
Abstract
We evaluated the effects of the inclusion of Kappaphycus alvarezii (KA), Gracilaria salicornia (GS) and Eucheuma spinosum (ES) as feed additives on the growth, immune and antioxidant status, endocrine variables, nutrient utilization, and nitrogen balance in growing Karan fries calves. Twenty-four growing crossbred calves (body weight 104.00 ± 2.0 kg) were randomly blocked into four groups, each comprising six animals. The feeding schedule was similar in all the groups except that the treatment groups were supplemented with red seaweed (KA, GS, and ES) at 2.5% of dietary concentrate mixture during 180 days of the trial. No significant difference (p > 0.05) was noticed in body weight, average daily gain, dry matter intake and feed conversion ratio among the groups. Nutrient digestibility and nitrogen balance remained unaffected. There was no effect (p > 0.05) on haemato-biochemical parameters due to red seaweed supplementation. The levels of superoxide dismutase (SOD), catalase (CAT) and serum GSH-Px were similar in all the treatment groups. The addition of seaweeds did not exert any effect on mean serum levels of triiodothyronine (T3), thyroxine (T4) and cortisol. The serum concentration of immunoglobulin (IgG) was significantly higher among the seaweed-supplemented groups than the control group. Conclusively, the dietary supplementation of tropical red seaweeds at 2.5% in the concentrate mixture may significantly influence immunity without any alteration in hormonal profile, antioxidant status, serum metabolites, enzymes, and performance of growing crossbred calves.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Seaweeds (macroalgae) are an immense renewable resource that is not utilized in the world economy. Seaweeds do not require fertilisers, pesticides, or freshwater to grow and can grow on less surface area than that by terrestrial plants. The consideration for this extremely low-cost, huge, and relatively easy-to-cultivate animal feed source is continually developing and its utilization is diversifying into other areas of application. Seaweeds comprise approximately 6000 species of various forms and sizes. However, only 5% of the seaweeds are utilized as meals for both humans and livestock (Chojnacka et al. 2012). The most cultivated species of edible seaweed are the kelp (Saccharina japonica), which gives accounts for about 60% of the whole seaweed production followed byPorphyra, Kappaphycus, Undaria, Eucheuma and Gracilaria. Since 1970 seaweed production has been constantly growing with an average annual growth rate of 7.7%. In 2011 nearly 21 million t of aquatic plants were produced globally, valued at US$5.5 billion, of which 99% was seaweed production (by quantity or value). However, up to 2004, 1% of the seaweed value globally was used for farm animal feed only, wherein Ascophyllum nodosum (brown seaweed) constituted a maximum of US$10 million to the world economy (Chopin and Sawhney 2009; FAO and IFIF 2010; FAO 2013). There have been many studies on the inclusion of seaweeds in the diets of pigs, cows, sheep, and poultry. In those studies, nutritional supplementation of even a small amount of seaweed or its components improved the nutritional composition of feed and enhanced the overall performance of the livestock. For example, seaweed supplementation enhanced the microbial counts in the intestinal tract and improved the immune response in pigs (O’Doherty et al. 2010). Supplementary diets are certainly more suitable for rumen fermentation (Choi et al 2021), and digestion in cows, which impact milk quality and cattle meat (Bendary et al. 2013; Hwang et al. 2014; Maheswari et al 2021). Also, supplementation of seaweed is beneficial to alleviate stress in goats and sheep and is stated to lower the level of cholesterol in eggs and improve chicken wellbeing (Carrillo et al. 2008), suggesting the possibility for its industrial usage in livestock feeds (www.oceanharvest.ie). Thus, the incorporation of seaweed in animal feed could provide a great opportunity in feed formulation. According to Bendary et al. (2013) seaweed supplementation enhanced nutrient digestibility, milk yield and elements, feed conversion, rumen fermentation activity, and blood serum proteins when lactating Friesian cows were fed with commercial seaweed at the rate of 50 g seaweed per head per day. However, data on tropical seaweed as a feed supplement are comparatively few and are mostly on in vitro fermentation and anti-methanogenic activity. Thus, insufficient research has been done on the supplementation of three abundantly available tropical red seaweeds (Kappaphycus alvarezii, Gracilaria salicornia and Eucheuma spinosum) on rumen fermentation, blood biochemical profile and growth performance of crossbred calves. This study, therefore, assessed the comparative effects of supplementation of these tropical red seaweeds on growth performance, nutrient intake, haemato-biochemical parameters, antioxidant status and immunological and endocrine variables of growing crossbred calves.
Materials and methods
The experiment was carried out at the ICAR-National Dairy Research Institute (NDRI) particularly at the Livestock Research Centre in the Animal Nutrition Division shed in Karnal, Haryana, India. The Institutional Animal Ethics Committee (IAEC), which was established following Article Number Thirteen of the Committee for Control and Supervision on Experimentation on Animals (CPCSEA) regulations, was consulted before the experiment in order to obtain the necessary permission (Government of India).
The seaweed by-products were purchased from Aquagri Processing Private Limited in New Delhi. The fresh seaweeds were sun-dried to about 10% moisture content, thrashed by hand on a thrashing bed, and ground in a hammer mill. The three tropical red seaweed species used in this trial were K. alvarezii, G. salicornia and E. spinosum. Each was mixed thoroughly at a 2.5% level in a concentrate mixture.
Animals and experimental design
Twenty-four growing crossbred calves (Karan Fries) were selected from the ICAR-National Dairy Research Institute's, Livestock Research Centre in Karnal, and the animals were given a 14-day adaption period. After the adaptation period, the calves in the experiment were randomly divided into four groups (six calves in each group) based on their body weight. The experimental diets were protein and energy balanced. To meet their nutrient requirements, all the calves were fed according to the ICAR (2013) feeding standard recommendation. Calves of the control group (C) were fed a standard diet without seaweed. The concentrate mixture of calves in groups KA (K. alvarezii), GS (G. salicornia), and ES (E. spinosum) was offered concentrate mixture containing 2.5% of seaweed by-products alongside the basal diet for 180 days.
Housing and feeding management
The experimental crossbred calves were housed in a well-ventilated shed arranged for animal feeding wherein an individual animal has no access to another animal's feed. During the 180-day experiment, the shed was washed twice daily and thoroughly cleaned to remove faeces, urine, and dirt. The calves were dewormed against internal parasites based on their body weight before the start of the experiment and then at 3 months intervals. They also were treated against external parasites.
The quantity of concentrate mixture, maize fodder and wheat straw fed to the calves, obtained from the ICAR feed mill were measured and these feeds were offered daily. To meet the calves' nutrient requirements (ICAR, 2013), the quantity of concentrates fed to each calf was determined based on body weight. Before feeding, the required amount of seaweed by-products was thoroughly mixed with the concentrate mixture. At different intervals, maize fodder chaffed at 1.5 inches and chaffed wheat straw at 0.5 inches were offered. All calves were given clean and fresh drinking water ad libitum twice daily at 9.00 a.m. and 18.00 a.m. The materials of the concentration combination and common salt were supplied by Ram Chander Chhabil Dass of Hissar, and the mineral mixture was supplied by LUVAS of Hissar. The concentrate mixture was prepared by the NDRI feed mill plant, and Table 1 lists the ingredients of the experimental diet. The feeding schedule was changed every two weeks after the weight was recorded in order to supply nutrients as per the ICAR (2013) recommendation.
Sampling and measurement
Body weight, average daily dry, matter intake (DMI)
All the calves were weighed on an electronic scale for two consecutive days before beginning the experimental feeding, and subsequently body weight was measured every two weeks throughout the experimental feeding period of 180 days. Fortnightly Dry matter (DM) intake/daily DM intake was measured from the record of daily feeding of the measured quantity of concentrate mixtures of available green fodder and residues of feed ingredients. Feeds and residues were sampled twice a week for dry matter estimation in order to ascertain dry matter intake by each of the animals.
Sampling of blood and analyses
For the analysis of haematological, biochemical, hormonal, immune, and antioxidant activities, blood samples were collected from each calf at the start of the trial and then at monthly intervals (0th, 90th, and 180th day) by puncturing the jugular vein. Blood samples (10 mL) were collected into CAT Serum Activator Vacutainer tubes (BD Franklin, USA) and were kept in a slanting position for 45 min. The serum was separated by centrifuging the samples at 3000 rpm for 15 min after which it was collected in small plastic vials (5 mL) and stored at -20 °C for further analysis. In order to prepare the lysate, 2 mL of blood was collected in Eppendorf tubes containing 0.3 mL of acid-citrate dextrose solution. Blood samples were centrifuged at 2500 rpm for 20 min to remove the plasma and buffy coats. The packed cells were resuspended in the same solution to give a 33% suspension after the erythrocytes had been washed thrice with isotonic washing solution with phosphate buffer saline, pH 7.4. Antioxidant enzyme activity like catalase (CAT) and superoxide dismutase (SOD) were estimated 2 days after collection and processing of blood (Aebi 1984; Madesh and Balasubramanian 1998). Determination of glutathione peroxidase (GSH-Px) (Catalog No. E0006Bo) serum concentration was carried out by using specific bovine-ELISA GSH-Px quantification kits (Bioassay Technology Laboratory, China). GSH-Px was quantified following the manufacturer's recommendations. The detection range for GSH-Px was 1 to 300 ng mL−1, and the minimum detectable dose was less than 0.58 ng mL−1. The intra and inter-assay coefficients of variance (CV) were less than 8% and 10%, respectively.
Haematological parameters haemoglobin (Hb), white blood cell (WBC) count, red blood cell (RBC) count, and packed cell volume (PCV) were analysed using an MS4SE haematology analyser (Melet Schloesing Laboratories, France). The analysis was made within 1 h of blood collection. Serum biochemical profiles of glucose, total protein, albumin, globulin, creatinine, total cholesterol, total triglyceride, urea, and activity of liver serum enzymes aspartate transaminase (AST), alanine transaminase (ALT) and alkaline phosphatase (ALP) were determined using a commercial kit (Recombigen Laboratories Pvt. Ltd. New Delhi, India—cat no-GLU-L100, TTP-L100, ALB-L100, CRT-L100, CHO-L100, TGL-L100, URB-P100, AST-L100, ALT-L100 and ALP-L100 respectively). Globulin was calculated by subtracting albumin from total protein content.
Serum concentrations of hormones such as thyroxin (T3) (Catalog No. E0215Bo), triiodo-thyronine (T4) (Catalog No. E0216Bo) and cortisol (Catalog No. E0110Bo) were using kits from Bioassay Technology Laboratory, Shanghai, China. These kits are based on the ELISA principle. The minimum detectable dose for T3 was less than 0.01 ng mL−1, for T4, was less than 2.61 ng mL−1 for cortisol was less than 0.02 ng mL−1 and the detection range was 0.03 ng mL−1 to 9 ng mL−1, 5 ng mL−1 to 600 ng mL−1 and 0.5 ng mL−1 to 200 ng mL−1 for T3, T4 and cortisol respectively. The intra and inter-assay coefficients of variance (CV) were less than 8% and 10% for T3, T4 and cortisol.
Each plate was pre-coated with antibodies specific to bovine hormones (T3, T4 and cortisol). The various hormones found in the samples were bound to the antibodies coated on the wells. Following sample binding, a biotinylated detection antibody was added, and it bound with the captured hormone in the sample. Streptavidin-conjugated horseradish peroxidase (SA-HRP) was then included which was bound to the biotinylated antibody. After incubation, washing was used to remove unbound streptavidin-HRP. After that, a substrate solution was added. A blue result from the colourimetric reaction turned yellow when the reaction was stopped by adding diluted sulfuric acid (stop solution). The amount of a certain hormone present in samples was inversely related to the yellow product's absorbance at 450 nm. The standard solutions were prepared by reconstituting them with standard diluent after serial dilution to obtain the range of standards. A standard curve was prepared by plotting the absorbance obtained against the standard concentration where the corresponding hormone concentrations were calculated. To guarantee the development of colour in the sample well, a row of blank wells was retained in each plate. The test's reliability was determined by comparing the results of the control group to the normal reference values for each estimation. Serum concentrations of immunoglobulin G (IgG) (Catalog No. E0010Bo) were performed by using specific bovine-ELISA IgG quantification kits (Bioassay Technology Laboratory, China). The IgG was quantified based on the manufacturer’s directions. The detection range for IgG was 2 to 600 g mL−1, with a minimum detectable dose of less than 1.03 ng mL−1. The intra and inter-assay coefficients of variance (CV) were less than 8% and 10%, respectively.
Metabolic trial
In order to assess the nutrient digestibility and nutrient balance, a metabolism trial of 6 consecutive days collection period was carried out on all the calves during the end of the experimental trial. During the 24 h feeding cycle, the collection period of 6 days of an accurate record of feed intake, refusals, and faeces were voided and urine excreted by each animal was kept. Representative of these samples such as feed offered, residues, faeces and urine were collected at 24 h intervals. The faecal and urine samples were stored in 25% H2SO4 for nitrogen (N) estimation and for dry matter determination the faecal sample were also collected and kept for proximate analysis. To determine the dry matter (DM) content, feed, residues, and faeces were dried in a hot-air oven at 100 °C for 24 h. Dried samples of feed, refusals, and faeces were ground in a grinding mill to pass through a 1-mm screen before being stored at room temperature until further laboratory analysis. Proximate analyses of dry matter (DM), organic matter (OM), crude protein (CP), and ether extract (EE) were determined using the Association of Official Analytical Chemists' standard protocols (AOAC 2005). Neutral detergent fibre (NDF) and acid detergent fibre (ADF) were estimated according to Van Soest et al. (1991). Nutrient digestibility was calculated following the formula Nutrient digestibility (g kg−1) = Nutrient intake – Nutrient output/Nutrient intake described by Singh et al. (2021).
Statistical analysis
The data recorded during the 180 days of the trial like body weight, dry matter intake, metabolic trial data such as intake and digestibility of nutrients and nitrogen balance were subjected to one-way analysis of variance (ANOVA) using the general linear model procedure of Statistical Package for the Social Sciences (SPSS for Windows, V21.0; SPSS Inc., USA). Haemato-biochemical parameters, endocrine variables, antioxidant activity, and immunoglobulin G parameters were similarly subjected to two-way analysis of variance (ANOVA). The pair-wise comparison of means was carried out using Tukey’s honest significant difference (HSD) test. Significance was determined at P < 0.05.
Results
Proximate analysis of feedstuff and seaweed by-products
The chemical composition, viz maize fodder, wheat straw, concentrate mixture and seaweed by-products of K, alvarezii (KA), G. salicornia (GS) and E. spinosum (ES) and used during the feeding trial are given in Table 2. The dry matter (95.82%), total ash (51.12% on a DM basis), and crude protein (6.84%) contents of GS were found to be higher than ES and KA but organic matter (48.88%), and nitrogen-free extract (26.40%), contents of GS was found lower than ES and KA. However, the dry matter (94.51% and 93.65% respectively), organic matter (66.86% and 67.58% respectively) and crude protein (5.60% and 6.14%, respectively), acid detergent fibre (13.81% and 11.77%, respectively) contents of ES and KA were comparable.
Growth performance and feed conversion ratio
The effect of the experimental diet on initial body weight, final body weight, average daily gain, dry matter intake, DMI (% body weight), DMI (g kg−1 BW0.75), and feed conversion ratio during the 180th day of feed trials are provided in Table 3. Throughout the 6-month study period, the initial body weight, final body weight, and average daily gain were found to be similar among the groups. Similarly, there was no changes in dry matter intake, DMI (% body weight), DMI (g kg−1 BW0.75), or feed conversion ratio between groups.
Nutrient intake, digestibility, and Nitrogen balance
The data relating to nutrient intake and nutrient digestibility, and N balance are given in Tables 4 and 5. Dry matter intake (%BW) was found similar (p = 0.419) between treatment groups during the digestion trial. Seaweed inclusion had no effect on OM, CP, NDF, ADF, EE, or TDN (total digestible nutrients) intake. Likewise, similar results were detected in treatment diets on the nutrient digestibility (g kg−1 DM) of DM, OM, CP, NDF, ADF, and EE between the different treatment groups. Also, nitrogen intake (g day−1), faecal output (g day−1), N absorbed (g day−1), urinary nitrogen losses (g day−1), N retention (% N intake), and N absorbed (% N intake) were all comparable among the groups.
Serum haemato-biochemical profile
Data on the serum haemato-biochemical parameters are presented in Table 6. Critical observation of data revealed that the concentration of WBC, RBC, Hb, and PCV count were not influenced during the 180th day of the feeding trials in the different treatment groups. However, the levels of WBC, RBC, and PCV counts were increased significantly (p < 0.001) during period means at the advancement of calves' age. Similarly, serum biochemical parameters showed comparable results in glucose, total protein, albumin, globulin, creatinine, total cholesterol, total triglyceride, and urea during the 180th day of the feeding trials between groups but were within the normal reference range (Kahn and Line 2010) for healthy calves. Comparable effects were detected in liver serum enzymes like AST, ALT, and ALP during the 180th day of the experiment between the groups. The levels of ALP varied within the normal range (68–387 U L−1) (Kaneko et al. 1989). However, ALP significantly (P < 0.001) differed with the advancement of calves' age.
Serum endocrine variables, antioxidant status and immune response
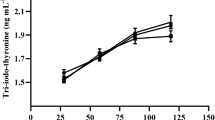
The results on endocrine variables (T3, T4 and cortisol), antioxidant profile activity and immunoglobulin (IgG) are presented in Table 7. The data analysis revealed that there were no changes between the treatment and control groups for T3, T4 and cortisol levels. A significant effect (p < 0.001) was observed in calves on T4 with the advancement of the calves' age during the feeding trials. Similarly, no changes were found in SOD, CAT and GSH-Px enzyme activity in calves among all the treatment groups. Conversely, a significant change (p = 0.045) was observed in serum IgG concentration in calves when seaweeds were supplemented during the 180th day of feeding trials. Further a period wise significant (p < 0.001) effect was also noted as the advancement of age in calves.
Discussion
Proximate analysis of feed ingredients and seaweeds
The nutritional compositions of maize fodder, wheat straw, concentrate mixture and chemical composition of three red seaweeds used during the feeding trials were as contained in previous reports (Ahmad et al. 2012; Rajauria 2015; Diharmia et al. 2019; Sharma and Datt 2020).
Growth performance
One the 180th day of the trial, there was no significant variation in initial body weight, final body weight, average daily gain, dry matter intake, DMI (% body weight), DMI (g kg−1 BW0.75), and feed conversion in any of the treatment groups. In line with our present study, Hong et al. (2015) reported that dietary inclusion of a brown seaweed by-product to Holstein cows (0, 2 and 4% levels) had no significant effect on daily gains during 275 days of the trial period. Anderson et al. (2006) also found no effect of A. nodosum inclusion on the growth performance of crossbred cattle when the supplement was given at different periods of 36–50 days, the last 14 days of feeding and both periods combined. When lambs were fed a seaweed containing diet at 0, 3 and 5% levels, the ADG and feed intake were stayed similar among all the supplemented groups (El-Waziry et al. 2015). Pickard et al. (2008), on the other hand, studied a 30% Sargassum spp. supplementation level as a dietary replacement in small ruminants and discovered a positive influence on growth performance. In line with the above mentioned study, Alagan et al. (2020) reported that feeding Azolla (5%) along with green seaweed U. lactuca (3%) resulted in the highest body weight gain, improved shank length, and feed efficiency in the seaweed plus Azolla group. Leonard et al. (2010) observed a higher ADG between days 0 and 21 post-weaning than pigs weaned from non-seaweed-supplemented sows. However, the DMI in all the treatment groups was unaffected which showed the feed was palatable and the level of supplementation of the seaweeds did not influence the goodness of the feed as earlier reported (Lee et al. 2005; Kinley et al. 2020; Sharma and Datt, 2020). Conversely, Leupp et al. (2005) showed significant effects on DM intake once brown seaweed meal was added to the diet of steers fed with hay that had low quality. Supplementation of red seaweed in calves on a different fortnight of feed conversion ratio was similar between the groups. Generally, the data on the use of seaweed in ruminant trials have produced inconsistent results and this could be attributed to the variations in the seaweed genera (Palmaria, Ulva, Ascophyllum), species (red algae, green algae, and brown algae), and discrepancies in the techniques used for harvesting and processing algae including collection, cleaning and drying, or the proportions used (Melton 2001).
Nutrient intake, digestibility, and nitrogen balance
From the results, it was revealed that the intake of nutrients (kg) between treatment groups did not change concerning the inclusion of red seaweed by-products. Statistically, the data, shows that seaweed by-products inclusion of up to 2.5% of concentrate mixture did not influence the palatability of the diet, therefore the dry matter intake stayed similar among the groups. Likewise, the cell wall constituents were not influenced by seaweed inclusion as the result, the TDN values in the different groups were similar. The current finding is consistent with the findings of Singh et al. (2015), who found that when Sargassum wightii extract was included in the concentrate mixture at a 20% level, the plane of nutrition had no effect on lactating Sahiwal cows. Additionally, Lee et al. (2005) found that adding 800 g day−1 of brown seaweed (Undaria pinnatifida) to dairy cow diets had no significant (p > 0.05) impact on DM intake. The results of the present study do not coincide with those of Cabrita et al. (2017) who found that adding alfalfa hay along with Ulva rigida and Gracilaria vermiculophylla up to 25% of the diet on a dry matter basis reduced DM intake by 24 and 25% respectively. El-Din et al. (2008) found that supplementation of kelp meal significantly (p < 0.05) increased the DM, TDN, and DCP intake in calves throughout the trial periods. The differences in responses found in these different studies as indicated earlier could have been due to many factors like the dose and kind of seaweed, the physiological condition of the animal and environmental conditions that have to do with where the experiment was carried out. It should be noted that red seaweeds contain a lot of other phenolics, whereas brown seaweeds contain phlorotannins (Ganesan et al. 2008). According to Wang et al. (2008) the phlorotannins mediated decrease in protein digestibility is either due to the decrease in microbial activity or due to a direct protective effect of phlorotannins on dietary protein. In the present research, it is evident that the inclusion of seaweed by-products has no impact on the digestibility of DM, OM, CP, EE, NDF, and ADF. The nutrient digestibility remained similar (p > 0.05) between the different groups. In general, the results of this study coincide well with many other reports on digestibility that showed no influence on seaweed meal inclusion in lactating animals (Antaya et al. 2015; Singh et al. 2016; Zhou et al. 2018; Maheswari et al. 2021). Sharma and Datt (2020) found no change in digestibility coefficients in lactating Karan Fries cows when K. alvarezii-based feed additives at levels 1.5 and 3% were supplemented in their ration. Similarly, Munde (2018) reported no changes in nutrient digestibility in crossbred cattle when seaweed by-products were included in their diet. Another study by Gardiner et al. (2008) stated no significant change in swine digestibility when fed with A. nodosum in their diet. Nitrogen balance was not affected (p > 0.05) by the inclusion of seaweed by-products. The current study findings corroborate well with those of Sharma and Datt (2020) who found no change in nitrogen intake and nitrogen voided through faeces, or urine in lactating Karan Fries cows when K. alvarezii-based feed additives at levels 1.5 and 3% were supplemented in their diet. A similar study by Hopkins et al. (2014) and Rjiba-Ktita et al. (2019) revealed that N retention did not show any significant impact by feeding seaweed powder. Similarly, Maheswari et al. (2021) reported a non-significant difference in N utilization in Murrah buffaloes when brown seaweed Turbinaria conoides along with two red seaweeds K. alvarezii and G. salicornia were included in the animal diet. In contrast to the results of this study, Gardiner et al. (2008) reported that supplementation of A. nodosum powder numerically reduced nitrogen digestion and retention in pigs.
One of the concerns about feeding seaweed, particularly brown seaweed, is the presence of phlorotannins, which bind proteins and reduce proteolysis, potentially reducing the degradability of N in the rumen (Wang et al. 2008). In the present experiment, the entire seaweed never surpassed 1% of the diet. Out of the three products verified, they did not contain any brown seaweed. In general, the results of this trial agree with Singh et al. (2017) who observed no significant difference in nitrogen intake, its excretion through faeces, urine, and milk, N absorption and balance, as well as N balance as percent of N intake and percent of N, absorbed when seaweed powder was added to their ration.
Serum haemato-biochemical profile
The serum haematological analysis showed no significant influence on WBC, RBC, Hb, and PCV among the groups and was within the normal physiological range as reported by Karatzia et al. (2012). A significant period effect was observed as the age of the calves advanced in the study. Similar findings of these experiment agree with some previous studies which reported that WBC, RBC, Hb, and PCV was not significantly influence by feeding seaweeds (Karatzia et al., 2012; Chugh 2020). Munde (2018) also reported no change in counts of PCV, RBC, and WBC during his study period when both K. alvarezii and G. salicornia were supplemented in the diet of crossbred cattle. Abdoun et al. (2014) also reported that blood PCV in lambs did not differ significantly due to dietary supplementation of seaweed (Ulva lactuca) extract. In this present study, Hb and PCV of crossbred calves were measured to evaluate complete health status. Red blood cells are a good model for studying oxidative stress because they are sensitive to oxidative damage (Saker et al. 2004). Likewise, blood-PCV and Hb values provide information in relation to disorders like anaemia, lack of amino acids, etc. (Ndlovu et al. 2007) whereas WBCs are connected to the immune system function (Archer et al. 2007). Since the current study did not show significant changes among controls and calves offered seaweeds (E. spinosum, K. alvarezii, and G. salicornia), it can be assumed that the seaweeds under study had no negative effects on erythropoiesis, and thus, the health of the calves.
The serum concentration of glucose, total protein, albumin, globulin, creatinine, total cholesterol, total triglyceride, urea, aspartate aminotransferase (AST), alanine aminotransferase (ALT), and alkaline phosphate (ALP) was similar (p > 0.05) between the different treatment groups. Likewise, serum concentration of glucose, total protein, albumin, globulin, creatinine, total cholesterol, total triglyceride, urea, AST, ALT, and ALP as detected in this trial coincide well with other researchers (Abd Ellah et al. 2013; Bendary et al. 2013). The current study shows that the inclusion of red seaweed by-product-based formulations at 2.5% in the concentrate mixture had no adverse effect on the serum biochemical profile of growing crossbred calves. Similarly, Abdoun et al. (2014) found no difference in serum glucose, total protein, and cholesterol with the supplementation of U. lactuca in the diet of growing lambs. In the same study, they found that plasma urea nitrogen concentrations were not affected by the seaweed extract inclusion. Another study with Ulva inclusion in fish (dusky kob), showed no significant difference in serum total protein, glucose, creatinine, albumin, globulin, blood triglycerides, and alanine aminotransferase among the groups. All the blood parameters in this present trial were similar to the past reports (Madibana et al. 2017). Furthermore, no significant variation was detected in albumin, ALT, and serum calcium level during 30th and 60th days when U. lactuca was added to the diet of chickens at 3% (Alagan et al. 2020). Nevertheless, the concentrations of total protein, globulin, serum phosphorus, and electrolytes such as Na, K, and Cl did not reveal any significant variations during the 30th day except where it was highly significant during the 60th day of the experimental period. In addition, Munde (2018) conducted an experiment on healthy crossbred calves and found no effect of supplementing seaweed by-products on blood serum AST, ALT, and ALP concentrations. These findings contradicted the findings of Alagan et al. (2020), who found a significant change in creatinine, AST, glucose, triglycerides, uric acid, and magnesium, as well as a significant difference in serum cholesterol concentration between the control (no supplementation) and the 3% seaweed, supplemented groups during the 30th and 60th days of the feeding trial. Bendary et al. (2013) also reported significantly higher variations in blood serum concentrations of total protein and globulin for premix and seaweed treatments than the control group.
Serum endocrine variables concentration
Serum levels of endocrine variables like T3 and T4 were similar between the different groups which prove that the inclusion of seaweed-based formulations had no adverse effect on the metabolic activity of crossbred calves. However, T4 increased with the period effect. Iodine is a vital precursor for the synthesis of the hormones T3 and T4, which are produced in the thyroid glands. Despite seaweed being a rich source of iodine, serum concentrations of thyroid hormones were not influenced by seaweed supplementation. The current findings are consistent with Hong et al. (2015), who found that brown seaweed by-products in the diet of Holstein cows had no effect on T3 and T4 concentrations. Likewise, Antaya (2016) reported that the inclusion of A. nodosum did not show any significant effect on serum levels of T3 and T4. In general, the results of this trial are in line with some past studies which reported that T3 and T4 was not significantly influenced by feeding seaweeds (Rasouli et al. 2004; Antaya et al. 2015; Maheswari et al. 2021). In case of cortisol, there was no significant (p > 0.05) variation observed among the calves from different treatment groups during the 180th experimental period. These outcomes are consistent with those of Kannan et al. (2007), who reported that seaweed supplementation did not affect goats' plasma cortisol concentrations. This present study is also in agreement with Galipalli et al. (2004) who found that when Boer goats were fed A. nodosum extract during and after 6 h of transportation plasma cortisol concentrations did not differ. Cortisol is often used as a marker to evaluate stress in domestic and wild mammals (Romero 2004). Increased plasma cortisol levels in goats are a sign of increased stress and have an adverse effect on their immune systems (Kannan et al. 2000). Higher levels of cortisol observed in animal trials may have been attributed to metabolic stress, and nutritional and management changes. Serum concentrations of cortisol were similar between the groups, and no significant impact of either treatment or period was recorded. Contrary to the current findings, previous studies reported that the inclusion of both Kappaphycus and Gracilaria in cattle tended to decrease the serum concentration of cortisol, however, a significant response was observed when both the seaweeds were combined (Munde 2018; Maheswari et al. 2021).
Antioxidant status
In this present trial, SOD, CAT, and glutathione peroxidase (GHS-Px) were measured as markers of antioxidant status. These markers have extensively been used to assess antioxidant status in animals (Fukai and Ushio-fukai 2011). The levels of SOD, CAT, and GHS-Px enzyme activity were similar among all the treatment groups. Our study findings disagree with Saker et al. (2004) who observed that including a brown seaweed (A. nodosum) by-product (Tasco) at 20% (v/w, as-fed) increased SOD activity in steers grazed on endophyte-infested tall fescue pasture. Similarly, the same authors reported that seaweed extract inclusion increased SOD activity in lambs exposed to prolonged heat stress for 10 days. Angulo et al. (2020) found a higher level of catalase activity in the rumen, liver, and intestinal mucus of goat kids in the groups supplemented with Sargassum at 2.5 and 5% compared with the control (no supplementation) for 70 days. Seaweeds contain antioxidants and are a rich source of sulphated polysaccharides, carrageenan, that are characterized by one or more -OH and -SO3H groups that render them the free radical scavenging capacity (Coelho et al. 1997). The antioxidant SOD is an intercellular antioxidant which feeds on superoxide that is produced in the cells as part of basic cellular metabolism (Saker et al. 2004). The differences in these reports can be linked to differences in seaweed type, the level of inclusion as supplements, the duration of the study and the method of processing the seaweeds. The method of processing may affect the rich phenolics contents of seaweeds. Thus, there may be a tendency for an enhanced level and activity of SOD in the calves which needs to be explored using different processing methods of the red seaweeds, increasing the proportion used in this study and also changing the experimental period.
Immune parameter
Serum immunoglobulin G level in the seaweed-added groups was significantly (p < 0.05) higher than that of the control group. As well, a time effect was detected with the advancement of the calves' age. It means that the inclusion of red seaweeds in the diet may increase IgG antibody concentration in crossbred calves. The present findings are in agreement with Hwang et al. (2014) who found that due to the inclusion of seaweed in Hanwoo steers, the IgG concentrations were improved significantly. In general, inclusion of seaweed resulted in increased concentration of IgG (Allen and Pond's 2002; Leonard et al. 2010). In addition, both K. alvarezii and G. salicornia are rich sources of vitamin C, carotenoids and vitamin E (Matanjun et al. 2010). Therefore, the increased immune response due to the inclusion of red seaweed could be connected to combination of bioactive compounds. On the contrary to our current experiment, Samarasinghe et al. (2021) stated that due to supplementation of U. lactuca, A. nodosum or Saccharina latissima in preweaning dairy calves, the plasma concentration of IgG was not varied but the innate immune response was increased significantly during the trial period. Novoa-Garrido et al. (2014) reported a decrease in serum IgG concentration in pregnant ewes when A. nodosum was supplemented at 546 g kg−1 diet. In brief, inclusion of tropical red seaweed improved immune response of growing crossbred calves.
Conclusion
The results of this experiment suggest that dietary supplementation with tropical red seaweed by-products at 2.5% in the concentrate mixture could positively improve immune response without any alteration in hormonal profile, serum metabolites, liver enzymes, endocrine variables, and growth performance in crossbred calves.
Data availability
All data generated or analysed in the present study are available upon reasonable request.
References
Abd Ellah MR, Hamed I, Ibrahim DR, Rateb HZ (2013) Reference values for haematological and serum biochemical constituents in late pregnant buffaloes. Assiut Vet Med J 59:35–46
Abdoun KA, Okab AB, El-Waziry AM, Samara EM, Al-Haidary AA (2014) Dietary supplementation of seaweed (Ulva lactuca) to alleviate the impact of heat stress in growing lambs. Pak Vet J 34:108–111
Aebi H (1984) Catalase in vitro. Methenzymol 105:121–126
Ahmad F, Sulaiman MR, Saimon W, Yee CF, Matanjun P (2012) Proximate compositions and total phenolic contents of selected edible seaweed from Semporna, Sabah, Malaysia. Borneo Sci 31:85–96
Alagan TV, Vatsala NR, Sagadevan I, Subbiah V, Ragothaman V (2020) Effect of dietary supplementation of seaweed (Ulva lactuca) and Azolla on growth performance, haematological and serum biochemical parameters of Aseel chicken. Beni-Suef Univ J Basic Appl Sci 9:1–9
Allen VG, Pond KR (2002) Seaweed Supplement Diet for Enhancing Immune Response in Mammals and Poultry. Texas Tech University, Lubbock
Anderson MJ, Blanton JR Jr, Gleghorn J, Kim SW, Johnson JW (2006) Ascophyllum nodosum supplementation strategies that improve overall carcass merit of implanted English crossbred cattle. Asian-Australas J Anim Sci 19:1514–1518
Angulo C, Chavez-Infante L, Reyes-Becerril M, Angulo M, Romero-Geraldo R, Llinas-Cervantes X, Cepeda-Palacios R (2020) Immunostimulatory and antioxidant effects of supplemental feeding with macroalga Sargassum spp. on goat kids. Trop Anim Hlth Prod 52:2023–2033
Antaya NT, Soder KJ, Kraft J, Whitehouse NL, Guindon NE, Erickson PS, Brito AF (2015) Incremental amounts of Ascophyllum nodosum meal do not improve animal performance but do increase milk iodine output in early lactation dairy cows fed high-forage diets. J Dairy Sci 98:1991–2004
Antaya N (2016) Kelp meal (Ascophyllum nodosum) supplementation to organic lactating dairy cows: effects on milk production, milk composition, animal health and nutrient utilization during the non-grazing and grazing seasons in New Hampshire. Doctoral dissertation, University of New Hampshire
AOAC (2005) Official Methods of Analysis, 18th edn. Association of Official Analytical Chemists, Washington, DC
Archer GS, Friend TH, Caldwell D, Ameiss K, Krawczel PD (2007) Effect of the seaweed Ascophyllum nodosum on lambs during forced walking and transport. J Anim Sci 85:225–232
Bendary MM, Bassiouni MI, Ali MF, Gaafar HM, Shamas AS (2013) Effect of premix and seaweed additives on productive performance of lactating Friesian cows. Int Res J Agric Sci Soil Sci 3:174–181
Cabrita AR, Correia A, Rodrigues AR, Cortez PP, Vilanova M, Fonseca AJ (2017) Assessing in vivo digestibility and effects on immune system of sheep fed alfalfa hay supplemented with a fixed amount of Ulva rigida and Gracilaria vermiculophylla. J Appl Phycol 29:1057–1067
Carrillo S, López E, Casas MM, Avila E, Castillo RM, Carranco ME, Pérez-Gil F (2008) Potential use of seaweeds in the laying hen ration to improve the quality of n-3 fatty acid enriched eggs. J Appl Phycol 20:721–728
Choi YY, Shin NH, Lee SJ, Lee YJ, Kim HS, Eom JS, Lee SS, Kim ET, Lee SS (2021) In vitro five brown algae extracts for efficiency of ruminal fermentation and methane yield. J Appl Phycol 33:1253–1262
Chojnacka K, Saeid A, Witkowska Z, Tuhy L (2012) Biologically active compounds in seaweed extracts-the prospects for the application. The Open Conf Proc J 3:20–28
Chopin T, Sawhney M (2009) Seaweeds and their mariculture. In: Steele JH, Thorpe SA, Turekian KK (eds) Encyclopedia of Ocean Sciences. Elsevier, Oxford, pp 4477–4487
Rishabh Chugh (2020) Maternal supplementation of seaweed by-products to improve performance in neonatal murrah buffalo calves. MSc Thesis, Animal Nutrition Division, ICAR-National Dairy Research Institute, Karnal-Haryana, India
Coelho RW, Fike JH, Schmidt RE Zhang X, Allen VG Fontenot JP (1997) Influence of seaweed extract on growth, chemical composition , and superoxide dismutase activity in tall fescue. In: Proceedings of the 1997 American Forage and Grassland Council. Vol. 6. American Forage and Grassland Council. Georgetown, TX. pp. 163–167
Diharmia A, Fardiazb D, Andarwulan N (2019) Chemical and minerals composition of dried seaweed eucheuma spinosum collected from indonesia coastal sea regions. Int J Ocean Oceanogr 13:65–71
El-Din AM, Gaafar HMA, El-Nahas HM, Ragheb EE, Mehrez AF (2008) Effect of natural feed additives on performance of growing Friesian calves. Egyptian J Anim Prod 45:401–413
El-Waziry A, Al-Haidary A, Okab A, Samara E, Abdoun K (2015) Effect of dietary seaweed (Ulva lactuca) supplementation on growth performance of sheep and on in vitro gas production kinetics. Turk J Vet Anim Sci 39:81–86
FAO and IFIF (2010) Good practices for the feed industry – implementing the Codex Alimentarius code of practice on good animal feeding. FAO Animal Production and Health Manual No. 9. Rome
FAO (2013) Global aquaculture production statistics for the year 2011. Fisheries and aquaculture department. Available from: ftp://ftp.fao.org/FI/news/GlobalAquacultureProductionStatistics2011.pdf. Accessed 30 May 2014
Fukai T, Ushio-Fukai M (2011) Superoxide dismutases: role in redox signaling, vascular function, and diseases. Antiox Redox Signal 15:1583–1606
Galipalli S, Gadiyaram KM, Kouakou B, Terrill TH, Kannan G (2004) Physiological responses to preslaughter transportation stress in Tasco-supplemented Boer goats. S Afric J Anim Sci 34:198–200
Ganesan P, Kumar CS, Bhaskar N (2008) Antioxidant properties of methanol extract and its solvent fractions obtained from selected Indian red seaweeds. Bioresour Technol 99:2717–2723
Gardiner GE, Campbell AJ, O’Doherty JV, Pierce E, Lynch PB, Leonard FC, Lawlor PG (2008) Effect of Ascophyllum nodosum extract on growth performance, digestibility, carcass characteristics and selected intestinal microflora populations of grower–finisher pigs. Anim Feed Sci Technol 141:259–273
Hong ZS, Kim EJ, Jin YC, Lee JS, Choi YJ, Lee HG (2015) Effects of supplementing brown seaweed by-products in the diet of Holstein cows during transition on ruminal fermentation, growth performance and endocrine responses. Asian-Australasian J Anim Sci 28:1296
Hopkins DL, Clayton EH, Lamb TA, Van de Ven RJ, Refshauge G, Kerr MJ, Ponnampalam EN (2014) The impact of supplementing lambs with algae on growth, meat traits and oxidative status. Meat Sci 98:135–141
Hwang JA, Islam MM, Ahmed ST, Mun HS, Kim GM, Kim YJ, Yang CJ (2014) Seamustard (Undaria pinnatifida) improves growth, immunity, fatty acid profile and reduces cholesterol in Hanwoo steers. Asian-Australasian J Anim Sci 27:1114
ICAR (2013) Nutrient Requirements of cattle and buffaloes, 3rd edn, Indian Council of Agriculture Research, New Delhi
Kaneko S, Feinstone SM, Miller RH (1989). Rapid and sensitive method for the detection of serum hepatitis B virus DNA using the polymerase chain reaction technique. J Clin Microbiol 27:1930-1933.
Kahn CM, Line S (2010) The Merck Veterinary Manual. Merck, Whitehouse Station
Kannan G, Terrill TH, Kouakou B, Gazal OS, Gelaye S, Amoah EA, Samake S (2000) Transportation of goats: effects on physiological stress responses and live weight loss. J Anim Sci 78:1450–1457
Kannan G, Terrill TH, Kouakou B, Galipalli S (2007) Blood metabolite changes and live weight loss following brown seaweed extract supplementation in goats subjected to stress. Small Rumin Res 73:228–234
Karatzia M, Christaki E, Bonos E, Karatzias C, Florou-Paneri P (2012) The influence of dietary Ascophyllum nodosum on haematologic parameters of dairy cows. Ital J Anim Sci 11:e31
Kinley RD, Martinez-Fernandez G, Matthews MK, de Nys R, Magnusson M, Tomkins NW (2020) Mitigating the carbon footprint and improving productivity of ruminant livestock agriculture using a red seaweed. J Cleaner Prod 259:120836
Lee HG, Hong ZS, Li ZH, Xu CX, Jin X, Jin MG, Choi YJ (2005) Effect of brown seaweed waste supplementation on lactational performance and endocrine physiology in Holstein lactating cows. J Anim Sci Technol 47:573–582
Leonard SG, Sweeney T, Bahar B, Lynch BP, O’doherty JV, (2010) Effect of maternal fish oil and seaweed extract supplementation on colostrum and milk composition, humoral immune response, and performance of suckled piglets. J Anim Sci 88:2988–2997
Leupp JL, Caton JS, Soto-Navarro SA, Lardy GP (2005) Effects of cooked molasses blocks and fermentation extract or brown seaweed meal inclusion on intake, digestion, and microbial efficiency in steers fed low-quality hay. J Anim Sci 83:2938–2945
Madesh M, Balasubramanian KA (1998) Microtiter plate assay for superoxide dismutase using MTT reduction by superoxide. Indian J Biochem Biophys 35:184–188
Madibana MJ, Mlambo V, Lewis B, Fouché C (2017) Effect of graded levels of dietary seaweed (Ulva sp.) on growth, hematological and serum biochemical parameters in dusky kob, Argyrosomus japonicus, sciaenidae. Egypt J Aquat Res 43:249–254
Maheswari M, Das A, Datta M, Tyagi AK (2021) Supplementation of tropical seaweed-based formulations improves antioxidant status, immunity and milk production in lactating Murrah buffaloes. J Appl Phycol 33:2629–2643
Matanjun P, Mohamed S, Muhammad K, Mustapha NM (2010) Comparison of cardiovascular protective effects of tropical seaweeds, Kappaphycus alvarezii, Caulerpa leniillifera, and Sargassum polycystum on high cholesterol/high-fat diet in rats. J Med Food 13:792–800
Melton CSM (2001) Influence of coated whole cottonseed on nutritional value and its potential for supplementation to ruminants. PhD Thesis, Texas Tech University
Munde VK (2018) Evaluation of red seaweed meal as ruminant feed supplement. Doctoral Thesis, ICAR-Indian Veterinary Research Institute (Deemed University) Izatnagar
Ndlovu T, Chimonyo M, Okoh AI, Muchenje V, Dzama K, Raats JG (2007) Assessing the nutritional status of beef cattle: current practices and future prospects. Afr J Biotech 6:2727–2734
Novoa-Garrido M, Aanensen L, Lind V, Larsen HJS, Jensen SK, Govasmark E, Steinshamn H (2014) Immunological effects of feeding macroalgae and various vitamin E supplements in Norwegian white sheep-ewes and their offspring. Livestock Sci 167:126–136
O’Doherty JV, Dillon S, Figat S, Callan JJ, Sweeney T (2010) The effects of lactose inclusion and seaweed extract derived from Laminaria spp. on performance, digestibility of diet components and microbial populations in newly weaned pigs. Anim Feed Sci Technol 157:173–180
Pickard RM, Beard AP, Seal CJ, Edwards SA (2008) Neonatal lamb vigour is improved by feeding docosahexaenoic acid in the form of algal biomass during late gestation. Animal 2:1186–1192
Rajauria G (2015) Seaweeds: a sustainable feed source for livestock and aquaculture. In: Tiwari BK, Troy DJ (eds) Seaweed sustainability. Elsevier, Amsterdam, pp 389–420
Rasouli A, Nouri M, Khajeh GH, Rasekh A (2004) The influences of seasonal variations on thyroid activity and some biochemical parameters of cattle. Sci Info Datab 5:55–62
Rjiba-Ktita S, Chermiti A, Valdés C, López S (2019) Digestibility, nitrogen balance and weight gain in sheep fed with diets supplemented with different seaweeds. J Appl Phycol 31:3255–3263
Romero LM (2004). Physiological stress in ecology: lessons from biomedical research. Trends Ecol Evol 19:249–255
Samarasinghe MB, Sehested J, Weisbjerg MR, Vestergaard M, Hernández-Castellano LE (2021). Milk supplemented with dried seaweed affects the systemic innate immune response in preweaning dairy calves. J Dairy Sci 104:3575–3584
Saker KE, Fike JH, Veit H, Ward DL (2004) Brown seaweed-(TascoTM) treated conserved forage enhances antioxidant status and immune function in heat-stressed wether lambs. J Anim Physiol Anim Nutr 88:122–130
Singh RP, Kumari P, Reddy CRK (2015) Antimicrobial compounds from seaweeds-associated bacteria and fungi. Appl Microbiol Ecol 99:1571–1586
Sharma A, Datt C (2020) Supplementation effect of red seaweed power on dry matter intake, body weight and feed conversion efficiency in crossbred cows. J Entomol Zool Stud 8:1056–1059
Singh BK, Chopra RC, Rai SN, Verma MP, Mohanta RK (2016) Effect of feeding seaweed as mineral source on mineral metabolism, blood and milk mineral profile in cows. Proc Nat Acad Sci India B 86:89–95
Singh BK, Chopra RC, Rai SN, Verma MP, Mohanta RK (2017) Nutritional evaluation of seaweed on nutrient digestibility, nitrogen balance, milk production and composition in Sahiwal cows. Proc Nat Acad Sci India B 87:437–443
Singh M, Kumar S, Banakar PS, Vinay VV, Das A, Tyagi N, Tyagi AK (2021) Synbiotic formulation of Cichorium intybus root powder with Lactobacillus acidophilus NCDC15 and Lactobacillus reuteri BFE7 improves growth performance in Murrah buffalo calves via altering selective gut health indices. Trop Anim Hlth Prod 53:1–9
Van Soest PV, Robertson JB, Lewis BA (1991) Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J Dairy Sci 74:3583–3597
Wang Y, Xu Z, Bach SJ, McAllister TA (2008) Effects of phlorotannins from Ascophyllum nodosum (brown seaweed) on in vitro ruminal digestion of mixed forage or barley grain. Anim Feed Sci Technol 145:375–395
Zhou M, Hünerberg M, Chen Y, Reuter T, McAllister TA, Evans F, Guan LL (2018) Air-dried brown seaweed, Ascophyllum nodosum, alters the rumen microbiome in a manner that changes rumen fermentation profiles and lowers the prevalence of foodborne pathogens. Msphere 3:e00017–18
Acknowledgements
The authors thank the Director of ICAR-National Dairy Research Institute, Karnal, India for providing all the necessary support and facilities to conduct the present research.
Author information
Authors and Affiliations
Contributions
P.A. conducted the feed trials, laboratory work, haemato-biochemical analysis, data collection, and manuscript writing-review and editing. R.M. designed, outlined, and drafted the manuscript of the study. L.O. worked out the statistical analysis of data and helped in making tables. B.AM. language and critical revision of the manuscript and necessary suggestions for manuscript improvement. H.B.N. carried out the collection of seaweeds and formulation of the feed. All authors have approved the manuscript for submission.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Conflict of interest
The authors declare that there is no personal or professional conflict of interest.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Anderson, P., Malik, R., Ojha, L. et al. Investigations on modulating effect of three tropical red seaweed by-products on growth performance, immune response, antioxidant status and endocrine variables in crossbred calves. J Appl Phycol 35, 445–457 (2023). https://doi.org/10.1007/s10811-022-02871-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10811-022-02871-3