
Abstract
In the Bengal Delta Plains (BDP) of South Asia, there is an increased report of bioaccumulation of arsenic (As) in rice grains and plants which can ultimately result in health hazards in human population consuming rice as a primary staple food. Five abundant cyanobacteria were isolated from the rice fields of BDP and maintained in vitro. The characterized isolates resembled Leptolyngbya sp. (isolate LBK), Nostoc spp. (isolates NOC and NOK) and Westiellopsis spp. (isolates WEC and WEK) based on polyphasic taxonomy. All the five isolates were assessed for biotransformation potential of As vis-à-vis adaptability and survivality under different levels of arsenite compared to control set of experiments. Adaptive changes of cyanobacterial photosynthetic pigments in terms of autofluorescence emission along with nitrogenase activity and exopolysaccharide production were measured for all isolates. The inorganic As absorption in terms of bioconcentration factor (BCF) in dry biomass was found to be highest in NOC (0.201–0.220), followed by NOK (0.147–0.150), WEK (0.071–0.074), WEC (0.051) and LBK (0.014) when exposed in presence of higher (200–400 μM) to lower (100 μM) arsenite concentrations respectively for 7 days. The transformation of arsenite to relatively less toxic arsenate was detected in varying efficiency in all the studied isolates. When treated with 100–400 μM arsenite, 9.58–78.4 % arsenate was detected in growth medium whereas 33–100 % in dry biomass of cyanobacterial isolates. The cyanobacterial isolates of this study could be potentially applied to reduce bioavailability of As in rice fields of South Asia based on further field trials, thereby ultimately rendering rice grains safe for human consumption.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Arsenic (As) is present in the earth crust as a natural element in varying concentration since the origin of this planet (Nordstrom 2002). This metalloid is ranked first as it is considered to be highly hazardous to various organisms including humans (Vaughan 2006). In recent decades, the environmental fate and behaviour of As has received increased attention due to ongoing crisis in parts of South Asia and South-East Asia (Nickson et al. 2000; Bhattacharya et al. 2001). Millions of people in this part of the Asian continent have been exposed to high levels of As due to intake of As-contaminated drinking water (Smedley and Kinniburgh 2002). Bioaccumulation of As has been also reported in rice grains and plants (Williams et al. 2007). This can ultimately affect human populations consuming rice as a primary staple food (Zhu et al. 2008; Meharg et al. 2009). It is now known that the use of As-contaminated ground water for irrigation is the main source behind As bioaccumulation in rice (Zhao et al. 2013; Newbigging et al. 2015). The average contribution to total As intake in human population from drinking water has been reported to be 13 % (Ohno et al. 2007) whereas from cooked rice it is 56 % (Zheng et al. 2013), thus clearly highlighting that rice contributes most to daily As intake in this part of the world.
Globally, rice is normally cultivated in flooded agricultural soil. The flooded rice soil ecosystem is characterized by aerobic and anaerobic zones represented in three major ecological layers (Venkataraman 1993). Inorganic As species such as arsenite [As(III)] or arsenate [As(V)] are generally more abundant and toxic than organoarsenic (methyl arsenic) species in the environment (Rossman 2003). The As(III) is most prevalent under reduced conditions (Hasegawa et al. 2010) as observed in flooded rice fields (Boivin et al. 2002; Gao et al 2002). Arsenite is more mobile than As(V) due to its neutral nature and remains loosely adsorbed to soil particles (Lakshmipathiraj et al. 2006). As(V) is found to be more stable in oxic environments (Zhang et al. 2015). Thus As(III) is more bio-available and get transported to the rice plant and ultimately bioaccumulates in rice grains (Zhang et al. 2015).
Microorganisms undertake As cycling in terrestrial ecosystem by oxidation, reduction, methylation and volatilization (Oremland and Stolz 2003; Qin et al. 2006). Oxidation of As(III) to less toxic As(V) is thought to be the most important detoxification mechanism which ultimately leads to methylation (Rahman and Hassler 2014). Oxidation of As(III) by different organisms involves a key enzyme arsenite oxidase. Over the last few decades, numerous studies have described different responses in microorganisms exposed to this metalloid including induction of transport system (Thiel 1988), expression of resistance genes (Jackson and Dugas 2003; López-Maury et al. 2003), phytochelatin production (Pawlik-Skowrońska et al. 2004) and de novo synthesis of thiol and related compounds (Morelli et al. 2005).
In the rice field ecosystem of tropical countries, cyanobacteria are one of the indigenous nitrogen-fixing soil-borne oxygenic photosynthetic prokaryotic microorganisms abundantly present in soil-water interphase and act as natural bio-fertilizers. In this specialized niche, cyanobacteria play a critical role in As transformation, mobilization and transport from soil-water interphase to rice plants (Zhang et al. 2015). Cyanobacteria can take up and concentrate As from the surrounding environment and As-resistant genera such as Synechocystis, Phormidium, Oscillatoria, Nostoc and Anabaena have been reported as dominant taxa from different As-contaminated areas globally, including rice field environments (Maeda et al. 1998; Shaheen et al. 2007; Banerjee 2008; Yin et al. 2011). Some studies have used natural algal populations from fresh water ecosystems and exposed them to As(V) and these algae showed promising biotransformation potential (Pawlik-Skowrónska et al. 2004; Bahar et al. 2013; Wang et al. 2013). Only a handful of studies have been undertaken with freshwater cyanobacteria which were exposed to As(III) and showed ability to transform into As(V) followed by methylation (Yin et al. 2011; Wang et al. 2013).
The Bengal Delta Plain (BDP) located in South Asia and formed by the Gangetic alluvial deposits have characteristic topography, geomorphological and geological features. The BDP is drained by three of the major rivers of the world—the Ganges, Brahmaputra and Meghna (also known as the GBM Delta), which originate from the Himalayas. In West Bengal, India, a part of the BDP has been formed by elevated alluvial terraces of the Barind and Madhupur tracts of Pleistocene age overlaid by Holocene flood plain deposits (Alam et al. 1990). The nine As-affected districts in West Bengal have an area of 38,865 km2 and a population of 42.7 million. Around six million people living in this region presently consume As-contaminated water with As level of 50 μg L−1. It has been estimated around 300,000 people may be suffering from As-related diseases in the BDP region (Rahman et al. 2001). The source of As in groundwater of this deltaic plain is considered to be geogenic in origin and largely restricted to the Holocene aquifer sediments (Bhattacharya et al. 1997; BGS and DPHE 2001). Several models have been put forward to explain the source and mobilization mechanism of As in groundwater of the BDP, an area of international interest (Nath et al. 2011). Large areas of the BDP are home to rice fields and rice cultivation is entirely dependent on contaminated ground water for the purpose of irrigation which ultimately results in bioaccumulation of As in rice grains and plants (Mondal and Polya 2008a, b; Chatterjee et al. 2010). The rice field ecosystems are home to indigenous microbial communities including cyanobacteria which may have the potential to biotransform different species of As. However, almost no study has been undertaken to date to look at the biotransformation potential of indigenous cyanobacteria present in As-contaminated rice field ecosystems of BDP in the presence of As(III) under in vitro conditions.
The objectives of the present study were the identification of the five most abundant indigenous cyanobacteria isolated from As-contaminated rice fields of BDP using polyphasic taxonomy and subsequent evaluation of their adaptive responses, tolerance, absorption and biotransformation potential in the presence of arsenite (sodium arsenite) in vitro.
Materials and methods
Study area
The district of Nadia in West Bengal, India, is situated in the alluvial zone of BDP and designated as one of the As-affected ‘hot spots’ nationally and globally (WHO 2011; Chowdhury et al. 2000; Bhattacharya et al. 2001). In the present study, soil samples were collected from two highly As-affected blocks from Nadia, namely Karimpur II (KMP) and Chakdaha (CHK). In both the sampling areas, As concentration is much higher in irrigation as well as drinking water exceeding the permissible limits of FAO (1985) and WHO (2011). One shallow tube well-irrigated rice field from each blocks namely Barbakpur of Karimpur II (KMP: N 23° 55.064′, E 88° 33.350′) and Gontera of Chakdaha (CHK: N 23°41.86′, E 87°40.66′), were selected for sampling activities. The Karimpur II site is routinely monitored by our laboratory to study the role of microbial communities in arsenic biogeochemical cycling (Ghosh et al. 2014, 2015). The other site, CHK, was chosen since it has numerous rice fields with very high concentration of As in the soil (Shaheen et al. 2007; Chatterjee et al. 2010; Nath et al. 2011).
Sample collection and storage
Soil cores from each site were collected between April and June, 2014. Soil cores of 15 cm in length were collected using a soil auger (6 inches; locally made). Soil from the upper crust (10 mm thick) was used for isolation of cyanobacteria. Soil samples representing a depth of up to 6 inches (excluding upper crust) were separated and air-dried. Subsequently, air-dried soil samples were ground and passed through sieve and homogenized to make a representative sample for subsequent nutrient analyses.
Soil physico-chemical analysis
In situ soil temperature was measured in the field using a thermometer. Soil pH was determined by a Sartorius digital pH meter and electrical conductivity (EC) was measured using a Systronics conductivity bridge at 25 °C. Available soil nitrogen was determined by alkaline-KMnO4 method (Subbaiah and Asija 1956) which primarily measures easily oxidizable nitrogen in a Kjeltec Auto 1030 Analyzer (Tecator, Sweden). Available phosphorus content was estimated based on a previously published methodology (Bary and Kurtz 1945). The percentage of organic carbon (OrC) was estimated following Walkley and Black (1934). For total As determination, digestion of soil samples was undertaken following Bhattacharya et al. (2009) and samples were subsequently analysed by inductively coupled plasma emission mass spectrometer (ICP-MS) (Q-ICP-MS, X Series 2, Thermo Scientific, USA).
Isolation of cyanobacteria
Ten gram of finely ground soil was put into 100 mL of distilled water, stirred and allowed to settle. The supernatant was subsequently used as inoculum. Agar plates (1.2 % agar) containing nitrogen-free BG-11 medium (Stanier et al. 1971) were used as growth medium. One mL of soil suspension was put in each plate. Subsequently, agar plates were kept at 25 ± 2 °C in a culture rack receiving 25–30 μmol photons m−2 s−1 from a cool fluorescent lamp under 20:4-h light/dark (LD) cycle. Visible blue green or brown colonies appeared within 7–10 days of inoculation which were characterized using a stereo zoom microscope. Colonies which were 15–20 days old were subsequently used to establish liquid cultures.
Cyanobacterial isolates and culture condition
Pure cultures were maintained in liquid BG11 medium with or without combined nitrogen at 25 ± 2 °C and 25–30 μmol photons m−2 s−1 with a 20:04-h L:D cycle. All five isolates are presently maintained in algal culture collection in the Department of Biological Sciences, Indian Institute of Science Education and Research Kolkata (IISERK), India.
Morphotaxonomy of isolates
Morphological features of isolates grown in culture medium were studied under a light microscope (LM) enabled with a camera and differential interference contrast (DIC) images were taken using CellSens software (BX53, Olympus, Japan). Prior to field emission scanning electron microscopy (FESEM), cyanobacterial biomass was fixed in 2.5 % glutaraldehyde [in 100 mM phosphate buffered saline (PBS), pH-7.4] and left overnight followed by washing with PBS. Dehydration was performed in graded ethanol series (40–100 %). Both treated and untreated samples were drop coated on a 1-cm glass piece and air-dried. The samples were coated by gold-palladium sputtering (Quorom Technologies Ltd., UK) and subsequently imaged by FESEM (SUPRA 55VP, Carl Zeiss, Germany). The cyanobacterial isolates were identified based on available morpho-taxonomic keys (Desikachary 1959; Anagnostidis and Komárek 1990; Komárek and Anagnostidis 2005; Komárek et al. 2014).
Molecular phylogeny
Genomic DNA was extracted from 2 mL of cyanobacterial culture representing each isolate using standard method of Böstrom et al. (2004). The yield of extracted genomic DNA was quantified by Nano Drop (Thermo Scientific, USA) prior to PCR amplification. Partial 16S rRNA fragments were amplified from the extracted genomic DNA using cyanobacterial specific forward primer CyaF581 and reverse primer CyaR631 (Mühling et al. 2008). PCR reaction conditions were as per Mühling et al. (2008). The generated PCR products were purified using Qiagen PCR purification kit as per manufacturer’s instructions. The purified PCR products were sequenced in an ABI 3730 Genetic Analyzer based on Big Dye Terminator chemistry. Generated chromatograms were checked in BioEdit v7.0 (Hall 1999) for any ambiguity before further analysis. Generated sequences were compared with published cyanobacterial 16S rRNA sequences (GenBank/RDB/EMBL databases) using Blastn tool. The alignment, which included the 16S rRNA sequence (approx. 298 bp) of each isolate generated from this study and published 16S rRNA sequences of representative cultured cyanobacterial strains (GenBank), was generated using Clustal Omega (http://www.ebi.ac.uk/Tools/msa/clustalO) and subsequently checked in SEAVIEW. A phylogenetic tree based on Maximum Likelihood method (ML) with GTR model and gamma distribution was constructed in MEGA v6.0 (Tamura et al. 2013). The ML tree was subsequently bootstrapped in MEGA v6.0 (1000 replicates) to generate consensus tree retaining significant branching of >45 %. The generated partial 16S rRNA sequences representing the five isolates have been submitted to GenBank and their accession numbers are KT361166 (NOC isolate), KT361167 (WEC isolate), KT361168 (NOK isolate) and KT361169 (LBK isolate) KT361170 (WEK isolate).
Determination of inorganic arsenic [As(III)] tolerance and inhibition by cyanobacterial isolates
The inorganic salt of sodium arsenite (NaAsO2; Sd Chemicals, India) was used to test cyanobacterial isolates under various concentration (0, 10, 50, 100, 200, 500 and 1000 μM). Medium used to grow each isolate but without sodium arsenite was designated as control. The sodium arsenite solution was filter sterilized and spiked aseptically to each set up of autoclaved culture medium before inoculation. Arsenite tolerance and inhibition in each isolate were determined and expressed in terms of representative concentration of As(III), viz., no observed effective change (NOEC, 10–100 μM); effective concentration 50 (EC50; growth reduces by 50 %, >200–1000 μM) and concentration lower than EC50 or sub-effective concentration (SEC50, growth present above 50 %, 200–400 μM) as compared to control, based on survivality response in terms of chlorophyll-a (Chl-a) content of 7-day-old cyanobacterial culture (Debnath et al. 2012). The NOEC and SEC50 were also calculated from the same growth experiments.
The absorbance of inoculum (homogenized biomass) during inoculation in the liquid medium containing sodium arsenite at 750 nm was 0.2. Inoculation was done aseptically and finally 100 mL of cultures were maintained at 25 ± 2 °C and 25–30 μmol photons m−2 s−1 under 20:04-h LD cycle conditions. All experiments were conducted under same incubation condition unless mentioned separately.
Physiological characterization
Growth, specific growth rate, generation time, pigment content, pigments relative autofluorescence signature, nitrogenase activity and extracellular polymeric substances (EPS) production were characterized for each isolate in this study. Biomass and growth of the isolates were determined in terms of Chl-a content using methods described by Mackinny (1941). The growth rate was calculated by using the formula k = ln (N 1 / N 0) / t of Guillard (1973), where N 1 and N 0 are Chl-a amount at the end and beginning of experiments in time t days. Specific growth rate (k) and generation time (24/k h) were calculated from Chl-a readings taken on every 3rd day up to 15 days.
Relative autofluorescence and emission spectra of pigments for each cyanobacterial isolate were determined using confocal laser scanning microscopy (CLSM). Twenty microlitre of the resultant sample was places on a glass slide and sealed with cover slip for observation compared to untreated control set. Filaments or cells were imaged with a Zeiss CLSM 710 with Axioobserver and Zen 2010 software (Carl Zeiss, Germany) using DIC optics with a 40×/1.3NA oil immersion Plan-Neofluar objective lens or 63×/1.4NA oil immersion Plan-Apo objective lens. DIC imaging was conducted with excitation by a 488-nm argon laser at 10 % power, using an NT 80/20 filter. Initial autofluorescence settings for chlorophyll [in Photosystem II, (PSII)] detection were based on settings adapted from previously published method (Sinha et al. 2002). To determine adaptability and survivality under As(III) stress in terms of Chl-a and phycobiliproteins (PBPs) such as phycocyanin (PC), phycoerythrin (PE) and allophycocyanin (APC) autofluorescence, spectral scanning was initiated for each isolate. Emission scans were collected in ~10.7 nm bandwidth with increments ranging from 565 to 724 nm.
Nitrogenase activity of a 7-day-old culture for each isolate was estimated based on an acetylene reduction assay (ARA) method following the principle of Turner and Gibson (1980) by gas chromatography (YL6000, Youngling, Korea). EPS extraction were done by separating cells from culture medium by centrifugation at 12,000 rpm for 30 min. Total carbohydrate content of the supernatant was measured following the protocol of Dubois et al (1956).
Arsenic absorption and transformation in cyanobacterial isolates
To investigate the As absorption and transformation in cyanobacterial isolates, sodium arsenite was used at the indicated concentration (NOEC and SEC50 of each isolate). Each set of cyanobacterial culture (in 100 mL medium) exposed to As(III) was incubated for 7 days, subsequently harvested, rinsed with de-ionized water and ice-cold phosphate buffer for 10 min to remove apoplastic As. For absorption study, total As was determined from cyanobacterial biomass and culture medium cyanobacterial biomass against a control set using ICP-MS (Q-ICP-MS, X Series 2, Thermo Scientific, USA) and reference standard of 23 element standard (ICP23A20, Reagecon, Ireland). Samples were prepared as mentioned below. The bioconcentration factor (BCF) was estimated to find out the bioabsorption potential for each cyanobacterial isolate. The ratio of total inorganic arsenic in cyanobacterial biomass [dry weight, (CBM)] to initial As concentration in growth medium (CIN) was used to calculate BCF following Bahar et al (2013).
To evaluate the biotransformation potential of each cyanobacterial isolate, fractions of inorganic As [As(III) and As(V) fractions] were determined. Approximately 0.02 g (dry weight) of cyanobacterial biomass was put in 50 mL centrifuge tubes and treated in 10 mL of 2 % HNO3 overnight. The samples were subsequently heated in a microwave based on published protocol (Zhu et al. 2008). The digested extract of each isolate and corresponding 1 % HNO3 acidified culture medium was filtered separately through 0.45 μm nitrocellulose filter disc (Rankem, India) and stored for absorption (ICP-MS) and transformation analysis. Inorganic As(III) and As(V) were separated using As speciation cartridge (Metal Soft Center, USA) which retains As(V) through alluminosilicate adsorbent. A Varion AA240 model atomic absorption spectrophotometer (AAS) equipped with vapour generation accessories (model no. VGA77) was used for estimation of total inorganic As and filtrate collected from speciation cartridge following the protocol of Datta et al. (2010) for undertaking transformation study. Reducing agent (Aquas solution of 0.6 % sodium borohydride in 0.5 % sodium hydroxide) and acid (40 % hydrochloric acid) were freshly prepared before use. For reliability of this method, all As samples were previously analysed by ICP-MS for total inorganic As and total As concentration was determined by AAS. A reference standard (ACCU Trace; AA03N-1, USA) and reagent blanks were used to maintain accuracy of the analysis.
Results
Physico-chemical properties of soil samples
The physico-chemical characteristics of soil samples collected from sites KMP (Karimpur II) and CHK (Chakdaha) are detailed in Table 1. Briefly, both the soils were slightly alkaline, low in electrical conductivity, moderate in available nitrogen with low available phosphate and organic carbon. Total inorganic As content of both the soils were found to be higher than recommended globally [up to 10 and 20 mg kg−1 in normal and agriculture soil, respectively; (Rahaman et al. 2013 and references therein)]. The soils are alluvial, sub humid and representative of megathermal tropical agro climatic zone of South Asia.
Organisms and culture conditions
Cyanobacterial colonies appeared within 7–10 days on the agar plates exhibiting various colour, size and shapes. The most abundant colonies were chosen for further study. Three isolates from KMP site and two isolates from CHK site were selected for establishment of pure liquid cultures. Mucilaginous round colony of olive green to brown in colour was most abundant followed by non-mucilaginous radiating blue green colour designated as NOC and WEC respectively isolated from rice field soil of CHK. From KMP soil sample, same colonies (as in CHK) appeared with maximum abundance and designated as NOK and WEK throughout the study. All these NOC, WEC, NOK and WEK were grown on BG11 agar plates without combined nitrogen. In agar plates with combined nitrogen, no significant colony appeared when inoculated with CHK soil, but very distinct irregular mat-like blue green colony appeared following inoculation with KMP soil. This particular isolate referred as LBK isolate was also selected for further study.
Morphotaxonomy of isolates
The five most abundant isolates of cyanobacteria obtained from both sites were chosen for this study. Isolates were assigned to three distinct filamentous genera namely unbranched nonheterocystous (isolate LBK), unbranched heterocystous (isolate NOC and NOK) and true branched heterocystous (isolate WEC and WEK) based on LM and FESEM (Table 2). Based on morphotaxonomy, LBK isolate resembled Phormidium exhibiting comparatively thinner (1–2 μm broad) and loosely entangled filaments in mat-like formation (Fig. 1a, b). The NOC and NOK isolates showed morphological properties with overlapping characteristics and resembled Nostoc ellipsosporium. In case of NOC isolate, the filament width ranges from 2 to 4 μm whereas for NOK the filament ranges from 2.5 to 3.5 μm (Fig. 1c, d). The remaining two isolates, WEK and WEC, showed typical cellular dimension and uniform morphology resembling Westiellopsis (Fig. 1e, f). Based on FESEM, finer morphological details including surface texture variation, cell dimension and constriction at cross wall were also noted for all the five isolates (Fig. 2a–f) (Table 2).

LM micrographs of five cyanobacterial isolates: a LBK filaments loosely arranged; b LBK trichome; c NOC with heterocyst; d NOK with heterocyst; e WEC branch with lateral heterocyst; f WEK branch with intercalary heterocyst (scale bars = 10 μm)

FESEM micrographs of five cyanobacterial isolates: a LBK filament; b LBK trichome; c NOC with intact and dividing cells; d NOK with intact and dividing cells; e WEC branch with short cell; f WEK branch with elongated cells (scale bars: a, b = 2 μm, c–f = 5 μm)
Molecular phylogeny of isolates
Partial 16S rRNA sequences (approx 298 bp) representing the five isolates showed 97–100 % identity with published 16S rRNA sequences of cultured filamentous cyanobacteria (Table S1). Five isolates identified by morphological attributes were subjected to molecular phylogenetic study to get a clear understanding of their taxonomic affiliation. The top Blastn result for each isolate has been provided in Table S1. Based on ML approach, five isolates showed strong taxonomic affiliation with 16S rRNA sequences of cultured cyanobacteria and represented across four clades with strong bootstrap support for some of them (Fig. 3). In clade A, the isolate LBK clustered with sequences belonging to the genus Leptolyngbya. The 16S rRNA sequence of isolate NOK was found to be clustered with cultured sequences of Nostoc and represented in clade B. In clade C, the isolate NOC was found to be clustered with 16S rRNA sequences of cultured cyanobacteria belonging to Nostoc commune, Nostoc punctiforme and Nostoc verrucosum. The remaining two isolates, WEK and WEC, were part of the clade D along with cultured 16S rRNA sequences of belonging to Westiellopsis, Fischerella and Hapalosiphon; however, both the isolate sequences diverged from the rest indicating that these two isolates are novel.

ML tree represents relationship among 16S rRNA sequences of five cyanobacterial isolates of this study (NOC, NOK, WEC, WEK and LBK) and related cultured cyanobacteria based on 16S rRNA sequences. The 16S rRNA sequence of Cyanobacterium aponium used as outgroup. The ML tree was bootstrapped with 1000 replicates to produce consensus tree retaining significant branching of >45 %
Growth rate, generation time and EC50 of cyanobacterial isolates exposed to As(III)
Effect of As(III) on five cyanobacterial isolates was tested using 10, 50, 100, 200, 400, 500 and 1000 μM (1.3, 6.6, 13, 26, 52, 65 and 130 mg L−1, respectively) sodium arsenite. After treatment with 10–100 μM As (NOEC), growth was found to be accelerated in four isolates as evident from generation time (67.2 to 79 h) except for LBK (95 h) (Fig. 4). However, when As concentration reached 200–500 μM, growth in all the tested isolates were found to be inhibited by 50–60 % (generation time, 124.2–178.5 h) (Fig. 4). From this growth experiments, we calculated EC50 and SEC50 of each cyanobacterial isolate. The EC50 of NOC, NOK, WEC, WEK and LBK isolates were found to be 62.7, 60.3, 47.5, 50.2 and 28.60 mg L−1, respectively. The SEC50 was found to be 52 mg L−1 (400 μM) for NOC and NOK, 39 mg L−1 (300 μM) for WEC and WEK, and 26 mg L−1 (200 μM) for LBK, respectively. In all cases, NOEC was found to be 13 mg L−1 (100 μM). Among the five isolates, NOC was most tolerant to As(III) toxicity followed by NOK, WEC, WEK and LBK.

Generation time (hour) in terms of chlorophyll-a of five cyanobacterial isolates exposed to different concentration of sodium arsenite in vitro; (±SD; n = 3)
Photosynthetic pigments profile under As(III) exposure
The fluorescence emission spectra acquired by CLSM were retrieved from vegetative cells present in As filaments treated (NOEC and SEC50) and untreated biomass representing each isolate. In vivo maximum wavelength (λmax) typical of four distinct pigments was detected in this study (Table 3). The PBPs are attached to the thylakoid membrane and transfer energy to Chl-a in the photosynthetic reaction centres. The changes in the PBPs that occur during As treatment consist of changes in the antenna molecule PBPs content and reaction centre (PSII) Chl-a. By nature, PBPs are also highly autofluorescent proteins. Three PBPs were found in the studied cyanobacterial isolates. PBPs exhibited distinct wavelengths of fluorescence emission maximum (λemax): PE, λemax ~575 nm; PC, λemax ~635–645 nm; and APC, λemax ~656 nm. The Chl-a pigment also is autofluorescent and has λemax ~675–685 nm, which sometimes may overlap with PC and APC (λemax ~630–685) (Fig. 5).

Percent representation of relative autofluorescence intensity of PBPs and Chl-a emission spectra (after 488 nm laser excitation) of five cyanobacterial isolates exposed to different concentration of arsenite (n = 3)
Under NOEC, fluorescence intensity of PE and Chl-a decreased whereas PC and APC increased compared to control in case of all the cyanobacterial isolates (Fig. 5). A clear decrease in fluorescence intensity was detected for each isolate when exposed to SEC50. For WEK and WEC isolates, very weak fluorescent intensity of PE was detected compared to other three isolates. Maximum PE intensity was recorded in NOK isolate followed by NOC and LBK isolates. More of less stable fluorescence intensity of APC was found in every set of experiments followed by PC in every tested cyanobacterial isolate (Fig. 5).
Nitrogenase activity and EPS production
In the five cyanobacterial isolates, nitrogenase activity in terms of ARA activity was measured. ARA activity in four heterocystous isolates increased about twofold at NOEC (100 μM) compared to control (0.0 μM). Maximum activity was noted in WEK isolate and lowest activity was found in LBK isolate (Table 4). At SEC50, almost 50 % decrease in enzyme activity was noted in all the five isolates (Table 4).
Except in case of LBK isolate, EPS was detected for the remaining isolates under control condition which increased (40–60 %) in dose-dependent manner when exposed to NOEC and SEC50 (Table 4).
Arsenic absorption by cyanobacterial isolates
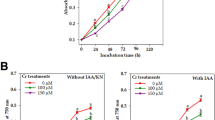
Intracellular accumulation of total As in the studied cyanobacterial isolates under NOEC (100 μM) and SEC50 (200–400 μM) were analysed after 7 days of exposure to sodium arsenite. Absorption experiment was not performed at EC50 to avoid selective low absorption value below 50 % or less viable biomass. After 7 days incubation, all isolates showed ability to absorb As based on BCF values (Fig. 6). Intracellular concentrations of total inorganic As increased with the increase of As(III) dose (Table 5) but BCF value decreased in all isolates. In NOC isolate, highest BCF (0.150) and As absorption (1.96 mg kg−1 at NOEC) was recorded and lowest was found in LBK isolate while intermediate values were found in NOK, WEK and WEC isolates for both treatments (Fig. 6).

Inorganic arsenic absorption ability expressed as bioconcentration factor (BCF) of five cyanobacterial isolates exposed at NOEC and SEC50 in vitro evaluated for respective biomass (±SD; n = 3)
After 7 days of incubation, amount of total As in cyanobacterial growth medium also decreased compared to initial concentrations (NOEC and SEC50 of each isolate). At NOEC, maximum total As was present in the medium which was used to grow WEC isolate (~85 %) followed by growth media representing WEK, LBK, NOK and least in case of medium used to grow NOC isolate (72.30 %) (Table 5). At SEC50 (200–400 μM), higher residual As was detected compared to NOEC (100 μM) which ranged from 0.03 to 4.1 % (Table 5).
Arsenic transformation in cyanobacterial growth medium and biomass
Arsenic transformation in cyanobacterial growth medium and within cyanobacterial biomass was performed from the same set of samples used for total As determination under NOEC and SEC50. A trend for isolate-specific As transformation was observed when treated with sodium arsenite.
In growth medium, under both the concentration (NOEC and SEC50), As(III) and As(V) forms were detected. In growth medium, As(III) fraction was dominant in case of media used to grow NOK and NOC isolates, whereas As(V) was dominant in case of WEK and WEC growth media (Table 6). In the growth medium and biomass of LBK, As(V) was more (~60 %) compared to As(III) (~40 %) at NOEC but almost reverse at SEC50 (Table 6). In all representative As(III)-treated samples except biomass of NOK isolate (at NOEC, 100 μm), both As(III) and As(V) were found in varying amounts (Fig. S1). In biomass representing all the isolates, As(V) was most predominant (Table 6). After 7 days of incubation, intracellular concentration of As(III) increased with the increase in As(III) dose. Highest transformation to As(V) form was recorded in NOC isolate and lowest in LBK isolate and intermediate transformation was found for NOK, WEK and WEC isolates under both treatments (Table 6).
Discussion
Reclamation of contaminated environment and thereby human health safety are two major issues that may be linked to sustainable development (Harnandez et al. 1998; ATSDR-2001) Microorganisms are key players in mediating these processes due to their high ability to thrive in degraded environments such as As-contaminated rice field soils (Costa et al. 2014). It is also believed that native or indigenous population of microorganisms such as cyanobacteria are the best candidates for long-term remediation of contaminated environments (Satoh et al. 2005).
In the present study, soil samples analysed from the two study sites, which are known to have higher As level as reported in previous studies (Nath et al. 2011; Ghosh et al. 2015), were found to be slightly alkaline. As evident, available phosphate and organic carbon concentrations were found to be comparatively lower, and these attributes are known to control the mobility and bioavailability of As(III) (Chatterjee et al. 2010).
Five indigenous diazotrophic cyanobacterial isolates (LBK, NOC, NOK, WEC and WEK) were obtained from the study sites and subsequently characterized to determine potential for arsenite biotransformation and sorption. Polyphasic taxonomic approach was adopted in this study to assign taxonomic rank (genus and species) for cyanobacterial isolates. The LBK isolate showed morphological similarity with Phormidium sp. based on light microscopy. However, using FESEM, deeply constricted trichome with thin and firmly attached sheath was clearly visible in LBK isolate and therefore the taxonomic placement of the same was confirmed within the genus Leptolyngbya (Komarek and Anagnostidis 2005). Additionally, molecular phylogenetic approach reconfirmed the taxonomic affiliation of this isolate with the genus Leptolyngbya. Two other isolates, namely NOC and NOK, showed morphological features which were mainly overlapping and therefore clear taxonomic discrimination was difficult based on available keys (Desikachary 1959; Anagnostidis and Komárek 1990). Morphologically, NOC and NOK isolates closely resembled N. ellipsosporium. The cell length for both isolates showed overlapping with maximum and minimum values as stated earlier in Table 1. Given their quite similar colony and filament morphology and the occurrence of these taxa in the same habitat, it was nearly impossible to distinguish in culture plate using light microscopy. Therefore, the observed morphological characteristics could not be used as diacritical features for assigning these two isolates. In FESEM, surface texture was found to be almost similar in NOC and NOK isolates except the presence of constriction at the cross wall and pattern in line of division in dividing cells in case of NOC isolate. In molecular phylogenetic tree, the 16S rRNA sequences representing NOC and NOK isolates clustered with two different clades with strong bootstrap support; however, both the isolates showed strong taxonomic affiliation with several 16S rRNA sequences representing cultured species belonging to the genus Nostoc. Additionally, the partial 16S rRNA sequence of NOK differed by only 16 bp from the NOC 16S rRNA sequence. These findings also indicate that thorough study involving application of multiple molecular markers may be necessary for species delineation. For the remaining two isolates WEK and WEC, the observed morphological characters including cellular dimensions resembled those of the genus Westiellopsis based on taxonomic keys (Desikachary 1959; Komárek and Anagnostidis 1990). In FESEM, no distinct differences were found between the two isolates except that the cell dimension of branching vary which could be due to phase of development as also reported for the genus Westiellopsis (Komárek and Anagnostidis 1990). The molecular phylogeny representing these two isolates revealed that they belong to the Westiellopsis clade; however, these isolates may represent possibly new species from As-contaminated rice field soil environments as evident from divergent branch length.
Many As-resistant cyanobacterial genera such as Synechocystis, Synechococcus, Phormidium, Oscillatoria, Nostoc, Anabaena and Hapalosiphon have been reported as dominant taxa in As-contaminated aquatic environments globally (Maeda et al. 1998; Shaheen et al. 2007; Banerjee 2008; Yin et al. 2012; Sánchez-Riego et al. 2014; Zutshi et al. 2014). Cyanobacteria isolated from the study sites and belonging to the genus Westiellopsis and Leptolyngbya are the first report in terms of their presence in As-contaminated rice field ecosystems.
Arsenic metabolism has been widely reported in microbes and plants (e.g., Rosen 2002). In the present investigation, As(III) was used for toxicity assessment on cyanobacterial isolates in vitro. Under NOEC (100 μM) and SEC50 (200–400 μM), cyanobacterial isolates were able to accumulate As and grow; however, growth rate slowed with increasing As concentration. In particular, the isolates NOC and NOK (Nostoc) were most tolerant followed by WEC and WEK isolates (Westiellopsis clade) and LBK isolate (Leptolyngbya). Similar observations were also reported for Synechocystis sp. PCC6803 (Yin et al. 2011, 2012). Microcystis sp. PCC7806 (Yin et al. 2011), Microcystis aeruginosa (Wang et al. 2013), Oscillatoria tenuisa, Nostoc sp. PCC7120 (Yin et al. 2011), Nostoc muscorum and Nostoc minutum (Ferrari et al. 2013), Anabaena affinis, Tolypothrix tenuis (Ferrari et al. 2013) and Hapalosiphon fontinalis-339 (Zutshi et al. 2014) isolated from diverse aquatic environments and when exposed to As(V) or As(III). As(III) exerted strongest toxic effect causing cell death in the range of 200–500 μM concentration among the isolates and more so at the intrageneric level. Ferrari et al. (2013) showed that sensitivity to As(III) varied among different species of the genus Nostoc and the same has been observed in our study between NOC and NOK isolates.
In this study, CLSM approach was adopted to detect physiological change in cyanobacterial isolates in terms of in vivo fluorescence properties of photosynthetic pigments as an indicator of cell adaptability and survivality (Billi et al. 2011; Schulze et al. 2011). Cyanobacteria possess highly abundant autofluorescent PBPs and Chl-a, which can be used as intrinsic fluorescent markers for probing the regulation of their synthesis, accumulation and photosynthetic activity under natural and stress conditions (Bordowitz and Montgomery 2010). In the present study, PBPs (10–25 %) and Chl-a (20–30 %) autofluorescence decreased with the increase in As(III) concentration as evident from SEC50 values (Fig. 6). Loss of pigment fluorescence (Geider et al. 1993) has been correlated with decreased enzyme activity and increased membrane permeability which could be a useful indicator of senescence for any species of algae (Franklin et al. 2012). We also found loss of pigment fluorescence in our study. Once this process begins, degradation of the photosynthetic pigments, in particular chlorophyll (Berges et al. 1998), leads to the final stage of autolysis and this was observed in our study based on quantified fluorescence loss at SEC50. However, the persistence of PBPs autofluorescence serves as a survival marker and has been related to genome stability, undamaged plasma membranes and dehydrogenase activity (Billi et al. 2011). Thus, in our study, consistent or even higher APC fluorescence was found in all the tested cyanobacterial isolates thereby indicating adaptability under As(III) exposure for survival and performance of essential cellular functions such as efficient energy transfer from PC to Chl-a via APC.
In four out of five isolates, namely NOC, NOK, WEC and WEK, nitrogenase activity increased by almost twofold when exposed to NOEC (100 μM) but reduced to half under SEC50 (200–400 μM) compared to control conditions. Nitrogen fixation and its incorporation in cyanobacterial cell (ammonium assimilation) depend on cellular energy and represent a complex interdependent metabolic cycle. This energy and reducing power normally come from cell’s metabolic energy through TCA cycle and electron transport chain (Wallsgrove et al. 1983). In heterocystous cyanobacteria, photosynthetically fixed carbon which is essential for nitrogen fixation is supplied to the heterocyst by the vegetative cells (Stewart 1980) and plays a key role in the linking of nitrogen and carbon metabolism (Chen et al. 1988). Thus, in our study under NOEC, almost twofold increase in ARA activity might be coupled with the increased APC florescence in terms of photosynthetic energy transfer from antenna molecule to reaction centre PS II. These phenomena might enhance photosynthetic activity and energy production processes for cellular metabolism. The reduced nitrogenase activity under increased concentration (SEC50) of As(III) in LBK isolate may be related to reduction in photosynthetic activity due to lack of reducing agent pool (Leganés and Fernández-Valiente 1992) under As stress as indicated from our CLSM results. Thus ARA activity might be hampered in two ways, either by deficiency in energy supply by inhibition of photosynthesis or by inhibition of energy metabolism and reduction of supply of reducing molecules such as NADH or NADPH (Debnath et al. 2012). On the other hand, at SEC50, when nitrogen fixation was seriously inhibited, cyanobacterial cells may suffer from nitrogen deficiency. Nitrogen stress affects energy transfer from PC and Chl-a, and therefore, their spectral properties under both in vivo and in vitro conditions may change (Peter et al. 2010) and these may lead to low levels of photosynthesis during nitrogen limitation (Duke et al. 1989; Sauer et al. 2001). Therefore in our study, decrease in ARA activity corresponded to decrease in pigment fluorescence intensity.
In NOC, NOK, WEC and WEK isolates, EPS production increased under NOEC and SEC50 treatments. In the natural environment, bacteria usually grow within structured microbial communities embedded in EPS composed of sugars, proteins and DNA (Hall-Stoodley et al. 2004) which help them to thrive in high As environment (Muller et al. 2007) as also reported for Thiomonas arsenitoxydans when exposed to As(III) (Marchal et al. 2011). This is an adaptive response to changing ecological niche. The hyper-production of exopolysaccharide in cyanobacteria in response to metals also reflects one of the strategies for survival under stress, as these polymers have potential to interact strongly with cations as metal chelators (Kiran and Kaushik 2008; Huertas et al. 2014). This property has also been demonstrated in the green alga Chlorella sp. (Kaplan et al. 1987). The stimulation of exopolymer biosynthesis by Cu, Ni or Hg cations was first reported in the cyanobacterium Nostoc spongiforme (Singh et al. 1999) contrary to previous suggestions about the role C/N ratio in regulation of EPS (Bergmann 1986). In our study, increased EPS production may possibly indicate the role of As-mediated regulation which warrants further investigation.
Based on the As bioabsorption and transformation experiments, it was found that the As-tolerant cyanobacterial isolates not only grow at high As(III) concentration, but also have the ability to absorb substantial amounts of As. It has been reported that prokaryotes adapt to high concentration of As by activation or deactivation of As transporter genes or efflux pump genes (e.g., Yin et al. 2011; Sounders and Rocap 2015). In this study, absorption of As increased with increase in As(III) concentration in the growth media but not based on BCF values. Previous studies have documented similar finding in Synechococcus leopoliensis, Chlorella vulgaris, Synechocystis sp. PCC6803, Microcystis sp. PCC7806, Nostoc sp. PCC7120, and M. aeruginosa (Budd and Craig 1981; Maeda et al. 1992a, b; Yin et al. 2011; Wang et al. 2013). Three out of five isolates, namely NOC, NOK and WEK, exhibited higher capability to bioabsorb As (~0.958–2.870 mg kg−1 and BCF ~0.074–0.220 at NOEC) based on analysis of dry biomass compared to available reports involving freshwater algae (Levy et al. 2005; Yin et al. 2011; Wang et al. 2013). Therefore these isolates can be potentially used for bioremediation of As-contaminated rice fields located in BDP.
During exposure studies, the major As species detected in cyanobacterial isolates was the less toxic inorganic As(V). The transformation ability was found to be isolate-specific and highest in the experimental biomass of NOK isolate. Similar findings have been also reported for different species of algae (Yin et al. 2011; Wang et al. 2013). In our study, oxidation of As(III) to As(V) was detected in the growth medium as well as in cyanobacterial biomass when exposed to As(III). The process of As(III) to As(V) conversion in the culture was directly mediated by the cyanobacterial cells. The toxicity of As is dependent on speciation, with inorganic arsenicals thought to be more toxic than organic forms (Tamaki and Frankenberger 1992). It must therefore be assumed that As-resistant organisms either compartmentalize and/or transform As to other less phytotoxic arsenic species in order to withstand high cellular arsenic burden (Meharg 1994; Qin et al. 2009). Our results indicate that such pathways may exist in the five tested cyanobacterial isolates. It has been reported that As(III) within the cyanobacterial cell may be complexed with phytochelatins (PCs) (Sneller et al. 1999). When these PCs are extracted in media, PC-complexes may dissociate to form free PCs and As(V) [reduction of the PCs and oxidation of the As(III) to As(V)]. Thus, when As(V) was detected in biomass extracts as part of our study, it may have originated from PC complexed As(III). Yin et al. (2011) investigated As biotransformation in three cyanobacterial species (Microcystis sp. PCC7806, Nostoc sp. PCC7120 and Synechocystis sp. PCC6803). They encountered inorganic As(V) and As(III) as the predominant species, with As(V) making up more than 80 % of total As.
Bacterial adaptive response to As has been demonstrated in two ways (Cleiss-Arnold et al. 2010). First, bacterial cells express various genes involved in defense mechanisms and next several metabolic activities including for As(III) oxidation. Oxidation of As(III) to As (V) is an energy-producing reaction that is also believed to be a detoxification process since As(V) is less toxic than As(III) (Páez-Espino et al. 2009) and play a crucial role in microbial physiology (Stolz 2011). Generally, bio-oxidation is the first and primary transformation mechanism for As(III) biotransformation, while bioreduction together with subsequent methylation may play an important role in As(V) biotransformation (Qin et al. 2009; Bhattacharjee and Rosen 2007) and biological production of volatile As compounds is considered to be an important part in global As biogeochemical cycling (Michalke and Hensel 2004). Rapid changes of As speciation that occurred in the growth media containing inorganic As species indicated that the cyanobacterial isolates were able to detoxify inorganic As. Decrease of As(III) and increase of As(V) occurred synchronously in the growth medium after 7 days As(III) exposure. Moreover, total As in the medium decreased to 72–84 % within 7 days depending on the cyanobacterial isolate tested. The decrease in total As from culture medium showed the biological potential of these isolates to be able to remove As from contaminated flooded rice fields located in BDP. In particular, the dominant As species in cyanobacterial cells was As(V) regardless of exposure concentration, thereby illustrating the detoxification potential of these cyanobacterial isolates.
Previous studies have shown that the abundance of As(III)-oxidizing microbes could enhance As(III) oxidation to As(V) in the rice rhizosphere, resulting in decreased As mobility and bioavailability since As(V) can be sequestered on Fe/Mn hydroxide/oxyhydroxide in rhizosphere soil as well as in rice roots (Liu et al. 2006; Seyfferth et al. 2010). In several heterotrophic and chemoautotrophic microorganisms, As(III) oxidation is catalysed by As(III) oxidases, which are encoded by aioA and aioB genes of the aio enzyme (Hamamura et al. 2009; Slyemi and Bonnefoy 2012). In autotrophic cyanobacteria such as Synechocystis sp. PCC 6830, arsenic oxidation, reduction and methylation are mediated by the arsBHC operon (Sanchez-Reigo et al. 2014 and references therein for details). In the cyanobacteria Synechocystis sp. PCC 6803 and Nostoc sp. PCC7120, ArsM pathway exist with three ArsM homologs for As(III) resistance and to catalyse methylation of As(III) and the ArsM from Nostoc sp. PCC7120 exhibits highest methylation capacity (Yin et al. 2011 and references therein for detail). In Prochlorococcus populations of global marine ecosystem, the importance of the ArsM pathway (Qin et al. 2006; Paes-Espino et al. 2009) as well as As(III)-specific efflux detoxification pathway has been also reported (Saunders and Rocap 2015). In another study, Zhang et al. (2015) undertook a study to show microbe-mediated As biotransformation in paddy soils across South China. They showed wide distribution of arsenite oxidase genes (aioA), respiratory arsenate reductase genes (arrA), arsenate reductase genes (arsC) and arsenite adenosylmethionine methyltransferase genes (arsM) in paddy soils. The abundance of aioA genes was almost 10 times higher than arsM gene abundance, but the diversity and the OTU richness of arsM genes were much higher than aioA, suggesting highly diverse microbial communities are involved in As methylation in paddy fields. It would be therefore important to investigate genetic mechanisms of As bioaccumulation and biotransformation for our cyanobacterial isolates targeted from the As-contaminated rice fields of BDP as part of future studies.
Overall, as part of this study, five most abundant cyanobacterial isolates with varying metabolic potential have been screened for the first time from As-contaminated rice fields of Bengal Delta Plains. Our findings indicate that all the indigenous isolates showed ability to absorb and transform As(III), an inorganic form of As, and could be potentially used for bioremediation of As-contaminated rice field environments through extensive trials. Additionally, these fast-growing cyanobacterial isolates with moderate nitrogen fixation ability could additionally used for as a soil biofertilizer. To conclude, application of cyanobacterial biomass representing these isolates, which can significantly bioabsorb and biotransform As in rice fields, may ultimately result in less bio-available As for uptake by rice plants via plant transport system, thereby rendering food grains safe for human consumption in BDP as well as in South Asia.
References
Alam MK, Hassan AKMS, Khan MR, and Whitney JW (1990) Geological map of Bangladesh, Geol. Surv. of Bangladesh, Dhaka.
Anagnostidis K, Komárek J (1990) Modern approach to the classification system of cyanophytes. Teil 5-Stigonematales. Arch Hydrobiol Algol Stud 59:1–73
ATSDR (2001) CERCLA Priority list of hazardous substances. Available at http://www.atsdr.cdc.govlelist.html (last Access 4 October 2015)
Bahar MM, Meghraj M, Naidu R (2013) Toxicity, transformation and accumulation of inorganic arsenic species in a microalga Scenedesmus sp. isolated from soil. J Appl Phycol 25:913–917
Banerjee M (2008) Arsenic. A threatening environmental issue and role of cyanobacteria in toxicity mitigation. J Ecophysiol Occup Health 8:153–159
Bary RH, Kürtz LT (1945) Determination of total, organic and available forms of phosphorus in soils. Soil Sci 59:39–45
Berges JA, Falkowski PG (1998) Physiological stress and cell death in marine phytoplankton: induction of proteases in response to nitrogen or light limitation. Limnol Oceanogr 43:129–135
Bergman B (1986) Glyoxylate induced changes in the carbon and nitrogen metabolism of the cyanobacterium Anabaena cylindrica. Plant Physiol 80:698–701
BGS & DPHE (2001) Arsenic contamination of groundwater in Bangladesh. Technical report WC/00/19.4. In: Kinniburgh DG, Smedley PL (eds) British Geological survey, Keyworth
Bhattacharjee H, Rosen BP (2007) Arsenic metabolism in prokaryotic and eukaryotic microbes. In: Nies DH, Silver S (eds) Molecular microbiology of heavy metals. Springer, Berlin, pp 371–406
Bhattacharya P, Chatterjee D, Jacks G (1997) Occurrence of arsenic-contaminated groundwater in alluvial aquifers from delta plains, Eastern India: options for safe drinking water supply. Water Resour Dev 13:79–92
Bhattacharya P, Jacks G, Jana J, Sracek A, Gustafsson JP, Chatterjee D (2001) Geochemistry of the Holocene alluvial sediments of Bengal Delta Plain from West Bengal, India; implications on arsenic contamination in ground-water. In: Jacks G, Bhattacharya P, Khan AA (eds) Ground water arsenic contamination in the Bengal Delta Plain of Bangladesh: KTH special publication. TRITA-AMI Report, Stockholm, pp 21–40
Bhattacharya P, Samal AC, Majumdar J, Santra SC (2009) Transfer of arsenic from ground water and paddy soil to rice plant (Oryza sativa L.): a micro level study in West Bengal, India. World J Agric Sci 5:425–431
Billi D, Viaggiu E, Cockell CS, Rabbow E, Horneck G, Onofri S (2011) Damage escape and repair in dried Chroococcidiopsis spp. from hot and cold deserts exposed to simulated space and Martian conditions. Astrobiology 11:65–73
Boivin P, Favre F, Hammecker C, Maeght JL, Delarivière J, Poussin JC, Wopereis MCS (2002) Processes driving soil solution chemistry in a flooded rice-cropped vertisol: analysis of longtime monitoring data. Geoderma 110:87–107
Bordowitz JR, Montgomery BL (2010) Exploiting the autofluorescent properties of photosynthetic pigments for analysis of pigmentation and morphology in live Fremyella diplosiphon cells. Sensors 10:6969–6979
Boström KH, Simu Hagström Å, Riemann L (2004) Optimization of DNA extraction for quantitative marine bacterioplankton community analysis. Limnol Oceanogr Methods 2:365–373
Budd K, Craig SR (1981) Resistance to arsenate toxicity in the blue green alga Synechococcus leopoliensis. Can J Bot 59:1518–1521
Chatterjee D, Halder D, Majumder S, Biswas A, Nath B, Bhattacharya P, Bhowmick S, Mukherjee-Goswami A, Saha D, Hazra R, Maity PB, Chatterjee D, Mukherjee A, Bundschuh J (2010) Assessment of arsenic exposure from groundwater and rice in Bengal Delta region, West Bengal India. Water Res 44:5803–5812
Chen RD, Le MP, Vidal J, Jacquot JP, Gadal R (1988) Purification and comparative properties of the cytosolic isocitrate dehydrogenase (NADP) from pea (Pisum sativum) roots and green leaves. Eur J Biochem 175:565–572
Chowdhury UK, Biswas BK, Chowdhury TR, Samanta G, Mandal BK, Basu GC (2000) Ground water arsenic contamination in Bangladesh and West Bengal, India. Environ Health Perspect 108:393–397
Cleiss-Arnold J, Koechler S, Proux C, Fardeau M-L, Dillies M-A, Coppee J-Y, Arsene-Ploetze F, Bertin PN (2010) Temporal transcriptomic response during arsenic stress in Herminiimonas arsenicoxydans. BMC Genomics 11:709
Costa PS, Scholte LLS, Reis MP, Chaves AV, Oliveira PL, Itabayana LB, Suhadolnik MSL, Barbosa FAR, Chartone-Souza E, Nascimento AMA (2014) Bacteria and genes involved in arsenic speciation in sediment impacted by long-term gold mining. PLoS One 9:e95655
Datta BK, Mishra A, Singh A, Sar TK, Sarkar S, Bhattacharya A, Chakraborty AK, Mandal TK (2010) Chronic arsenicosis in cattle with special reference to its metabolism in arsenic endemic village of Nadia district West Bengal India. Sci Total Environ 409:284–288
Debnath M, Mandal NC, Ray S (2012) Effect of fungicides and insecticides on growth and enzyme activity of four cyanobacteria. Indian J Microbiol 52:275–280
Desikachary TV (1959) Cyanophyta. (Monographs on algae). Indian Council of Agricultural Research, New Delhi, p 686
Dubois M, Gilles KA, Hamilton JK, Rebers PA, Smith F (1956) Colorimetric method for determination of sugars and related substances. Anal Chem 28:350–356
Duke CS, Cezeaux A, Allen MM (1989) Changes in polypeptide composition of Synechocystis sp. strain 6308 phycobilisomes induced by nitrogen starvation. J Bacteriol 160:1960–1966
FAO (1985) Water quality guide lines for maximum crop production. Food and Agricultural Organization/UN. Available online at:www.fao.org/docrep/ T0551E.2006/9/13
Ferrari SG, Silvaa PG, Gonzálezb DM, Navonic JA, Silvaa HJ (2013) Arsenic tolerance of cyanobacterial strains with potential use in biotechnology. Rev Argent Microbiol 45:174–179
Franklin DJ, Airs RL, Fernandez M, Bell TG, Bongaerts RJ, Berges JA, Malin G (2012) Identification of senescence and death in Emiliania huxleyi and Thalassiosira pseudonana: cell staining, chlorophyll alterations, and dimethylsulfoniopropionate (DMSP) metabolism. Limnol Oceanogr 57:305–317
Gao S, Tanji KK, Scardaci SC, Chow AT (2002) Comparison of redox indicators in a paddy soil during rice-growing season. Soil Sci Soc Am J 66:805–817
Geider RJ, Laroche J, Greene RM, Olaizola M (1993) Response of the photosynthetic apparatus of Phaeodactylum tricornutum (Bacillariophyceae) to nitrate, phosphate, or iron starvation. J Phycol 29:755–766
Ghosh D, Bhadury P, Routh J (2014) Diversity of arsenite oxidizing bacterial communities in arsenic rich deltaic aquifers in West Bengal, India. Front Microbiol 5:1–14
Ghosh D, Routh J, Bhadury P (2015) Characterization and microbial utilization of dissolved lipid organic fraction in arsenic impacted aquifers (India). J Hydrol 527:221–231
Guillard RL (1973) Division rates. In: Stein JR (ed) Handbook of phycological methods.culture methods and growth measurements. Cambridge University Press, USA, pp 290–311
Hall TA (1999) BioEdit: a user-friendly biological sequence alignment editor and analysis programme for Windows95/98/NT. Nucleic Acids Symp 41:95–98
Hall-Stoodley L, Costerton JW, Stoodley P (2004) Bacterial biofilms: from the natural environment to infectious diseases. Nat Rev Microbiol 2:95–108
Hamamura N, Macur RE, Korf S, Ackerman G, Taylor WP, Kozubal M, Reysenbach AL, Inskeep WP (2009) Linking microbial oxidation of arsenic with detection and phylogenetic analysis of arsenite oxidase genes in diverse geothermal environments. Environ Microbiol 11:421–431
Harnandez A, Mellado RP, Martinez JL (1998) Metal accumulation and vanadium induced multidrug resistance by environmental isolates of Escherichia herdmanni and Enterobacter cloaeae. Appl Environ Microbiol 64:4317–4320
Hasegawa H, Rahman MA, Kitahara K, Itaya Y, Maki T, Ueda K (2010) Seasonal changes of arsenic speciation in lake waters in relation to eutrophication. Sci Total Environ 408:1684–1690
Huertas MJ, López-Maury L, Giner-Lamia J, Sánchez-Riego AM, Florencio FJ (2014) Metals in cyanobacteria: analysis of the copper, nickel, cobalt and arsenic homeostasis mechanisms. Life 4:865–886
Jackson CR, Dugas SL (2003) Phylogenetic analysis of bacterial and archaeal arsC gene sequences suggests an ancient, common origin for arsenate reductase. BMC Evol Biol 3:18–28
Kaplan D, Christiaen D, Arad SM (1987) Chelating properties of extracellular polysaccharides from Chlorella spp. Appl Environ Microbiol 53:2953–2956
Kiran B, Kaushik A (2008) Chromium binding capacity of Lyngbya putealis exopolysaccarides. Biochem Eng J 38:47–54
Komárek J, Anagnostidis K (2005) Cyanoprokaryota. Part 2. Oscillatoriales. Elsevier Spektrum, Munich, p 759
Komárek J, Kastrovosý J, Marešl J, Johansen JR (2014) Taxonomic classification of cyanoprokaryotes (cyanobacterial genera) 2014, using a polyphasic approach. Preslia 86:295–335
Lakshmipathiraj P, Narasimhan BRV, Prabhakar S, Raju B (2006) Adsorption of arsenate on synthetic goethite from aqueous solutions. J Hazard Mater 136:281–287
Leganés F, Fernéndez-Valiente E (1992) Effects of phenoxyacetic herbicides on growth, photosynthesis and nitrogenase activity in the cyanobacteria from rice fields. Arch Environ Contam Toxicol 22:130–134
Levy JL, Stauber JL, Adams MS, Maher WA, Kirby JK, Jolley DF (2005) Toxicity, biotransformation, and mode of action of arsenic in two freshwater microalgae (Chlorella sp. and Monoraphidium arcuatum). Environ Toxicol Chem 24:2630–2639
Liu WJ, Zhu YG, Hu Y, Williams PN, Gault GA, Meharg AA, Charnock JM, Smith FA (2006) Arsenic sequestration in iron plaque, its accumulation and speciation in mature rice plants (Oryza sativa L.). Environ Sci Technol 40:5730–5736
López-Maury L, Florencio FJ, Reyes JC (2003) Arsenic sensing and resistance system in the cyanobacterium Synechocystis sp. strain PCC 6803. J Bacteriol 185:5363–5371
Mackinney G (1941) Absorption of light by chlorophyll solutions. J Biol Chem 40:315–322
Maeda S, Ohki A (1998) Bioaccumulation and biotransformation of arsenic, antimony and bismuth compounds by freshwater algae. In: Wong YS, Tam NFY (eds) Wastewater treatment with algae, 2nd edn. Springer, Berlin, pp 73–91
Maeda S, Kusadome K, Arima H, Ohki A, Naka K (1992a) Biomethylation of arsenic and its excretion by the alga Chlorella vulgaris. Appl Organomet Chem 6:407–413
Maeda S, Kusadome K, Arima H, Ohki A, Naka K (1992b) Uptake and excretion of total inorganic arsenic by the freshwater alga Chlorella vulgaris. Appl Organomet Chem 6:399–405
Marchal M, Briandet R, Halter D, Koechler S, Dubow MS, Lett M-C, Bertin PN (2011) Subinhibitory arsenite concentrations lead to population dispersal in Thiomonas sp. PLoS One 6:e23181
Meharg AA (1994) Integrated tolerance mechanisms-constitutive and adaptive plant- responses to elevated metal concentrations in the environment. Plant Cell Environ 17:989–993
Meharg AA, Williams PN, Adomako E, Lawgali YY, Deacon C, Villada A, Cambell RCJ, Sun G, Zhu Y-G, Feldmann J, Raab A, Zhao F-J, Islam R, Hossain S, Yanai J (2009) Geographical variation in total and inorganic arsenic content of polished (white) rice. Environ Sci Technol 43:1612–1617
Michalke K, Hansel R (2004) Biovolatilisation of metal(loid)s by microorganisms. In: Hiran AV, Emons H (eds) Organic metal and metalloid species in the environment. Springer, Berlin, pp 137–50
Mondal D, Polya DA (2008a) Rice is a major exposure route for arsenic in Chakdah block, Nadia district, West Bengal, India: a probabilistic risk assessment. Appl Geochem 23:2987–2998
Mondal D, Polya DA (2008b) Rice is a major exposure route for arsenic in Chakdaha block, Nadia district, West Bengal, India: a probabilistic risk assessment. Appl Geochem 23:2986–2999
Morelli E, Mascherpa MC, Scarano G (2005) Biosynthesis of phytochelatins and arsenic accumulation in the marine microalga Phaeodactylum tricornutum in response to arsenate exposure. Biometals 18:587–593
Muhling M, Woolven-Allen J, Murrell JC, Joint I (2008) Improved group-specific PCR primers for denaturing gradient gel electrophoresis analysis of the genetic diversity of complex microbial communities. ISME J 2:379–392
Muller D, Médigue C, Koechler S, Barbe V, Barakat M, Talla E (2007) A tale of two oxidation states: bacterial colonization of arsenic-rich environments. PLoS Genet 3:e53
Nath B, Maity JP, Jean J-S, Birch G, Kar S, Yang H-J, Lee M-K, Hazra R, Chatterjee D (2011) Geochemical characterization of the arsenic-affected alluvial aquifers of the Bengal Delta (West Bengal and Bangladesh) and Chianan Plains (SW Taiwan): implications for human health. Appl Geochem 26: 705e713.
Newbigging AM, Paliwoda RE, Le XC (2015) Rice: reducing arsenic content by controlling water irrigation. J Environ Sci 30:129–131
Nickson RT, McArthur JM, Ravenscroft P, Burgess WG, Ahmed KM (2000) Mechanism of arsenic poisoning of ground water in Bangladesh and West Bengal. Appl Geochem 15:403–413
Nordstrom DK (2002) Worldwide occurrences of arsenic in ground water. Science 296:2143–2145
Ohno K, Yanase T, Matsuo Y, Kimura T, Rahman MH, Magara Y, Matsui Y (2007) Arsenic intake via water and food by a population living in an arsenic-affected area of Bangladesh. Sci Total Environ 381:68–76
Oremland RS, Stolz JF (2003) The ecology of arsenic. Science 300:939–944
Páez-Espino D, Tamames J, de Lorenzo V, Cánovas D (2009) Microbial responses to environmental arsenic. Biometals 22:117–130
Pawlik-Skowrónska B, Pirszel J, Kalinowska R, Skowrónski T (2004) Arsenic availability, toxicity and direct role of GSH and phytochelatins in As detoxification in the green alga Stichococcus bacillaris. Aquat Toxicol 70:201–212
Peter P, Sarma AP, Hasan MDA, Murthy SDS (2010) Studies on the impact of nitrogen starvation on the photosynthetic pigments through spectral properties of the cyanobacterium, Spirulina platensis: identification of target phycobiliprotein under nitrogen chlorosis. Bot Res Int 3:30–34
Qin J, Rosen BP, Zhang Y, Wang G, Franke S, Rensing C (2006) Arsenic detoxification and evolution of trimethylarsine gas by a microbial arsenite S-adenosylmethionine methyltransferase. Proc Natl Acad Sci U S A 103:2075–2080
Qin J, Lehr CR, Yuan CG, Le XC, McDermott TR, Rosen BP (2009) Biotransformation of arsenic by a Yellowstone thermoacidophilic eukaryotic alga. Proc Natl Acad Sci U S A 106:5213–5217
Rahaman S, Sinha AC, Pati R, Mukhopadhyay D (2013) Arsenic contamination: a potential hazard to the affected areas of West Bengal. Environ Geochem Health 35:119–132
Rahman MA, Hassler C (2014) Is arsenic biotransformation a detoxification mechanism for microorganisms? Aquat Toxicol 146:212–219
Rahman MM, Choudhury UK, Mukherjee SC, Mondal BK, Paul K, Lodh D, Biswas BK, Chanda CR, Basu GK, Saha KC, Roy S, Das R, Palit SK, Quamruzzaman Q, Chakraborti D (2001) Chronic arsenic toxicity in Bangladesh and West Bengal India. J Toxicol Clin Toxicol 39:683–700
Rossmann TG (2003) Mechanism of arsenic carcinogenesis: an integrated approach. Mutat Res 533:37–65
Rosen BP (2002) Biochemistry of arsenic detoxification. FEBS Lett 529:86–92
Sánchez-Riego AM, López-Maury L, Florencio FJ (2014) Genomic responses to arsenic in the cyanobacterium Synechocystis sp. PCC 6803. PLoS One 9:e96826
Satoh A, Vudikaria LQ, Kurano N, Miyachi S (2005) Evaluation of the sensitivity of marine microalgal strains to the heavy metals, Cu, As, Sb, Pb and Cd. Environ Int 31:713–722
Saunders JK, Rocap G (2015) Arsenic detoxification strategies vary with environmental phosphate concentrations in global Prochlorococcus populations. ISME J. doi:10.1038/ismej.2015.85
Sauer J, Schreiber U, Schmid R, Vo¨ lker U, & Forchhammer K (2001) Nitrogen starvationinduced chlorosis in Synechococcus PCC 7942. Low-level photosynthesis as a mechanism of long-term survival. Plant Physiol 126, 233–243
Schulze K, López DA, Tillich UM, Frohme M (2011) A simple viability analysis for unicellular cyanobacteria using a new autofluorescence assay, automated microscopy, and ImageJ. BMC Biotechnol 11:118–125
Seyfferth AL, Webb SM, Andrews JC, Fendorf S (2010) Arsenic localization, speciation, and co-occurrence with iron on rice (Oryza sativa L.) roots having variable Fe coatings. Environ Sci Technol 44:8108–8113
Shaheen R, Mahmud R, Sen J (2007) A study on arsenic decontaminating cyanobacteria of an arsenic affected soil. J Soil Nature 1:23–29
Singh N, Asthana RK, Kayastha AM, Pandey S, Chaudhary AK, Singh SP (1999) Thiol and exopolysaccharide production in a cyanobacterium under heavy metal stress. Process Biochem 35:63–68
Sinha RP, Sinha JP, Gröniger A and Häder D-P (2002) Polychromatic action spectrum for the induction of a mycosporine-like amino acid in a rice-field cyanobacterium, Anabaena sp., J. Photochem. Photobiol., B 66:47–53
Slyemi D, Bonnefoy V (2012) How prokaryotes deal with arsenic. Environ Microbiol Rep 4:571–586
Smedley PL, Kinniburgh DG (2002) The review of source, behaviour and distribution of arsenic in natural waters. Appl Geochem 17:517–568
Sneller FEC, Van Heerwaarden LM, Kraaijeveld-Smit FJL, Ten Bookum WM, Koevoets PLM, Schat H, Verkleij JAC (1999) Toxicity of arsenate in Silene vulgaris, accumulation and degradation of arsenate-induced phytochelatins. New Phytol 144:223–232
Stanier RY, Kunisawa R, Mandel M, Cohen-Bazire G (1971) Purification and properties of unicellular blue-green algae (Order Chroococcales). Bacteriol Rev 35:171–205
Stewart WDP (1980). Some aspects of structure and function in N2-hing cyanobacteria. Annual Review of Microbiology 34:497–536
Stolz JF (2011) Microbial metal and mettaloid metabolism: advances and applications, 1st edn. ASM Press, Washington DC
Subbaiah BV, Asija GC (1956) A rapid procedure for determination of available nitrogen in soils. Curr Sci 25:259–260
Tamaki S, Frankenberger WT Jr (1992) Environmental biochemistry of arsenic. Rev Environ Contam Toxicol 124:79–110
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013) MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol Biol Evol 30:2725–2729
Thiel T (1988) Phosphate transport and arsenate resistance in the cyanobacterium Anabaena variabilis. J Bacteriol 170:1143–1147
Turner GL, Gibson AH (1980) Measurement of nitrogen fixation by indirect means. In: Bergerson FJ (ed) Methods for evaluating biological nitrogen fixation. Wiley, UK, pp 111–138
Vaughan DJ (2006) Arsenic. Elements 2:71–75
Venkataraman GS (1993) Blue-green algae (cyanobacteria). In: Tata Wadhwani AM, Mehdi MS (eds) Biological nitrogen fixation. Indian Council of Agric Res, New Delhi, pp 45–76
Walkley A, Black IA (1934) An examination of the Degrjareff method for determining soil organic matter and a proposed modification of the chromic acid titration method. Soil Sci 37:29–36
Wallsgrove RM, Keys AJ, Lea PJ, Miflin BJ (1983) Photosynthesis, photorespiration and nitrogen metabolism. Plant Cell Environ 6:301–309
Wang NX, Li Y, Deng XH, Miao AJ, Ji R, Yang LY (2013) Toxicity and bioaccumulation kinetics of arsenate in two freshwater green algae under different phosphate regimes. Water Res 47:2497–2506
WHO (2011) Guidelines for drinking-water quality. Fourth Edition. World Health Organization, Geneva, Switzerland Available: http:// http://www.who.int/water_sanitation_health/dwq/guidelines/en
Williams PN, Villada A, Deacon C, Raab A, Figuerola J, Green AJ, Feldmann J, Meharg AA (2007) Greatly enhanced arsenic shoot assimilation in rice leads to elevated rice grain levels compared to wheat and barley. Environ Sci Technol 41:6854–6859
Yin XX, Chen J, Qin J, Sun GX, Rosen BP, Zhu YG (2011) Biotransformation and volatilization of arsenic by three photosynthetic cyanobacteria. Plant Physiol 156:1631–1638
Yin X, Wang LH, Bai R, Huang H, Sun G (2012) Accumulation and transformation of arsenic in the blue-green alga Synechocysis sp. PCC6803. Water Air Soil Pollut 223:1183–1190
Zhang SY, Zhao FJ, Guo XS, Su JQ, Yang XR, Li H, Zhu YG (2015) Diversity and abundance of arsenic biotransformation genes in paddy soils from Southern China. Environ Sci Technol 49:4138–4146
Zhao FJ, Zhu YG, Meharg AA (2013) Methylated arsenic species in rice: geographical variation, origin, and uptake mechanisms. Environ Sci Technol 47:3957–3966
Zheng MZ, Li G, Sun GX, Shim H, Cai C (2013) Differential toxicity and accumulation of inorganic and methylated arsenic in rice. Plant Soil 365:227–238
Zhu YG, Williams PN, Meharg AA (2008) Exposure to inorganic arsenic from rice: a global health issue? Environ Pollut 154:169–171
Zutshi S, Bano F, Ningthoujam M, Habib K, Fatma T (2014) Metabolic adaptations to arsenic-induced oxidative stress in Hapalosiphon fontinalis-339. IJIRSET 3:9386–9394
Acknowledgments
Manojit Debnath from PostGraduate Department of Botany, Hooghly Mohsin College, Chinsurah, thankfully acknowledges IISER Kolkata for the provision of IISERK Postdoctoral Fellowship to carry out this research. The FESEM and CLSM as part of Central Imaging Facility of IISERK are acknowledged. This work is supported by FIRE grant of IISERK awarded to Punyasloke Bhadury.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Table S1
Blastn result of 16S rRNA sequences representing the five cyanobacterial isolates compared with published sequences of cultured cyanobacteria available in GenBank/EMBL/DDBJ databases (DOCX 19.9 kb)
Fig. S1
Arsenic speciation of five cyanobacterial isolates exposed at NOEC in vitro evaluated in biomass and respective growth medium (n=3) (PDF 106 kb)
Fig. S2
Arsenic speciation of five cyanobacterial isolates exposed at SEC50 in vitro evaluated in biomass and respective growth medium (n=3) (PDF 106 kb)
Rights and permissions
About this article

Cite this article
Debnath, M., Bhadury, P. Adaptive responses and arsenic transformation potential of diazotrophic Cyanobacteria isolated from rice fields of arsenic affected Bengal Delta Plain. J Appl Phycol 28, 2777–2792 (2016). https://doi.org/10.1007/s10811-016-0820-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10811-016-0820-9