ips typographus (L.) v NPR Zofinsky prales (Novohradske hory, Ceska republika). Silva Gabreta 15:143–154
Muller J, Bußler H, Goßner M, Rettelbach T, Duelli P (2008) The European spruce bark beetle Ips typographus in a national park: from pest to keystone species. Biodivers Conserv 17:2979–3001
Muller J, Jarzabek-Muller A, Bussler H (2013) Some of the rarest European saproxylic beetles are common in the wilderness of Northern Mongolia. J Insect Conserv 17:989–1001
Nieto A, Alexander K (2010) European Red list of saproxylic beetles. Publication Office of the EU, Luxembourg
Paine TD, Raffa KF, Harrington TC (1997) Interactions among scolytid bark beetles, their associated fungi, and live host conifers. Annu Rev Entomol 42:179–206
Park O, Lockett JA, Myers DJ (1931) Studies in nocturnal ecology with special reference to climax forest. Ecology 12:709–727
Pouska V, Svoboda M, Lepsova A (2010) The diversity of wood-decaying fungi in relation to changing site conditions in an old-growth mountain spruce forest, Central Europe. Eur J For Res 129:219–231
Prikryl ZB, Turcani M, Horak J (2012) Sharing the same space: foraging behaviour of saproxylic beetles in relation to diet compounds of their morphologically similar larvae. Ecol Entomol 37:117–123
Slama EF (1998) Cerambycidae of the Czech and Slovak Republics (Beetles-Coleoptera). Milan Sláma, Czech Republic
Svoboda M, Janda P, Nagel TA, Fraver S, Rejzek J, Bace R (2012) Disturbance history of an old-growth sub-alpine Picea abies stand in the Bohemian Forest, Czech Republic. J Veg Sci 23:86–97
Viitanen J, Toppinen A, Hanninen R, Holopainen P (2005) Forest resources, production and exports of roundwood and sawnwood from Russia. Finnish Forest Research Institute, Finland
Wermelinger B (2004) Ecology and management of the spruce bark beetle Ips typographus – a review of recent research. For Ecol Manage 202:67–82
Acknowledgments
I would like to thank Alexander Kohutka for his help with logistics and during the work in Primorsk, Mr. Terwillinger and John W. Dover improved the English. This study was supported by the Internal Grant Agency (IGA no. A10/14), Faculty of Forestry and Wood Sciences, Czech University of Life Sciences, Prague.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Horak, J. What is happening after an abiotic disturbance? Response of saproxylic beetles in the Primorsky Region woodlands (Far Eastern Russia). J Insect Conserv 19, 97–103 (2015). https://doi.org/10.1007/s10841-014-9750-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10841-014-9750-5