
Abstract
Purpose
This study aims to characterize the regulations of histone methylations, key epigenetic markers of oocyte competence, in germinal vesicle (GV) from different follicles (preantral, early, small, or large antral stage) using the domestic cat model.
Methods
In Experiment 1, the incidence of H3K4me3 or H3K79me2 was determined in GVs from the diverse follicle stages directly or after exposure to (1) a methyltransferase inhibitor, (2) sonication to fracture the cytoplasmic membranes and wash away the cytoplasmic content, or (3) methyltransferase inhibitor followed by sonication. In Experiment 2, the presence and maintenance of nuclear methyltransferases SMYD3 and DOT1L (regulating H3K4me3 and H3K79me2, respectively) was characterized in separate GV stages before and after sonication. Functionality of GVs from the various follicle stages (with or without transient isolation from the cytoplasm) then was assessed in Experiment 3 by transfer into recipient competent oocytes.
Results
The incidence of histones H3K4me3 and H3K79me2 within the GV were influenced by the cytoplasmic environment at all stages except at the transition to the early antral stage where nuclear regulating factors appeared to be mainly involved. The methyltransferase SMYD3 and DOT1L also appeared tightly bound to the nucleus at that transition. Interestingly, oocytes reconstructed with a GV isolated from the cytoplasm for a prolonged period had the capacity to form an embryo after fertilization which proved that communication between the donor GV and the host cytoplasm (likely including the regulation of epigenetic factors) could be restored.
Conclusions
Histone methylation apparently becomes regulated by specific nuclear factors at the acquisition of competence during the folliculogenesis and does not seem to be disrupted by prolonged isolation from the surrounding cytoplasm.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
It now is well-established in multiple mammalian species (e.g., mouse [1], pig [2], and cow [3]) that oocyte competence is influenced by epigenetic factors controlling gene expression and chromatin configuration in the germinal vesicle (GV) [4]. In general, epigenetic marks, such as DNA methylation or histone modifications, are changed by enzymes that lead to either gene activation or silencing [5]. For instance, altering histone lysine methylations is known as a core process for governing cell differentiation and appropriate transmission of the required epigenome to ensure normal embryo development [6]. While there are a few macroscale illustrations to date, we know little about epigenetic programming within the oocyte itself, including its sensitivity to the surrounding follicular microenvironment or natural modifications associated with fertilization and embryo development [7].
Earlier studies in our laboratory have determined that the cat GV acquires meiotic and developmental capacity during the changeover from the preantral to antral follicle stage [8]. We also know that GV chromatin configuration during this transition slightly changes from a filamentous to a reticular pattern, while overall gene expression remains detectable throughout the entire nucleus [8]. Histone methylations appear to be the most probable epigenetic factors since modifications may occur independently of the change in GV chromatin configuration, as observed in mouse [9, 10]. Specifically, methylation of histone H3 in mouse oocytes is known to be part of selective gene activation as well as aging [11]. Increased amount of H3K4me3 coincides with oocyte growth [9] and H3K79me2 has been identified as a key activator for transcription during folliculogenesis [12]. In an earlier study, we also reported an important observation pertaining to H3 histones in the cat oocyte. Specifically, methylation of H3K4me3 and H3K79me2 is more prevalent in GVs at the early antral compared to the preantral follicular stage, the former period exactly coinciding with acquisition of meiotic and developmental competence for this species [13]. Therefore, those observations suggested that the kinetics of the process were follicle stage-dependent.
These earlier findings and collective conclusions motivated us to conduct the present study that was designed to decipher and, thus, better understand the role of nuclear versus cytoplasmic factors during folliculogenesis on the capacity to produce a functional intraovarian oocyte. We speculated that this could be discerned by examining the proportions of GVs containing histone methylations at each follicular stage after exposures to methyltransferase inhibitors and/or isolation from the surrounding cytoplasm. An essential component to our strategy was using 3-deazaneplanocin A (DZNep) that has been reported to globally and reversibly inhibit histone methylation in a non-selective fashion [14]. The other critical approach was to remove the cytoplasm by extensive rinsing after oocyte membrane disruption by sonication. Also essential to understanding these developmental mechanisms was knowing the presence and maintenance in the GV of enzymes that are likely regulating these epigenetic patterns during folliculogenesis. Interestingly, the SET (Su(var) 3-9, Enhancer of Zeste (E (Z))) domain of the enzyme SMYD3 has been shown to have H3K4-specific methyltransferase activity [15] and is present in both the mouse and bovine oocyte [9, 16]. The DOT1-like (DOT1L) enzyme has H3K79me2-specific histone methyltransferase activity that can be localized in the mouse GV [5, 17]. Additionally, we have demonstrated that GVs recovered from early stage follicles can be transferred to a later stage, enucleated oocytes that then resume meiosis, fertilize, and sustain early embryo development [8]. Therefore, we additionally tested the restoration of the nuclear-cytoplasmic functions by transferring isolated GVs into conspecific cytoplasts. The possibility of handling isolated GVs before transfer into a cytoplast also is highly relevant to new female fertility preservation strategies including GV desiccation and long-term storage at ambient temperature as an alternative to cryopreservation methods [18, 19]. Lastly, there is now a growing body of evidence of the value of the laboratory cat as a reproductive model for humans, largely due to intriguing morphological and biological similarities between the ovaries and oocytes of the two species, including addressing practical issues such as fertility preservation and reproductive health [8, 20, 21].
The overall objective therefore was to develop new knowledge on nuclear versus cytoplasmic regulation of epigenetic factors that influence GV competence during folliculogenesis in the cat model. In addition, the study would offer new insights into how the GV communicates with nuclear and cytoplasmic factors, information that may be exploited for preserving and using GVs from early stage follicles to produce viable oocytes.
Materials and methods
Collection and classification of oocytes from diverse follicles
Ovaries from adult domestic cats were recovered after routine ovariohysterectomy at local veterinary clinics and were transported immersed in phosphate-buffered saline (PBS) at 4 °C to the laboratory within 6 to 12 h of excision. Cumulus–oocyte complexes (COCs) were collected from large antral follicles (1–3.5 mm diameter) by slicing the follicles visible at the ovarian surface with a scalpel blade in Hepes-buffered Minimum Essential Medium (HMEM; Gibco Laboratories, Grand Island, NY) supplemented with 1 mM pyruvate, 2 mM L-glutamine, 100 IU/ml penicillin, 0.1 mg/ml streptomycin, and 4 mg/ml bovine serum albumin (Sigma-Aldrich, St Louis, MO). Each COC was classified according to standard quality criteria [22] as Grade 1 (uniformly dark cytoplasm, ≥5 compact layers of cumulus cells) or Grade 2 (same as Grade 1, but with 5 cell layers). Lesser grades (mostly partially denuded oocytes) were discarded. Remaining follicles (smaller and deeper within the ovarian cortex) were dissected from the ovarian tissue with a needle and forceps and classified into different categories according to the criteria characterized previously in our laboratory [8]: preantral follicles contained one or several layers of granulosa cells surrounded by a basal lamina; and antral follicles contained a fluid-filled antrum and were categorized into two size groups (<0.5 mm, early antral; and 0.6–1.0 mm, small antral).
Immunostaining of the GV
The oocyte of each follicle was denuded from cumulus cells by immersion in 1.0 % hyaluronidase (Sigma) for 30 min at 38 °C. In each treatment group (see experimental design), oocytes were first fixed in 4 % paraformaldehyde in PBS solution (Sigma) for 30 min at 38 °C. Oocytes were rinsed with wash solution (PBS with 2 % fetal calf serum (FCS; Irvine Scientific, Santa Ana, CA) and 0.5 % Triton X-100 (Sigma)) and blocked with saturation solution (PBS with 20 % FCS and 1 % Triton X-100) for 1 h at 38 °C. Oocytes then were incubated with primary mouse polyclonal H3K4me3 antibody (Abcam, Cambridge, MA) at 1:100 or a primary rabbit polyclonal H3K79me2 antibody (Abcam) at 1:100 in wash solution overnight at 4 °C according to previous reports [6, 9]. A negative control in which the primary antibodies were omitted was included in each trial. After three washes, oocytes were incubated with secondary antibody (goat anti-rabbit or anti-mouse IgG-fluorescein isothiocyanate (IgG-FITC) at 1:100 (Sigma)) for 1 h at 38 °C, and then excess antibody was washed away by three additional rinses.
Targeted methyltransferases were detected using a primary rabbit polyclonal SMYD3 antibody (Fisher Scientific, Rockville, MD) at 1:100 or a primary mouse monoclonal DOT1L antibody (Abnova, Taipei, Taiwan) at 1:100. A negative control in which the primary antibodies were omitted was included in each trial. Oocytes were incubated with secondary antibody goat anti-rabbit IgG-FITC at 1:100 (Sigma) or goat anti-mouse (IgG-FITC at 1:100, for 1 h at 38 °C).
To visualize the chromatin, oocytes were counterstained with 5 μg/ml propidium iodide (PI; Sigma) for 5 to 10 min at 38 °C before mounting with Vectashield mounting medium (Vector Laboratories, Burlingame, CA). Images were captured and analyzed using an Olympus BX41 epifluorescence microscope (Olympus BX41; Olympus Corporation, Melville, NY) and the SPOT 5.0 Advance® Software (Diagnostic Instruments, Inc., Sterling Heights, MI). Oocyte and GV diameters were measured with the SPOT software, and the resulting values were used to confirm the developmental stages of the source follicles (as described above).
For histone methylations and methyltransferases, exposure times from fluorescent light were kept constant for all FITC pictures. SPOT image analysis software was first used to set the detection threshold in negative controls. GVs then were considered as positive when the FITC intensity was above the threshold (the FITC intensity in the GV being normalized to the correspondent chromatin staining with propidium iodide). Within the positive GV, the intensity of FITC fluorescence (reflecting the amount of GV histone methylation or methyltransferase) was expressed in lumen/μm2.
Oocyte treatments
To examine nuclear versus cytoplasmic factors controlling regulation of H3K4me3 and H3K79me2 effects on the GV, oocytes were exposed to one of three different treatments: (1) exposure to 3-deazaneplanocin A (DZNep, a methyltransferase inhibitor; Cayman Chemical, Ann Arbor, MI); (2) sonication to fracture the cytoplasmic membranes and wash away the cytoplasmic content; or (3) exposure to DZNep followed by sonication/isolation. Immature oocytes from preantral, early, small, or large antral follicles were denuded and exposed to 0 or 5 μM DZNep in HMEM for 2 h at 38 °C. Sonoporation or cellular sonication was used to disrupt cellular membranes and wash away the cytoplasmic content. Oocytes were sonicated in 0.5 ml of HMEM in a 2-ml Eppendorf tube by inserting the sonication probe (60 Sonic Dismembrator; Fisher Scientific) midway in the medium and applying five repeated low-strength pulses (setting 3 of the sonicator) with 5-s pauses between each pulse. The sonicated oocytes then were extensively rinsed three times in 1 ml of HMEM before sitting for 1 h at 38 °C. When oocytes were sonicated after the exposure to DZNep, the inhibitor was present in the sonication and washing medium as well.
Oocyte micromanipulations
Recipient oocytes that met grades 1 and 2 criteria (as above) were denuded of cumulus cells via 0.2 % hyaluronidase and gentle pipetting. To visualize the GV, these oocytes were exposed to 7.5 μg/ml cytochalasin B (Sigma) at 38.5 °C for 30 min followed by centrifugation (15,000×g; 5 min) in HMEM to polarize the cytoplasmic lipid droplets [8]. Micromanipulations were conducted in microdrops covered with mineral oil and maintained on a heated (38.5 °C) stage of an inverted microscope (Olympus IX 70) equipped with micromanipulators (Narishige, Sterling, VA). After partial zona dissection with a microlance (Origio, Trumbull, CT), each recipient oocyte was enucleated by extrusion of a small GV-karyoplast that was aspirated in a biopsy micropipette (40 μm inner diameter; Origio) [8]. Each donor GV recovered from preantral, early, small, or large antral oocytes were (1) picked up directly from within the donor oocyte using a ROSNI pipette of 15 μm diameter (Origio) and then immediately injected into a recipient cytoplast or (2) picked up after sonication/rinsing and sitting for 1 h before injection into a recipient cytoplast using the same kind of ROSNI pipette.
In vitro culture and oocyte and embryo assessments
In vitro maturation (IVM) medium consisted of MEM (Sigma) plus 1 μg/ml FSH (NIDDK-ovine FSH-18; National Hormone and Pituitary Program), 1 μg/ml LH (NIDDK-oLH-25; National Hormone and Pituitary Program), and 1 μg/ml estradiol (Sigma). Oocytes denuded for micromanipulation were cocultured with intact oocytes (ratio 1:1 [23]) in 50-μl microdrops under mineral oil (28 h at 38.5 °C in air with 5 % CO2). After fixation in 90 % ethanol and chromatin staining [24], incidence of nuclear maturation was defined as the number of oocytes at the metaphase II (MII) stage relative to the total number of oocytes cultured in vitro. In vitro fertilization (IVF) then was performed using the routine protocol from our laboratory [25]. Briefly, frozen/thawed, motile spermatozoa from a single sperm donor (three males were alternatively used) were selected by swim-up processing in Ham’s F10 medium (Irvine Scientific, Santa Ana, CA) supplemented with 25 mM Hepes, 1 mM pyruvate, 2 mM glutamine, 100 IU/ml penicillin, 100 μg/ml streptomycin, and 5 % FCS (Irvine Scientific; complete Ham’s F10 with Hepes). Cocultured groups of intact and denuded oocytes (ratio 1:1; [23]) were inseminated with 1 × 106 motile spermatozoa/ml in 50-μl microdrops of complete Ham’s F10 without Hepes under equilibrated mineral oil (38.5 °C in air with 5 % CO2). Approximately 10 % of total oocytes in each treatment group were incubated without spermatozoa to assess incidence of spontaneous activation (parthenogenetic control). At 18 h postinsemination, oocytes were cleaned by gentle pipetting. Presumptive zygotes were cultured in vitro for 7 days in complete Ham’s F10 (38.5 °C in air with 5 % CO2) before fixation in ethanol and Hoechst staining. Embryo stages were determined by the number of blastomeric nuclei. A cat embryo with 25 to 45 blastomeres without a blastocoele was defined as a morula [25]. An embryo with a visible blastocoele before fixation and comprised of more than 50 blastomeres was classified as a blastocyst. Blastocyst quality was evaluated by the total number of blastomeres (with more cells indicative of higher quality) [25].
Experimental design and statistical analysis
The objective of Experiment 1 was to determine the role of nuclear versus cytoplasmic elements that may be regulating histone methylation in the GV during folliculogenesis (Table 1). This examination was facilitated by determining the impact of a methyltransferase inhibitor, DZNep, and the isolation of the GV from its surrounding cytoplasm (by sonication). To explore the ability to modify the methylation pattern at each of the four follicular stages, we collected immature oocytes (n = 166; 5 replicates) from preantral, early, small, or large antral follicles that then were denuded and exposed to 0 or 5 μM DZNep. This was followed by fixation and immunostaining of methylated histones (as above). To examine the influence of the absence of cytoplasmic content, another batch of immature oocytes (n = 181; 5 replicates) from the same follicular stage groups were denuded but then sonicated (as described above) before immunostaining (as above). The next step was to determine the combined effect of inhibition and sonication. Here, additional immature oocytes (n = 254; 5 replicates) were exposed to DZNep combined with sonication before immunostaining. Proportional comparisons among groups were compared using uncorrected Chi-square and Mid-P exact tests with differences considered significant at P < 0.05.
The objective of Experiment 2 was to determine the presence and maintenance of the methyltransferases SMYD3 and DOT1L in cat GVs at various follicular stages (Table 1). Immature oocytes from the different follicular types (n = 392; 5 replicates) were treated with or without sonication and then immunostained with SMYD3 and DOT1L antibodies (as above). Proportional comparisons among groups were compared using uncorrected Chi-square and Mid-P exact tests with differences considered significant at P < 0.05.
The objective of Experiment 3 was to investigate the restoration of the communication between GVs and a recipient cytoplasm after isolation (Table 1). As in the other experiments, our strategy also was to examine the influence of follicle stage so oocytes were recovered from preantral (n = 178), early (n = 153), small (n = 166), or large (n = 164) antral follicles (in 6 replicates). Within each stage, half the oocytes were left intact and the other half were used to donate GVs that were transferred to a Grade 1 cytoplast (as above). Half the intact and half of the reconstructed oocytes were fixed after IVM to determine the percentage of fusion, nuclear stage, and MII spindle integrity. The other half was inseminated (as above) with the exception of 115 total oocytes that served as parthenogenetic controls. After 7 days of culture, all oocytes were fixed and assessed (as above) to determine the proportion of cleaved embryos and number of blastomeres per embryo so as to calculate embryonic stage. Percentage data were transformed using arcsine transformation before statistical analysis. Comparisons between treatments and among replicates were evaluated by analysis of variance (ANOVA), Tukey’s multiple test for mean comparison, and a Bartlett test for the homogeneity of the variances. Data not normally distributed were analyzed by the Kruskal–Wallis ANOVA on ranks and the Dunn method for all pairwise comparisons. Differences were considered significant at P < 0.05 (SAS Institute Inc., Cary, NC, USA).
Results
Experiment 1. Presence and regulation of H3K4me3 and H3K79me2 in GVs at different stages of folliculogenesis
Across follicular stages, the proportion of control GVs containing H3K4me3 decreased (P < 0.05) at the transition from the early-to-small antral stage (Table 2). Within each follicular type, the exposure to the methylation inhibitor DZNep led to lower (P < 0.05) proportions of GVs containing methylated H3K4me3 compared to controls, generally by about three times; the exception was at the small antral stage where the difference occurred only at the P = 0.06 level (Table 2). After only sonication/rinsing to isolate the genomic package from the cytoplasm, the proportions of GVs containing H3K4me3 remained unchanged (P ≥ 0.05) compared to controls (Table 2). Additionally, GV exposure to DZNep coupled with sonication/rinsing (in the presence of the inhibitor) reversed the inhibitory effect (P ≥ 0.05) except at the early antral stage where the combined treatment resulted in fewer GVs being positive for H3K4me3 compared to controls (Table 2). The lack of a decrease of positive GVs at the early-to-small antral transition (as observed in controls) occurred because GVs at the early antral stage were highly sensitive to the DZNep exposure that then could not be reversed by sonication/isolation (Table 2). Thus, factors ensuring the presence of H3K4me3 in the GV population at the early antral stage seemed to be independent of the surrounding cytoplasm and likely located in the nucleus. By contrast, maintenance of a specific proportion of GVs containing H3K4me3 at the other follicular stages depended on the cytoplasm.
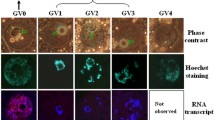
As the stage of the follicle advanced, the proportion of control GVs containing H3K79me2 decreased (P < 0.05) at the small-to-large antral transition stage (Table 3). Within each follicle type, exposure to DZNep alone led to lower (P < 0.05) proportions of GVs containing H3K79me2 compared to controls, generally by about half (Table 3). The inhibitor effect on this histone appeared less pronounced than on H3K4me3 (Table 2). After sonication/rinsing, the proportions of GVs containing H3K79me2 remained unchanged (P ≥ 0.05) compared to controls, with the exception of the early antral stage where high sensitivity to change appeared to be occurring (Table 3). GV exposure to the combined DZNep plus sonication/rinsing treatment had influence (P < 0.05) at the early and large antral follicular stage. This appeared logical as H3K79me2 sensitivity to treatments had already been shown in GVs of early stage follicles (Table 3). At the large antral stage, the combined treatment appeared to cause a reversal of the inhibitor impact (Table 3). The decrease at the small-to-large antral transition stage persisted after DZNep treatment alone likely because sonication/isolation reversed the impact of DZNep or increased the numbers of GVs containing H3K79me2 at the larger antral size. Thus, factors ensuring the presence of H3K79me2 in the GV population at early antral stage appeared to be dependent on the cytoplasmic environment, but they also existed in the nucleus. By contrast, the maintenance of GVs containing H3K79me2 at the other follicular stages depended solely on the cytoplasm. In GVs containing H3K4me3 or H3K79me2, distribution of the methylated histones remained homogeneous in the whole chromatin (Fig. 1) at all follicular stages and regardless of treatment. In controls, the average intensity in FITC-positive GVs ranged from 1.7 to 2.2 lumen/μm2 within each follicular stage and did not differ across stages (P ≥ 0.05). The treatments only affected the proportion of positive GVs but not the intensity of FITC-labeled GVs or the fluorescence distribution. Chromatin configuration of GVs from each follicular stage also was unaffected by the different treatments (Fig. 1a–d, i–l).

Representative chromatin configuration stained with propidium iodide (a–d, i–l) and nuclear distribution of methylated histones stained with FITC (H3K4me3: e–h; H3K79me2: m–p) in germinal vesicles from early antral follicle stages in controls (a, e, i, m), after DZNep exposure (b, f, j, n), sonication/rinsing (c, g, k, o), and DZNep + sonication (d, h, l, p) (Bar = 30 μm)
Experiment 2. Presence and maintenance of methyltransferases SMYD3 and DOT1L in GVs at different stages of folliculogenesis
Most GVs contained SMYD3, although the proportion was lower (P < 0.05) at the large antral compared to the small antral stage (Table 4). Sonication/rinsing reduced (P < 0.05) the proportion of GVs containing that particular methyltransferase at the preantral and small antral, but not the early or large antral stages (Table 4). Those observations suggested that the proportion of GVs containing SMYD3 likely was more dependent on the cytoplasmic environment in the preantral and small antral follicle, whereas the presence of SMYD3 was maintained by the nuclear environment in the early and large antral follicle.
The presence of DOT1L, on the basis of proportions of GVs containing this methyltransferase, was far lower than for SMYD3 at any follicular stage (Table 4). Incidence of DOT1L decreased (P < 0.05) at the preantral and small antral stages, a difference that disappeared after sonication/isolation, and a treatment that increased (P < 0.05) the proportion of DOT1L-containing GVs in small antral follicles (Table 4). DOT1L’s presence seemed maintained solely by the nuclear environment at all stages except in small antral follicles where the cytoplasm was involved. When SMYD3 or DOT1L was present in GVs, distributions were homogeneous within the nuclei and unmodified by the sonication/rinsing treatment (Fig. 2c, d, e, h). In controls, the average intensity in FITC-positive GVs ranged from 2.0 to 2.3 lumen/μm2 within each follicular stage and did not differ across stages (P ≥ 0.05). The sonication only affected the proportion of labeled GVs but not the intensity of FITC-labeled GVs or the fluorescence distribution (Fig. 2d, h).

Representative chromatin configuration stained with propidium iodide (a–b, e–f) and nuclear distribution of the methyltransferases stained with FITC (SMYD3: c–d; DOTL1: g–h) in germinal vesicles from early antral stages in controls (a, c, e, g) and after sonication/rinsing (b, d, f, h) (Bar = 30 μm)
Experiment 3. Functional stability of GVs isolated from the cytoplasmic environment
The percentage of successful GV injection/transfer (based on the absence of oocyte degeneration after 24 h of culture) ranged from 79 to 85 % and was unaffected (P ≥ 0.05) by the donor GV origin. After IVM, oocytes that failed to reach the MII stage showed the same distribution of nuclear status (GV, GV breakdown, and MI) regardless of the treatment group (P ≥ 0.05; data not shown). Although none of the oocytes from preantral follicles achieved MII, the incidence of meiotic maturation increased with the stage of follicle origin (Table 5). However, the incidence of nuclear maturation increased (P < 0.05) when GVs from preantral and early antral follicles were transferred immediately to high quality cytoplasts (Table 5). No such benefit was observed for other donor GVs, and in fact, there was a detriment (P < 0.05) for GVs from large antral follicles. The isolation/rinsing of the GV from the cytoplasm prior to the injection had no impact (P > 0.05) on nuclear status posttransfer (Table 5).
After IVF and embryo culture, there was no evidence of fertilization or development using oocytes or GVs from preantral follicles (Table 5). Although none of the whole oocytes from early antral follicles were fertilized, half of those reconstructed with GVs from this follicular type that achieved MII subsequently cleaved after transferring into high quality cytoplasts. A similar benefit (P < 0.05) occurred after transfer of GVs from small antral counterparts (Table 5). In both cases, embryos reached the blastocyst stage, and there was no disadvantage of isolating/rinsing the scheduled-for-transfer GVs for several hours prior to inserting into a recipient cytoplast (Table 5). GVs from large antral follicles transferred to cytoplasts from the same stage had lower (P < 0.05) meiotic and developmental competence compared to intact controls.
When blastocysts were produced, there were no differences (P ≥ 0.05) among treatments or follicle stages in blastomere number (range, 72–89). There was no (0 %) parthenogenetic activation in any group.
Discussion
We previously have demonstrated the importance of enhanced methylation of the histones H3K4me3 and H3K79me2 at the preantral–antral transition in the cat GV, a time signaling the onset of GV acquisition of competence (resumption of meiosis and sustaining normal embryo development) [8, 13]. The present study provided new information on the nuclear versus cytoplasmic regulation of these epigenetic markers especially in the context of stage of follicle growth. Specifically, we observed in our experimental conditions that the incidence of histones H3K4me3 and H3K79me2 within the GV seems to be mainly controlled by the cytoplasmic environment. However, this was different at the early antral stage when factors within the nucleus became responsible for maintaining H3K4me3 in the majority of GVs. At this same time and by contrast, the incidence of H3K79me2 was being modified by a combination of both nuclear and cytoplasmic factors. The methyltransferase SMYD3 was present in a majority of GVs and appeared more bound to the nucleus in the early antral stages compared to the other stages. Similarly, DOT1L was tightly bound to the nucleus in GVs from early antral follicles. Figure 3 summarizes these observations into a tentative model. Finally, results reaffirmed the resiliency of the GV as an organelle potentially useful in fertility preservation. Even after isolation for 1 to 2 h from the surrounding cytoplasm, it was possible to reconstruct viable oocytes by combining a GV from an immature follicle/oocyte with a high grade cytoplast that then had the capacity to undergo meiosis, fertilization, and embryo development. Therefore, even after the loss of its surrounding cytoplasm, there clearly was an ability of a GV to reconnect and establish communication with a fresh recipient cytoplast.

Nuclear versus cytoplasmic origins of the regulation of histone methylations (H3K4me3 and H3K79me2) and nuclear binding of methyltransferases (SMYD3 and DOT1L) in the germinal vesicle (GV) during folliculogenesis
Others have found in the mouse model that non-manipulated GVs recovered from different follicle stages express variable proportions of histone methylations [4]. However, unlike these other studies that examined batches of GVs from a heterogeneous pool of premature follicles, our study was designed to examine a significant-sized GV population from each of four distinct follicle types (based on well-established, developmental criteria) [8]. This allowed us to make multiple interesting observations, for example, that at any given time, only a portion of the GV population contained H3K4me3 or H3K79me2. However, the quantity of methylated histone appeared constant in positive GVs. At no time did 100 % of GVs express one or the other methylated histone. Secondly, we can hypothesize that the histone methylation status of a given GV was the result of a balance between methyltransferases and demethylases. Thirdly, this is the first case showing that nuclear–cytoplasmic communications can be suspended for an extended period of time before successful oocyte reconstruction.
Interestingly, the use of the DZNep provided a general inhibitory effect on some, but not all, of the GVs in a given follicular population. The quantity of methylated histones also appeared constant in the GVs remaining positive after treatment. The fact that some GVs were resilient to the DZNep treatments may be due to abnormal homeostatic mechanisms in the population of degenerating or atretic follicles naturally present in the cat ovarian tissue [20, 26]. However, H3K4me3 was more influenced by DZNep than H3K79me2. This higher sensitivity to this specific inhibitor in the cat was in agreement with others [14] who have made a similar observation for the H3K4me3 histone using human cell lines. Although, in the latter study, investigators noted an induction of transcription even after H3K4me3 was inhibited, suggesting that the effect of DZNep was not permanent. The duration of effect is not known in our cat system, but also could be transitory based on our ability to effectively reconstruct viable oocytes from isolated GVs. A short-term influence by the DZNep also may be indicated by the lack of a detrimental effect on chromatin configuration and integrity. Regardless, it seemed that both H3K4me3 and H3K79me2 were regulated mostly through cytoplasmic factors, except at the early antral stage when the former was mostly influenced by the nucleus and the latter by both locations (Fig. 3). A particularly important observation was derived from the methyltransferase inhibition before GV isolation (after sonication/rinsing) revealing that inhibitory factors from the cytoplasm could essentially be reversed, enabling the expeditious recovery to a normal methylation pattern. This also confirmed that the transition to the early antral stage was a pivotal moment. The dual effect of nuclear and cytoplasmic factors at the early antral stage may not be coincidental, especially given our earlier observations that it is at this follicular stage that the cat oocyte acquires its meiotic and developmental competence [13].
Our findings were the first to characterize the presence and maintenance of the methyltransferases SMYD3 and DOT1L in the cat GV, both of which are known in other species to be tightly affiliated with histone methylation in the nuclear chromatin [9, 27]. The human gene, SMYD3, contains a SET domain that is a sequence motif found in several chromatin-associated proteins [28, 29]. Others have shown that SMYD3 has an important role in transcriptional regulation and is specific to H3K4 [15, 16]. The existence of SMYD3 in the cat also revealed site-variant regulation with loose association with the nucleus at the preantral and small antral stage and a tight “relationship” at the early and large antral stages. This is in contrast to a recent finding in the bovine oocyte demonstrating that (1) SMYD3 mRNA levels are constant during in vitro oocyte maturation, and (2) immunofluorescence expression for this methyltransferase is completely absent at the GV stage [16]. Interestingly, these same investigators showed that bovine SMYD3 immunofluorescence expression is located around chromosomes during oocyte maturation and transition to the cytoplasmic and nucleus during embryonic development. Thus, findings in both the bovine and cat model agree that cytoplasmic and nuclear factors are regulating histone methylations but at different stages.
Although not reported to contain a SET domain [5], DOT1L is known to be essential for heterochromatin structural formation and, at least in the mouse, specific to H3K79me2 [27]. In our cat model, virtually all GV presence/maintenance was associated with the nucleus, except in small antral follicles where the methyltransferase could leave the nucleus. This observation was in agreement with others where DOT1L was predominantly localized in the nuclei of mouse GV stage oocytes [13]. That the nucleus binding was so important for both of these methyltransferases was illustrated by the lack of impact of sonication/isolation on either SMYD3 or DOT1L expression except at the small antral stage (Fig. 3). This affirmed that there was close linkage with the chromatin, so much so that others have demonstrated that severe disruption of the cytoplasm fails to influence enzymatic expression [15, 27].
How this works functionally is still theoretical, although it has been suggested that GVs from preantral and earlier follicular stages may progressively acquire a nucleoplasmic factor essential for meiotic resumption and subsequent embryo development [30]. The question of type of transport between nuclear and cytoplasmic regulations also is pertinent. Our finding that the nucleus plays a major role in controlling methylations (and at some stages in the complete absence of cytoplasm) demonstrates how epigenetic patterns may maintain a high level of stability. However, there must be a reliable transport system for histones to translocate from the cytoplasm to the nucleus [31]. Others have speculated on the existence of “chaperones” that may modulate histone availability and localization to regulate posttranslational modifications [32]. In this concept, the chaperones actively import histones into the nucleus from the cytoplasm for subsequent assembly into the nucleosome. However, this design has been questioned by others [33] who have noted that posttranslational modifications could also be occurring in the cytoplasm, which already contains the required modifying enzymatic environment [32].
The present work illustrated again the value of the GV transfer technique [30] firstly by demonstrating the ability of a GV from an early stage follicle being inserted into a later (large antral), enucleated cytoplast and fertilizing and becoming an embryo. This finding complemented our original work in this area [8], while confirming that both meiotic and developmental competence was secured in the cat GV before being conferred to the whole oocyte via its cytoplasm. Furthermore, while it continued to be clear that the ability to resume meiosis occurred in GVs from preantral follicles, the intrinsic capacity to properly condense in chromosomes was not fully achieved until the early antral stage. We also learned that the epigenetic and methyltransferase modifications we observed in this study were conserved in the GV, even after isolation from the cytoplasm and that there appeared to be no major disruptions in function when those GVs were translocated into the cytoplast of oocytes from large antral recipients. For GVs from all antral stages, it is possible that the cytoplasmic regulations were reestablished after the transfer into the recipient cytoplast (Fig. 3 and Table 5). Interestingly, SMYD3 and DOT1L that are less bound to the GV at the small antral stage were probably provided by the recipient cytoplast after the GV transfer. The restoration of nuclear–cytoplasmic communication after GV transfer has not been reported. Using cytoplasts from large antral follicles is probably advantageous when conducting GV transfers by conferring some mitigation from the stresses associated with GV isolation and the ongoing epigenetic and methyltransferase modifications. These shifts demonstrate in our experimental conditions the complexity of the regulation of epigenetic factors in GVs, revealing that regulations and maintenance occur over a serial sequence related to stages of intraovarian follicle development.
Our findings also helped to advance our notion that the GV is an attractive target in fertility preservation. The challenges of storing whole oocytes are well known [25], but we speculate that the GV alone could be used for preserving the maternal genome and enhancing reproduction in humans, livestock, laboratory animal models, and endangered species [8, 18, 19]. For such a tactic to be safe requires protecting not only a viable DNA package but also epigenetic patterns that are known to be vulnerable to stressors [34, 35]. Interestingly, vitrification does not significantly alter global DNA methylation of mouse GV stage oocytes that can subsequently mature in vitro [36]. Our studies so far have demonstrated impressive resilience of GV cat oocytes that have been artificially compacted, vitrified, and even isolated from the cytoplasm [37]. When microinjected into a recipient cytoplast, ~50 % of these reconstructed oocytes achieve metaphase II in vitro, ~70 % are fertilized, and ~20 % produce a morula or blastocyst [8]. With the present data, we now have evidence that some epigenetic modifications, including key histone methylations, appear normative and sustained by factors contained in the GV at any stage of cat folliculogenesis. The next validation step is ensuring a functional, normal genome, because to date, our methylation assessments have been limited to the whole nuclear content, not at the level of individual genes. A genome-wide analysis will provide insights into those specific genes potentially influenced by epigenetic modifications that may occur during GV preservation. If the effects of our in vitro processing continue to be non-significant, then the last step will be large-scale, in vivo animal testing to determine if this tool is an effective and safe means of conserving the female genome.
References
Zuccotti M, Ponce RH, Boiani M, Guizzardi S, Govoni P, Scandroglio R, et al. The analysis of chromatin organisation allows selection of mouse antral oocytes competent for development to blastocyst. Zygote. 2002;10:73–8.
Wu C, Rui R, Dai J, Zhang C, Ju S, Xie B, et al. Effects of cryopreservation on the developmental competence, ultrastructure and cytoskeletal structure of porcine oocytes. Mol Reprod Dev. 2006;73:1454–62.
Lodde V, Modina S, Galbusera C, Franciosi F, Luciano AM. Large-scale chromatin remodeling in germinal vesicle bovine oocytes: interplay with gap junction functionality and developmental competence. Mol Reprod Dev. 2007;74:740–9.
Zuccotti M, Merico V, Cecconi S, Redi CA, Garagna S. What does it take to make a developmentally competent mammalian egg? Hum Reprod Update. 2011;17:525–40.
Feng Q, Wang H, Ng HH, Erdjument-Bromage H, Tempst P, Struhl K, et al. Methylation of H3-lysine 79 is mediated by a new family of HMTases without a SET domain. Curr Biol. 2002;12:1052–8.
Zhang A, Xu B, Sun Y, Lu X, Gu R, Wu L, et al. Dynamic changes of histone H3 trimethylated at positions K4 and K27 in human oocytes and preimplantation embryos. Fertil Steril. 2012;98:1009–16.
Spinaci M, Seren E, Mattioli M. Maternal chromatin remodeling during maturation and after fertilization in mouse oocytes. Mol Reprod Dev. 2004;69:215–21.
Comizzoli P, Pukazhenthi BS, Wildt DE. The competence of germinal vesicle oocytes is unrelated to nuclear chromatin configuration and strictly depends on cytoplasmic quantity and quality in the cat model. Hum Reprod. 2011;26:2165–77.
Kageyama S, Liu H, Kaneko N, Ooga M, Nagata M, Aoki F. Alterations in epigenetic modifications during oocyte growth in mice. Reproduction. 2007;133:85–94.
Inoue A, Nakajima R, Nagata M, Aoki F. Contribution of the oocyte nucleus and cytoplasm to the determination of meiotic and developmental competence in mice. Hum Reprod. 2008;23:1377–84.
Shao G-B, Wang J, Zhang L-P, Wu C-Y, Jin J, Sang J-R, et al. Aging alters histone H3 lysine 4 methylation in mouse germinal vesicle stage oocytes. Reprod Fertil Dev. 2015;27:419–26.
Ooga M, Inoue A, Kageyama S, Akiyama T, Nagata M, Aoki F. Changes in H3K79 methylation during preimplantation development in mice. Biol Reprod. 2008;78:413–24.
Phillips TC, Wildt DE, Comizzoli P. Increase in histone methylation in the cat germinal vesicle related to acquisition of meiotic and developmental competence. Reprod Domest Anim. 2012;47 Suppl 6:210–4.
Miranda TB, Cortez CC, Yoo CB, Liang G, Abe M, Kelly TK, et al. DZNep is a global histone methylation inhibitor that reactivates developmental genes not silenced by DNA methylation. Mol Cancer Ther. 2009;8:1579–88.
Hamamoto R, Furukawa Y, Morita M, Iimura Y, Silva FP, Li M, et al. SMYD3 encodes a histone methyltransferase involved in the proliferation of cancer cells. Nat Cell Biol. 2004;6:731–40.
Bai H, Li Y, Gao H, Dong Y, Han P, Yu H. Histone methyltransferase SMYD3 regulates the expression of transcriptional factors during bovine oocyte maturation and early embryonic development. Cytotechnology. 2015.
Ooga M, Suzuki MG, Aoki F. Involvement of DOT1L in the remodeling of heterochromatin configuration during early preimplantation development in mice. Biol Reprod. 2013;89:145.
Graves-Herring JE, Wildt DE, Comizzoli P. Retention of structure and function of the cat germinal vesicle after air-drying and storage at suprazero temperature. Biol Reprod. 2013;88:139.
Elliott GD, Lee P-C, Paramore E, Van Vorst M, Comizzoli P. Resilience of oocyte germinal vesicles to microwave-assisted drying in the domestic cat model. Biopreserv Biobank. 2015;13:164–71.
Bristol-Gould S, Woodruff TK. Folliculogenesis in the domestic cat (Felis catus). Theriogenology. 2006;66:5–13.
Comizzoli P, Songsasen N, Wildt DE. Protecting and extending fertility for females of wild and endangered mammals. Cancer Treat Res. 2010;156:87–100.
Wood TC, Wildt DE. Effect of the quality of the cumulus-oocyte complex in the domestic cat on the ability of oocytes to mature, fertilize and develop into blastocysts in vitro. J Reprod Fertil. 1997;110:355–60.
Godard NM, Pukazhenthi BS, Wildt DE, Comizzoli P. Paracrine factors from cumulus-enclosed oocytes ensure the successful maturation and fertilization in vitro of denuded oocytes in the cat model. Fertil Steril. 2009;91:2051–60.
Comizzoli P, Wildt DE, Pukazhenthi BS. Effect of 1,2-propanediol versus 1,2-ethanediol on subsequent oocyte maturation, spindle integrity, fertilization, and embryo development in vitro in the domestic cat. Biol Reprod. 2004;71:598–604.
Comizzoli P, Wildt DE, Pukazhenthi BS. Impact of anisosmotic conditions on structural and functional integrity of cumulus-oocyte complexes at the germinal vesicle stage in the domestic cat. Mol Reprod Dev. 2008;75:345–54.
Wood TC, Montali RJ, Wildt DE. Follicle-oocyte atresia and temporal taphonomy in cold-stored domestic cat ovaries. Mol Reprod Dev. 1997;46:190–200.
Jones B, Su H, Bhat A, Lei H, Bajko J, Hevi S, et al. The histone H3K79 methyltransferase Dot1L is essential for mammalian development and heterochromatin structure. PLoS Genet. 2008;4:e1000190.
Martin C, Zhang Y. The diverse functions of histone lysine methylation. Nat Rev Mol Cell Biol. 2005;6:838–49.
Gu L, Wang Q, Sun Q-Y. Histone modifications during mammalian oocyte maturation: dynamics, regulation and functions. Cell Cycle. 2010;9:1942–50.
Gao S, Gasparrini B, McGarry M, Ferrier T, Fletcher J, Harkness L, et al. Germinal vesicle material is essential for nucleus remodeling after nuclear transfer. Biol Reprod. 2002;67:928–34.
Nashun B, Hill PWS, Smallwood SA, Dharmalingam G, Amouroux R, Clark SJ, et al. Continuous histone replacement by Hira is essential for normal transcriptional regulation and de novo DNA methylation during mouse oogenesis. Mol Cell. 2015;60:611–25.
Keck KM, Pemberton LF. Histone chaperones link histone nuclear import and chromatin assembly. Biochim Biophys Acta. 1819:277–89.
Poveda A, Pamblanco M, Tafrov S, Tordera V, Sternglanz R, Sendra R. Hif1 is a component of yeast histone acetyltransferase B, a complex mainly localized in the nucleus. J Biol Chem. 2004;279:16033–43.
Wu X-F, Yuan H-J, Li H, Gong S, Lin J, Miao Y-L, et al. Restraint stress on female mice diminishes the developmental potential of oocytes: roles of chromatin configuration and histone modification in germinal vesicle stage oocytes. Biol Reprod. 2015;92:13.
Huidobro C, Fernandez AF, Fraga MF. The role of genetics in the establishment and maintenance of the epigenome. Cell Mol Life Sci. 2013;70:1543–73.
Yan J, Zhang L, Wang T, Li R, Liu P, Yan L, et al. Effect of vitrification at the germinal vesicle stage on the global methylation status in mouse oocytes subsequently matured in vitro. Chin Med J (Engl). 2014;127:4019–24.
Comizzoli P, Wildt DE, Pukazhenthi BS. In vitro compaction of germinal vesicle chromatin is beneficial to survival of vitrified cat oocytes. Reprod Domest Anim. 2009;44 Suppl 2:269–74.
Acknowledgments
This project was funded by the National Center for Research Resources (R01 R026064), a component of the National Institutes of Health (NIH) and is currently supported by the Office of Research Infrastructure Programs/Office of the Director (R01 OD 010948). No conflict of interests to be declared. We thank Dr. Brent Whitaker (Maryland Line Animal Rescue) for providing domestic cat ovaries.
Author information
Authors and Affiliations
Corresponding author
Additional information
Capsule Using the domestic cat model, the incidence and regulation of histone methylations in germinal vesicles during folliculogenesis was studied showing that specific nuclear remodeling events occur during the preantral–antral transition when developmental competence is acquired.
Rights and permissions
About this article

Cite this article
Phillips, T.C., Wildt, D.E. & Comizzoli, P. Incidence of methylated histones H3K4 and H3K79 in cat germinal vesicles is regulated by specific nuclear factors at the acquisition of developmental competence during the folliculogenesis. J Assist Reprod Genet 33, 783–794 (2016). https://doi.org/10.1007/s10815-016-0706-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10815-016-0706-4