
Abstract
There has been much debate regarding the optimal oxygen tension in clinical embryo culture. The majority of the literature to date has compared 5% oxygen to atmospheric levels (20–21%). While the majority of modern IVF labs have accepted the superiority of 5% oxygen tension, a new debate has emerged regarding whether a further reduction after day 3 of development represents the most physiologic system. This new avenue of research is based on the premise that oxygen tension is in fact lower in the uterus than in the oviduct and that the embryo crosses the uterotubal junction sometime on day 3. While data are currently limited, recent experience with ultra-low oxygen (2%) after day 3 of development suggests that the optimal oxygen tension in embryo culture may depend on the stage of development. This review article will consider the current state of the literature and discuss ongoing efforts at studying ultra-low oxygen tension in extended culture.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
In their classic Nature article in 1971, Patrick Steptoe and Robert Edwards reported the first successful culture of a human embryo to the blastocyst stage. In the report, the authors describe the culture conditions that facilitated this breakthrough. More specifically, the investigators noted that the oxygen (O2) tension in their culture system did not utilize atmospheric O2 (approximately 21%). Instead, they described the gas phase as consisting of “5% oxygen, 5% carbon dioxide, and 90% nitrogen” [1].
By the time of this breakthrough, multiple studies had described the in vivo oxygen tension in the female reproductive tract of mammalian species as ranging from 2 to 8% [2, 3]. Furthermore, improved in vitro development of mammalian embryos under physiologic oxygen tension had also been reported [4]. Thus, it was logical that Edwards and Steptoe designed a culture system with reduced oxygen tension in their early experiments.
Yet, for decades, the majority of clinical embryology has been performed at atmospheric O2. The most plausible reason for this is the avoidance of the additional cost associated with reducing the oxygen concentration in the culture environment. Reduced oxygen requires a nitrogen gas system, specialized incubators, and additional quality assurance associated with oxygen sensors [5]. Additionally, multiple early studies failed to demonstrate a statistically significant difference in outcomes at lower oxygen concentrations [6, 7].
However, as more data have accumulated, it has become increasingly clear that a more physiologic oxygen tension results in improved laboratory parameters and clinical outcomes. Multiple meta-analyses now exist demonstrating improved pregnancy rates and live birth rates at 5% oxygen [5, 8]. It is notable, however, that most studies comparing 5% versus atmospheric oxygen tension evaluated outcomes after day 3 embryo transfers. If the goal of the embryo culture system is to recapitulate the in vivo environment, an obvious question for programs performing blastocyst transfers is whether the oxygen concentration of the female reproductive tract is consistent throughout the embryo’s journey from the oviduct, past the tubal ostium, and into the uterus. This question has become more important as an increasing number of clinical ART programs are utilizing extended culture [9].
Indeed, data from multiple mammalian species suggest that the oxygen concentration in the uterus may in fact be closer to 2% [10]. While studies on the normal progression of the preimplantation embryo through the female reproductive tract are lacking, limited data suggest that entry into the uterus may occur around the time of compaction [11]. Thus, a reduction in oxygen concentration at time of day 3 changeover in programs utilizing extended culture with sequential media may in fact represent a more physiologic culture system.
This theory is strengthened by a recent randomized controlled trial by Kaser et al., which compared blastulation rates between embryos randomized to 2 versus 5% oxygen concentration after day 3 of development and found that the “ultra-low” oxygen environment was superior [12]. While these initial data are exciting, further study is needed. The embryos utilized in the Kaser investigation were not intended for clinical use. Indeed, many were previously frozen and some had abnormally fertilized. Thus, to further characterize whether an ultra-low oxygen concentration after day 3 of development does improve the performance of preimplantation embryos in the laboratory, a prospective study of embryos intended for clinical use is necessary.
This review will briefly review how oxygen utilization impacts the physiology of the preimplantation embryo, discuss the current state of the clinical evidence for reduced oxygen concentration, and introduce ongoing efforts at studying the impact of ultra-low oxygen concentration on IVF success.
Characterization of the in vivo oxygen concentration
Embryo culture is a complex task with many components. Each facet of the culture system is built to optimally support the metabolic needs of the preimplantation embryo by attempting to recapitulate the in vivo environment. Not surprisingly, oxygen is an essential component of this system as it plays a central role in cell respiration and strongly influences energy production [13].
Thus, a significant amount of early attention was paid to characterizing the oxygen tension in the female reproductive tract. Initial attempts date back to the 1950s, when analyses of the luminal fluids of the oviduct demonstrated an oxygen tension between 2 and 8% in multiple mammalian species [2, 14]. Comparisons of embryo development under different oxygen concentrations quickly followed. In 1969, Wes Whitten demonstrated that while mouse embryos cultured at 5% oxygen progressed in the laboratory, all arrested when the oxygen tension was set at 20%.
Soon thereafter, Edwards and Steptoe conducted their own experiments comparing embryonic developmental milestones at different oxygen concentrations [1]. Upon successfully culturing the first human blastocyst, the authors remarked: “we seem to have achieved this improved embryonic development through better handling of the cultures than previously…under reduced oxygen tension.” This finding helped to establish 5% oxygen tension as the standard condition for early research in clinical IVF.
Impact of oxygen on embryo physiology
When comparing the performance of preimplantation embryos cultured at different oxygen tensions, a wide variation in indicators of metabolic health has been documented across multiple mammalian species. In sheep, a reduction in oxygen tension from atmospheric levels to more physiologic levels has been demonstrated to increase the catabolic utilization of glucose in preimplantation embryos [15]. Furthermore, improved pyruvate oxidation has been reported under lower oxygen tension in mouse embryos [16]. These factors may conspire to improve the energetic function of preimplantation embryos, supporting biosynthetic activity. Indeed, multiple studies have reported greater cell number and decreased apoptosis in blastocysts cultured at reduced oxygen tensions [17, 18].
While the mechanisms underlying the reduced developmental efficiency of embryos cultured at higher oxygen tensions are not well described, many authors point to an increase in oxidative stress as potential cause of suboptimal development. Reactive oxygen species can damage lipids (affecting the stability of cell membranes), DNA (resulting in abnormal protein synthesis), and proteins (impacting protein function and cell signaling) [19]. Indeed, production of hydrogen peroxide (H2O2) is reduced in pig embryos cultured at 5 versus 20% oxygen. These embryos also demonstrated less DNA fragmentation [20].
Additional analyses have confirmed the notion that gene expression and downstream protein production are influenced by oxygen tension. Rinaudo et al. compared the global gene expression pattern of mouse zygotes cultured at either 5 or 20% oxygen tension and found greater perturbations in expression in the latter group [21]. Other authors have described an increase in global DNA methylation under higher oxygen exposure, suggesting that oxidative stress can also alter the embryonic epigenome [22]. These alterations ultimately can lead to alterations in the proteome. In fact, embryos cultured at reduced oxygen exhibit protein profiles that more closely resembled those from embryos which developed in vivo [23].
Oxygen concentration and IVF outcomes
Despite an abundance of literature suggesting improved performance among embryos cultured at reduced oxygen tension, the majority of clinical IVF has been performed under atmospheric oxygen levels in the decades following the birth of Louise Brown. In fact, a recent survey of embryo culture practices collected from 265 clinics representing 54 countries reported that <25% of programs exclusively utilize 5% oxygen tension during embryo culture [24].
The reasons for this lack of uniform implementation of physiologic oxygen in clinical IVF are multiple. First, maintenance of an incubator at reduced oxygen is associated with greater laboratory costs. Each time an incubator door is opened, the interior atmosphere must be re-equilibrated by instillation of nitrogen gas to purge the environment of excess oxygen [25, 26]. Installation of a modern triple-gas incubator has economic consequences and creates the need for additional quality assurance in the laboratory. Thus, clinics are incentivized to place the burden of proof of benefit on the side of reduced oxygen. Many early studies did not provide this compelling evidence. For example, Kea et al. randomized 106 patients to embryo culture at 5 or 20% oxygen and observed no difference in pregnancy rate between the groups [27]. Additional studies reported similar findings [7, 28].
However, as more clinical data have accumulated, the superiority of reduced oxygen tension has become increasingly clear. Two meta-analyses have been performed which concluded that embryo culture at 5% oxygen results in improved morphologic parameters, reduced embryo arrest, and higher live birth rates [5, 8]. Thus, the added expense and effort of creating a more physiologic culture system with respect to the oxygen environment is justified.
Is there more to the story? Evidence for dynamic oxygen concentration in extended culture
As the evidence in favor of reduced oxygen tension in embryo culture has become increasingly clear, a new question has emerged: is the oxygen tension in the female reproductive tract consistent throughout the embryo’s journey from the oviduct, past the uterotubal junction, and into the uterus? Close review of the initial observational studies describing the oxygen tension in the female reproductive tract demonstrates that the oxygen tension may in fact differ between the oviduct and the uterus.
In an elegant study of hamsters, rabbits, and rhesus monkeys, Fischer and Bavister evaluated the oxygen tension at different phases of the menstrual cycle in both the fallopian tube and the uterus. In the primates, they found the tubal oxygen tension to vary between 5 and 7.8%. However, the intrauterine oxygen tension was significantly lower (1.5–1.85%). In all species, the oxygen tension decreased significantly around the time of blastocyst formation [10].
These findings raise the question of where the embryo is located in the human reproductive tract at the various developmental stages. Surprisingly little is known regarding the chronology of normal progression of the embryo from the tubal ampulla, through the isthmus, across the uterotubal junction, and into the uterus. The best human data come from studies involving surgical sterilization via salpingectomy and uterine aspiration at known intervals following the luteinizing hormone (LH) peak [29, 30]. In one elegant study, 82 oocytes and fertilized ova were recovered from women under these experimental conditions. In the first 72 hours following the LH surge, oocytes, zygotes, and embryos were only recovered from the tube. As the interval from LH surge increased, the proportion recovered from the uterus increased. By 144 hours, none were recovered from the tube. Assuming ovulation at 17 hours after LH peak, the average transit time in the oviduct was estimated to be 80 hours. Interestingly, the most advanced embryo recovered from the tube was an early morula.
While there are certain limitations to this experimental model, these findings raise interesting questions regarding the in vivo environment to which the embryo is exposed at different time points during its transit through the reproductive tract. If it is true that (1) the oxygen tension in the uterus is indeed lower than in the oviduct and (2) the embryo is entering the uterus around the time of compaction, then a dynamic embryo culture system that adjusted the oxygen concentration around day 3 of development may represent the most physiologic system (Fig. 1).

Oxygen tension in the female reproductive tract. Data from primate models suggest that the oxygen tension in the uterus is lower (approximately 2%) than that in the oviduct (5–7%). If the embryo crosses the uterotubal junction around the time of compaction, then a physiologic culture system will reduce the oxygen concentration after day 3 of development
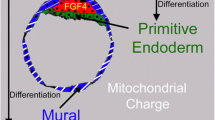
A change in oxygen tension around the time of compaction fits with a general shift in the metabolic strategy of the embryo during this transition [31]. Prior to compaction, embryos produce energy via oxidative phosphorylation [32]. There is no net growth of embryos during this developmental phase and very little oxygen consumption or biosynthetic activity. At compaction, oxygen consumption increases significantly. Glucose becomes the primary energy substrate and energy is primarily produced via aerobic glycolysis (Fig. 2). Differentiation into the trophectoderm and inner cell mass is initiated, and the growth rate of embryos significantly increases. A decrease in oxygen exposure during this developmental phase may protect against an excess in oxidative stress during this period of high metabolic activity [33].

Shifting metabolic strategy of the preimplantation embryo. A change in oxygen tension at time of compaction would be consistent with a general shift in the metabolic strategy of the embryo at time of embryonic genome activation
Ultra-low oxygen tension in extended culture: what is the current evidence?
While the majority of studies have compared a “monophasic” oxygen tension of 5% to atmospheric oxygen tension, some investigators have begun studying the effects of a “sequential” oxygen tension strategy whereby culture is performed at 5% prior to day 3 and at 2% thereafter. These studies are based on the premise that the composition of the culture system should temporally match the in vivo environment as tightly as possible to supply the embryo with the necessary metabolic substrates for a given time in its developmental progression. This idea is similar to the concept of sequential culture media, which involves changing the amino acid and carbohydrate composition of media after day 3.
While the literature is still sparse, available studies are divided in their conclusions. Additionally, findings have been species-specific. One early study examining the impact of a post-compaction adjustment in oxygen concentration compared the developmental milestones of 871 bovine embryos cultured at various oxygen tensions (0, 1, 2, 4, and 7%). In this study, embryos randomized to 2% oxygen produced the highest blastulation rate among the groups [34]. In contrast, Feil et al. performed mouse embryo culture at 7% from zygote to the morula stage and then randomized surviving morulae to either 2, 7, or 20% oxygen concentration thereafter [13]. While the blastulation rates were equivalent among the groups, pregnancies resulting from embryos cultured at 2% experienced a higher resorption rate.
Only two studies have been performed in humans—both reported in 2016. Yang et al. studied 120 donated embryos that had been cultured at 20% oxygen concentration from fertilization until day 3 of development [35]. All embryos in this study had been cryopreserved on day 3. Randomization occurred at time of warming, when the cryopreservation straws containing the embryos were then allocated to culture to the blastocyst stage at 2, 5, or 20% oxygen. The authors found no significant difference in the number of embryos reaching the blastocyst stage or the number of high-quality blastocysts. However, patient-specific parameters were not reported in this study and the randomization scheme produced uneven numbers of embryos among the groups (46 in 2%, 44 in 5%, and 30 in 20%).
A subsequent study by Kaser et al. also utilized donated embryos for evaluation [12]. However, there were important differences in the study design in this trial. First, to control for patient-specific variables, sibling oocytes were utilized for randomization into the intervention (2%) or control arm (5%). Second, all study embryos had been cultured in 5% oxygen until day 3. The authors found an almost twofold increase in the blastulation rate among embryos randomized to 2% oxygen (40.2 versus 22.5%, p = 0.02). Interestingly, among embryos that progressed to the blastocyst stage, cell counts were lower among embryos cultured at 2% oxygen.
Future directions
The implications of a potential improvement in laboratory efficiency with a sequential oxygen tension culture system are exciting from both a biological and clinical standpoint. However, more data are needed. The logical next step to evaluate the utility of reduced oxygen tension in extended culture is to study embryos intended for clinical use.
The ideal investigation would involve two phases. The first study would compare the blastulation rate of sibling embryos randomized to 2 versus 5% after day 3. The sibling embryo model allows control over any patient-specific or laboratory variables that may confound comparisons between the groups. In this experiment, embryologists would evaluate all day 3 embryos and then place all ongoing embryos in a single drop and mix them prior to randomly allocating them to either 2 or 5% oxygen tension for extended culture. This step would be done at low magnification so as to avoid any subconscious bias in allocating embryos to either experimental condition. Embryologists would also be blinded to the exposures at time of blastocyst assessment on days 5 and 6. This study is currently underway at Reproductive Medicine Associates of New Jersey (ClinicalTrials.gov identifier: NCT 02919384).
If the initial investigation suggests superior developmental efficiency of 2% oxygen tension in extended culture, a subsequent study could be performed to evaluate clinical outcomes between the groups. This would be essential as prior studies have demonstrated that improved laboratory parameters following laboratory interventions do not always correlate with improvements in the most important outcomes: achievement of pregnancy and live birth [36]. With alpha error set at 5% and power at 80%, approximately 500 patients would be required to detect a 12% improvement over a 55% live birth rate with euploid embryos. Like all laboratory trials, this study would benefit from the involvement of multiple centers and multiple culture systems to improve external validity.
Conclusions
The issue of oxygen tension in embryo culture has been studied and debated from the earliest days of clinical IVF. While incorporation of the most physiologic culture system has been slow across the world, the evidence is now convincing that reduced oxygen tension produces better laboratory and clinical outcomes. Along with improvements in culture media, reduced oxygen tension has helped spur greater efficiency in culturing embryos to blastocyst and empowered a greater utilization of elective single embryo transfer.
However, a further refinement in the oxygen exposure in extended culture may be on the horizon. Like the initial debates between 5% and atmospheric oxygen, detractors may point to current outstanding blastulation rates enjoyed by many clinics in a monophasic 5% oxygen culture system as evidence that no further changes are necessary. However, it has long been known that human embryos exhibit remarkable resiliency and can develop under a wide variety of conditions. Yet, as always, the focus should remain on providing embryos with the most physiologic culture system as the energetic requirements of adapting to suboptimal conditions undoubtedly come at some cost of impaired viability and lower pregnancy rates. Current studies will help answer the question of whether or not the theory that a greater reduction of oxygen tension to 2% after day 3 will provide more optimal support to the metabolic needs of post-compaction embryos.
References
Steptoe PC, Edwards RG, Purdy JM. Human blastocysts grown in culture. Nature. 1971;29(5280):132–3.
Bishop DW. Metabolic conditions within the oviduct of the rabbit. Int J Fertil Steril. 1957;2:11–22.
Mastroianni Jr L, Jones R. Oxygen tension within the rabbit fallopian tube. J Reprod Fertil. 1965;9:99–102.
Whitten WK. The effect of oxygen on cleavage of mouse eggs. In: Abstracts of 2nd Annual Meeting, Society for the Study of Reproduction, Davis, California; 1969. p. 29.
Bontekoe S, Mantikou E, van Wely M, Seshadri S, Repping S, Mastenbroek S. Low oxygen concentrations for embryo culture in assisted reproductive technologies. Cochrane Database Syst Rev. 2012;(7):Art. No.:CD008950.
Barbieri M, Orlando G, Sciajino R, Serrao L, Fava L, Preti S. High or low oxygen tension—comparison of embryo culture in different incubators. Hum Reprod. 2012;27(Suppl 2). ii62–205. Abstract no. P-174.
De los Santos MJ, Gamiz P, Albert C, Galan A, Viloria T, Perers S, et al. Reduced oxygen tension improves embryo quality but not clinical pregnancy rates: a randomized clinical study into ovum donation cycles. Fertil Steril. 2013;100:402–7.
Nasri CO, Nobrega BN, Teixeira DM, Amorim J, Diniz LM, Barbosa MW, et al. Low versus atmospheric oxygen tension for embryo culture in assisted reproduction: a systematic review and meta-analysis. Fertil Steril. 2016;106(1):95–104.
Kissin DM, Kulkarni AD, Mneimneh A, Warner L, Boulet SL, Crawford S, et al. Embryo transfer practices and multiple births resulting from assisted reproductive technology: an opportunity for prevention. Fertil Steril. 2015;103(4):954–61.
Fischer B, Bavister BD. Oxygen tension in the oviduct and uterus of rhesus monkeys, hamsters and rabbits. J Reprod Fertil. 1993;99(2):673–9.
Croxatto HB. Physiology of gamete and embryo transport through the fallopian tube. Reprod BioMed Online. 2002;4(2):160–9.
Kaser DJ, Bogale B, Sarda V, Farland LV, Racowsky C. Randomized controlled trial of low (5%) vs. ultra-low (2%) oxygen tension for in vitro development of human embryos. Fertil Steril. 2016;106(3):e4.
Feil D, Lane M, Roberts CT, Kelley RL, Edwards LJ, Thompson JG, et al. Effect of culturing mouse embryos under different oxygen concentrations on subsequent fetal and placental development. J Physiol. 2006;572:87–96.
Maas DH, Storey BT, Mastroianni Jr L. Oxygen tension in the oviduct of the rhesus monkey (Macaca mulatta). Fertil Steril. 1976;27:1312–7.
Harvey AJ. The role of oxygen in ruminant preimplantation embryo development and metabolism. Anim Reprod Sci. 2007;98(1–2):113–28.
Khurana NK, Wales RG. Effects of oxygen concentration on the metabolism of [U-14C]glucose by mouse morulae and early blastocysts in vitro. Reprod Fertil Dev. 1989;1:99–106.
Gardner DK, Lane M. Alleviation of the ‘2-cell block’ and development to the blastocyst of CF1 mouse embryos: role of amino acids, EDTA and physical parameters. Hum Reprod. 1996;11:2703–12.
Van Soom A, Yuan YQ, Peelman LJ, de Matos DG, Dewulf J, Laevens H, et al. Prevalence of apoptosis in inner cell allocation in bovine embryos cultured under different oxygen tensions with or without cysteine addition. Theriogenology. 2002;57:1453–65.
Catt JW, Henman M. Toxic effects of oxygen on human embryo development. Hum Reprod. 2000;15 Suppl 2:199–206.
Kitagawa Y, Suzuki K, Yoneda A, Watanabe T. Effects of oxygen concentration and antioxidants on the in vitro developmental ability, production of reactive oxygen species (ROS), and DNA fragmentation in porcine embryos. Theriogenology. 2004;62:1186–97.
Rinaudo PF, Giritharan G, Talbi S, Dobson A, Schultz RM. Effects of oxygen tension on gene expression in preimplantation mouse embryos. Fertil Steril. 2006;86 Suppl 4:1252–65.
Li W, Goossens K, Van Poucke M, Foreir K, Braeckmans K, Van Soom A, et al. High oxygen tension increases global methylation in bovine 4-cell embryos and blastocysts but does not affect general retrotransposon expression. Reprod Fertil Dev. 2014;28(7):948–59.
Katz-Jaffe MG, Linck DW, Schoolcraft WB, Gardner DK. A proteomic analysis of mammalian preimplantation embryonic development. Reproduction. 2005;130:899–905.
Christianson MS, Zhao Y, Shoham G, Granot I, Safran A, Khafagy A, et al. Embryo catheter loading and embryo culture techniques: results of a worldwide Web-based survey. J Assist Reprod Genet. 2014;31:1029–36.
Kasterstein E, Strassburger D, Komarovsky D, Bern O, Komsky A, Raziel A, et al. The effect of two distinct levels of oxygen concentration on embryo development in a sibling oocyte study. J Assist Reprod Genet. 2013;30(8):1073–9.
Gomes Sobrinho DB, Oliveira JB, Petersen CG, Mauri AL, Silva LF, Massaro FC, et al. IVF/ICSI outcomes after culture of human embryos at low oxygen tension: a meta-analysis. Reprod Biol Endocrinol. 2011;9:143.
Kea B, Gebhardt J, Watt J, Westphal LM, Lathi RB, Milki AA, et al. Effect of reduced oxygen concentrations on the outcome of in vitro fertilization. Fertil Steril. 2007;87(1):213–6.
Peng ZF, Shi SL, Jin HX, Yao GD, Wang EY, Yang HY, et al. Impact of oxygen concentrations on fertilization, cleavage, implantation and pregnancy rates of in vitro generated human embryos. Int J Clin Exp Med. 2015;8:6179–85.
Croxatto HB, Ortiz ME, Diaz S, Hess R, Balmaceda J, Croxatto HD. Studies on the duration of egg transport by the human oviduct. II. Ovum location at various intervals following luteinizing hormone peak. Am J Obstet Gynecol. 1978;132:629–34.
Diaz S, Ortiz ME, Croxatto HB. Studies on the duration of ovum transport by the human oviduct. III. Time interval between the luteinizing hormone peak and recovery of ova by transcervical flushing of the uterus in normal women. Am J Obstet Gynecol. 1980;137(1):116–21.
Chason RJ, Csokmay J, Segars JH, DeCherney AH, Armant DR. Environmental and epigenetic effects upon preimplantation embryo metabolism and development. Trends Endocrinol Metab. 2011;22(10):412–20.
Leese HJ. Metabolic control during preimplantation mammalian development. Hum Reprod Update. 1995;1(1):63–72.
Guerin P, el Mouatassim S, Menezo Y. Oxidative stress and protection against reactive oxygen species in the pre-implantation embryo and its surroundings. Hum Reprod Update. 2001;7(2):175–89.
Thompson JG, Simpson AC, Pugh PA, Donnelly PE, Tervit HR. Effect of oxygen concentration on in-vitro development of preimplantation sheep and cattle embryos. J Reprod Fertil. 1990;89(2):573–8.
Yang Y, Yanwen X, Ding C, Khoudja RY, Lin M, Awonuga AO, et al. Comparison of 2, 5, and 20% O2 on the development of post-thaw human embryos. J Assist Reprod Genet. 2016;33(7):919–27.
Karagenc L, Sertkaya Z, Ciray N, Ulug U, Bahceci M. Impact of oxygen concentration on embryonic development of mouse zygotes. Reprod BioMed Online. 2004;9(4):409–17.
Author information
Authors and Affiliations
Corresponding author
Additional information
Capsule
A sequential embryo culture system that reduces oxygen tension from 5 to 2% on day 3 may (1) be more physiologic and (2) improve blastulation rates.
Rights and permissions
About this article

Cite this article
Morin, S.J. Oxygen tension in embryo culture: does a shift to 2% O2 in extended culture represent the most physiologic system?. J Assist Reprod Genet 34, 309–314 (2017). https://doi.org/10.1007/s10815-017-0880-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10815-017-0880-z