
Abstract
The diatom Phaeodactylum tricornutum produces large quantities of lipids, especially triacylglycerols (TAGs) under nitrogen or phosphorus limitation. In this study, production of lipids and TAGs during this process was compared under conditions with different inputs of inorganic carbon. With an abundant supply of inorganic carbon, considerable accumulation of biomass, lipids, and TAGs was identified after a nitrogen/phosphorus-limiting “induction incubation.” TAGs were still synthesized and accumulated even under inorganic carbon limitation with a cessation in the production of biomass and cellular lipids. This part of accumulated TAGs could be synthesized through recycling and transformation of other lipids such as glycolipids and phospholipids. Additionally, some alterations in the fatty acid profile following TAG accumulation were found. The content of the C16:0 fatty acid increased with decreases in C16:3 and C20:5, which could have been caused by enzymatic selectivity for these fatty acids during the process of TAG synthesis. It was concluded that nitrogen and phosphorus metabolism regulates the synthesis of TAG, while carbon metabolism promotes it by providing sufficient substrates.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Microalgae are a readily available feedstock for potential sustainable and clean biofuel production due to great advantages, such as high potential oil yield, reduced emissions of CO2, value-added co-products or by-products, tolerance for adverse conditions unsuitable for conventional agriculture, etc. (Chisti 2007; Schenk et al. 2008). Since the US Aquatic Species Program was initiated in 1978 (Sheehan et al. 1998), many oleaginous microalgae species have been identified and their lipid productivities have been investigated. It has also been found that cultivation conditions have a great influence on the accumulation of microalgal lipids, especially neutral lipids. So, it is necessary to identify the possible biochemical triggers and environmental factors that might favor the accumulation of oil with less influence on microalgae growth rates (Chisti 2007).
Limitation of nutrients, especially inorganic nitrogen and phosphorus, has been shown to be able to promote the accumulation of cellular lipids in numerous species or strains of various algal taxa, such as Nannochloropsis (Yamaberi et al. 1998; Takagi et al. 2000; Rodolfi et al. 2009), Chlorella (Illman et al. 2000), Scenedesmus (Dean et al. 2010; Xin et al. 2010), Phaeodactylum (Qian and Borowitzka 1993), etc. During the process of triacylglycerol (TAG) accumulation, cellular composition changes take place, for example, the chlorophyll content decreases, phospholipids and glycolipids are hydrolyzed, photosynthetic membranes are degraded, and oil droplets are formed in microalgal cells (Liu and Lin 2001; Solovchenko et al. 2009).
During photoautotrophic cultivation, CO2 may be the only carbon resource for microalgae. The presence of high levels of CO2 by aeration has been shown to promote the production of biomass and lipids in microalgal cells (Chiu et al. 2009; Tang et al. 2011). This means that carbon and nitrogen/phosphorus metabolism have different effects on lipid metabolism. In Dunaliella viridis, lipid metabolism was found to be much more sensitive to nitrogen limitation when cells were cultured under higher CO2 (1 %) than under atmospheric CO2 levels (Gordillo et al. 1998); however, it is still unknown whether these nutrient depletions can induce modifications in the cellular lipid content and composition under carbon limitation. So it is necessary to understand the correlation effect of carbon supply and nutrient depletion on accumulation of microalgal lipids and TAGs.
Gardner et al. (2012) proposed that pH and bicarbonate could be coupled with nitrogen depletion to “trigger” TAG accumulation. And atmosphere air was bubbled instead of 5% CO2 to provide carbon limitation. However, the atmosphere still has a relatively low content of CO2. So in this study, pure nitrogen gas was aerated into the culture to break-off the carbon input from air and achieve incubation under carbon limitation. And the induction incubation of Phaeodactylum tricornutum CCMP632 was achieved in sea water without any addition of nitrogen and phosphorus. Changes in biomass, lipid contents, lipid composition, and fatty acid profiles with different carbon supply levels were investigated after the induction incubation, to understand the dependence of TAG accumulation on carbon supply.
Materials and methods
Phaeodactylum tricornutum
CCMP632, gifted by Dr. Hanhua Hu of the Institute of Hydrobiology, Chinese Academy of Sciences, was used in this study and sustained in f/2 medium (Guillard and Ryther 1962; Guillard 1975). It was kept at 25 ± 1 °C and illuminated at about 30 μmol photons m−2 s−1 by upper fluorescent lamps. Sea water was prepared by dissolution of 30 g sea salt in 1 L ddH2O and then filtered by 0.45 μm combined cellulose membrane.
Induction incubations
The inocula for induction incubation were prepared by cultivation in aerated 3-L conical flasks with a work volume of 1.5 L. The filtered air was continuously bubbled into 2f medium, which had four times additions of nutrients into sea water as regular f/2. For induction incubations, cells from logarithmic growth were harvested by centrifugation, washed three times, and suspended in artificial sea water without any addition of nutrients. The induction incubation was performed in glass columns (30 mm diameter; 28 cm long) with a working volume of 100 mL. The inoculating density was about 0.7–0.8 g L−1. The “carbon limitation” and “carbon sufficiency” conditions were achieved by continuously bubbling at about 0.1 vvm with either high-pure N2 or a 2 % CO2 gas (mixture of N2 and CO2). For the incubations with different inorganic carbon, 0, 4, 8, 20, and 40 mM NaHCO3 were added. The pure N2 gas was bubbled continuously. All of the incubations were kept at 25 ± 1 °C, and an irradiance of about 75 μmol photons m−2 s−1 provided by fluorescent lamps.
Quantifications of biomass and cell lipid content
Membrane filters were washed repeatedly in distilled water and dried to constant weight at 105 °C for subsequent use. Cells from 1 mL of the cultures were harvested by vacuum filtration and weighed on a Mettler XS105 dual-range analytical balance. The biomass content was calculated from the cell dry weight divided by the filtered culture volume.
The content of total lipids was determined according to Ben Amotz and Tornabene (1985). After the culture of about 100 mL was centrifuged and washed with ddH2O, the pellet was lyophilized and weighed as W c. Then the biomass was extracted with chloroform/methanol/1 %NaCl (2/2/1, v/v/v). Finally, the chloroform layer was collected, evaporated under N2, dried in a vacuum drying oven, and then weighed as W l. The cell lipid content was calculated as W l/W c.
Elemental analysis
The cells were harvested and washed in ddH2O by centrifugation, before lyophilizing. Then at least 3 mg of dry sample was weighed and then analyzed using a Vario EL Cube III.
Lipid composition analysis
The thin-layer chromatography (TLC) fractionation method of cellular lipids was as follows. Lipid samples were dissolved in chloroform to a concentration of 10 mg mL−1, and aliquots were spotted onto a silica gel plate (20 cm × 10 cm). Two developing solvents were utilized successively; benzene/chloroform/methanol (150:60:2, v/v/v) and benzene/hexane (50:50, v/v). After the developing steps, the plate was oven-dried at 70 °C for 2–3 min before it was stained in an air-proof cylinder with iodine.
Quantification was done using a TLC-flame ionization detection (FID) chromatography system (Iatron Laboratories, Japan) (Chen et al. 2012; Fedosov et al. 2011). Samples were dissolved in chloroform and then spotted onto Chromarod S-III silica-coated quartz rods. The rods were developed in a solvent system of chloroform/acetic acid/methanol (150:60:2:17.7, v/v/v) for the first migration to 7 cm, followed by benzene/hexane (50:50, v/v) for the second migration to 10 cm. The rods were oven-dried at 70 °C for 1 min before they were scanned in the Iatroscan analyzer operated at a flow rate of 160 mL min−1 for hydrogen and 2 L min−1 for air.
A sample of combined lipids was prepared from the mixture of four pure standards (mono-, di-, and triglyceride mix, 1787-1AMP Supelco; FAME Mix C8-C24, 18918-1AMP Supelco; mixture of sterol ester/TAG/free sterol, 10D07, Mitsubishi Chemical Analytech; soybean phosphatidylcholine, TL002301, Sinopharm Chemical Reagent Co).
Component separations of TAGs and PLs
The 1–1.5 mg of the total lipids was loaded onto a silica plate (20 × 10 cm). After the TLC fractionation, the seperated bands of TAGs and PLs were each scraped into a tube. Then the components were eluted from the silica by a mixture of methanol and chloroform (2:1). At last, the samples were dried at 60 °C under N2.
Fatty acid analysis
Preparation of methyl esters was as described by Bigogno et al. (2002). About 1 mg of lipid sample, 50 μg methyl nonadecanoate (74208-1G, Fluka) as internal standard, and 2.5 mL of 2 % H2SO4 in methanol were added into a 10-mL glass vial. This was kept at 85 °C for 2.5 h with shaking every 30 min. After cooling the vial to room temperature, 1 mL saturated NaCl solution and 1 mL heptane were added with shaking for 2 min. Finally, the supernatant after centrifuging was collected as the sample of methyl esters. For fatty acid profiling, the sample of methyl esters was analyzed by GC-MS on a Thermofisher Ultra-GC gas chromatograph coupled with a Thermofisher ITQ-1100 ion trap mass selective detector as described by Guan et al. (2011).
Statistical analysis
All experiments were done with duplicate samples and repeated at least once. In the figures, tables, and text, the mean values and the range are shown. The P value was calculated using SPSS software. All tests with a P < 0.05 were classified as having significant difference.
Results
Inhibited production of biomass and lipids under carbon limitation.
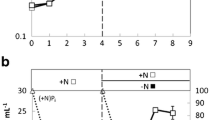
As shown in Fig. 1, after the nitrogen/phosphorus-limiting induction incubation, the biomass increased from 0.8 ± 0.01 to 1.48 ± 0.04 g L−1, and the cell increased from 34.5 ± 1.50 to 48.01 ± 1.01 % under carbon sufficiency while under carbon limitation; none of these obvious changes were identified. Therefore, during nitrogen/phosphorus depletion, sustained cell growth and accumulation of cell lipids appeared to rely totally on the fixation of external inorganic carbon in P. tricornutum CCMP632.

Biomass densities and total lipid contents in cells before and after incubations under carbon limitation and sufficiency. Data are presented as means from duplicate samples ± range
The main elemental composition in the biomass was also analyzed. As shown in Table 1, the contents of carbon (C%) and nitrogen (N%) in the inocula were, respectively, 36.50 ± 0.53 % and 3.73 ± 0.12 %. After incubation under carbon sufficiency, C% was significantly elevated to 54.37 ± 8.12 %, and N% decreased to 2.44 ± 0.39 %, while under carbon limitation, none of these alterations were identified. The carbon to nitrogen ratio (C/N) was 10.53, a little higher than 9.79 of the inocula but significantly lower than 22.30 following aeration with 2 % CO2. Under these conditions, the uptake and fixation of external inorganic carbon were effectively inhibited.
Triacylglycerol production under carbon limitation
Under nitrogen/phosphorus stress, microalgae will not only increase their cell lipid content but also modify their lipid composition; thus, lipid samples were also analyzed by TLC. As shown in Fig. 2, there was a small amount of TAG but a large amount of polar lipids (PLs) in the extracted lipids before incubation. After both incubations, an obvious band of TAGs was seen, although under carbon limitation, less was produced than under carbon sufficiency. Only under carbon sufficiency was an obvious decrease in the PL component found.

Qualitative comparative analysis of TAGs and PLs based TLC chromatography “Pre”, “Limitation”, and “Sufficiency” were respectively from the samples before and after incubations under carbon limitation and sufficiency in one set of experiment “Pre”, “0”, “4”, “8”, “20,” and “40” were respectively from the samples before and after incubations with carbon additions of 0, 4, 8, 20, and 40 mM TAG, triacylglycerols; PLs, polar lipids
To quantify these changes in lipid composition, TLC-FID was utilized. As shown in Fig. 3a, the standards could be effectively separated. As shown in Fig. 3b, c and d, cellular lipids of P. tricornutum could be classified into four categories based on different separation sequences, i.e., hydrocarbon and sterol esters (HC and SE), TAGs, diacylglycerols (DAGs), and PLs. The total lipid contents were reflected by the peak areas (Table 2). After incubation under carbon limitation and sufficiency, the TAG contents in total lipids were, respectively, 11.10 ± 3.57 % and 63.11 ± 1.85 %, both of which were significantly higher than the 0.76 ± 0.00 % of the inoculum. The same change could also be seen by the increased ratio of TAGs to PLs (TAGs/PLs). Therefore, under inorganic limitation, there is reduced but obvious TAG production after induction incubation, which might come from the transformation of other lipid components.

Chromatograms of TLC-FID analysis. (a) Mixture of lipid standards (SE: sterol ester; FAME: fatty acid methyl ester; TAG: triacylglycerol; FS: free sterol; DAG1: 1,3-diacylglycerol; DAG2: 1,2-diacylglycerol; MAG: monoacylglycerol; PC: phosphatidylcholine); before (b) and after incubations under carbon limitation (c) and sufficiency (d)
The production of lipids and TAGs with different inputs of inorganic carbon were also compared (Fig. 4). Nitrogen gas was also used for agitation and to block CO2 uptake from the air. As shown in Fig. 4, the TAG/PL ratio increased with an increasing input of inorganic carbon, leading to increased production of TAG, although 40 mM inorganic carbon did not further promote TAG production compared with 20 mM, and levels above 8 mM of inorganic carbon led to significant production of cellular lipids. Thus, inorganic carbon limitation can sustain the synthesis and accumulation of TAGs but stops lipid production under induction incubation.

Production of lipid and triacylglycerols with changing ratio of TAGs to PLs before and after incubations with carbon additions of 0, 4, 8, 20, and 40 mM. Data are means from duplicate samples ± range
Changes in the fatty acid composition
Among the fatty acids, the 16 carbon fatty acids (C16) were the largest component, under the three conditions (Fig. 5a). Next were the two of C20, C18, and C14 fatty acids in descending order. After incubation under carbon sufficiency, the five of C16 increased significantly, while C20 decreased from 23.56 ± 3.33 to 8.02 ± 0.48 %. Under carbon limitation, all of these changes were weakened or stopped. Correspondingly, C14 and C18 showed small changes after incubation, but it was surprising that, under carbon limitation, C14 increased from 6.77 ± 0.84 to 10.20 ± 0.29 %, although under carbon sufficiency, it decreased to 4.55 ± 0.56 %.

The fatty acid profiles in total lipids before and after incubations under carbon limitation and sufficiency. a The percentage of 14, 16, 18, and 20 carbon fatty acid in total; b the percentage of fatty acids with the carbon length and double bonds of C16:0, C16:1, C16:3, C18:0, and C18:1 in total. Data are means of duplicate samples ± range
Among the 16 carbon fatty acids, the fatty acids C16:0, C16:1, and C16:3 comprised the majority, but the changes after incubation were diverse. As shown in Fig. 5b, after incubation under carbon sufficiency, C16:0 increased from 17.86 ± 2.51 to 42.55 ± 1.39 %, while C16:3 decreased from 13.04 ± 2.40 to 1.19 ± 0.08 %. Even under carbon depletion, P. tricornutum still showed these minimal changes. Relatively, C16:1 showed decreased and increased proportions under carbon limitation and sufficiency, respectively. Among the 18 carbon fatty acids, C18:1 tended to accumulate, but C18:0 was decreased.
Most of these changes in the fatty acid composition were also verified by the results shown in Table 3. With increasing additions of inorganic carbon, the proportion of C16:0 and C18:1 increased, while C16:3 and C20:5 decreased. After incubation without carbon addition, the content of C14:0 was also elevated from 7.00 to 10.78 %, but with increasing addition of inorganic carbon, it gradually declined to 5.48 %. Thus, after induction incubation, P. tricornutum tended to accumulate some specific fatty acids but degraded or transformed some others along with TAG accumulation.
Fatty acid profiles in TAGs and PLs
The separate components of TAGs and PLs to be assessed by TLC were scraped from the silica plate, eluted, and utilized for fatty acid analysis. After induction incubation, there were no obvious changes identified in the fatty acid profile in either TAGs or PLs, but the selectivity of TAGs regarding the fatty acid chain was identified. As shown in Fig. 6, in the fatty acid chains of TAGs, only C14:0, C16:0, C16:1, C18:0, and C18:1 were identified, the proportions of which were, respectively, 5.50 ± 1.96, 53.22 ± 2.31, 6.13 ± 0.95, 27.05 ± 3.07, and 8.09 ± 0.75 %; in comparison, C16:0 in the PLs was 40.07 ± 1.51 %, significantly lower than in the TAGs. C16:3 and C20:5 both existed in PLs with proportions of 2.27 ± 0.70 and 4.27 ± 1.77 %, respectively. Thus, with the synthesis of TAGs, the accumulation of C16:0 and the reduction in C16:3 and C20:5 in cellular lipids could be understood.

Fatty acid profiles in triacylglycerols and polar lipids. Data are means of duplicate samples ± range
Discussion
External inorganic carbon is the most important element that influences the steady growth of microalgae. After induction incubation with nitrogen/phosphorus limitation, P. tricornutum CCMP632 still sustained cellular growth coupled with the production of lipids. However, under carbon limitation, these changes were totally inhibited, as shown in Fig. 1. It also has been reported that Nannochloropsis sp. can still maintain slow growth and rapid lipid accumulation under nitrogen depletion (Yamaberi et al. 1998). A loss of TAG accumulation was once identified due to TAG consumption under ambient aeration (Gardner et al. 2012). In the present study, N2 gas instead of air was combined with CO2, so the limited supply of O2 probably inhibited respiration, which probably led to about an 85 % increase in biomass. About 86.8 mg L−1 of lipids, on average, were synthesized de novo after 5 days of incubation. Therefore, in P. tricornutum CCMP632, lipid accumulation caused by depletion of nitrogen or phosphorus mainly comes from the fixation of external inorganic carbon and de novo synthesis, instead of carbon recycling and conversion from other metabolites.
Even under carbon limitation, there was still minimal but significant TAG accumulation identified after induction incubation in P. tricornutum. After incubation, the levels of newly synthesized TAG under carbon limitation and sufficiency were, respectively, 30.64 and 448.33 mg L−1. After incubation with carbon addition of 0 and 40 mM, these levels were, respectively, 52.94 and 218.28 mg L−1. In Scenedesmus sp. and P. tricornutum, minimal accumulation of TAGs under low carbon has also been found (Gardner et al. 2012). The synthesis of cellular TAGs is a process in which three fatty acyls are incorporated one by one into a glycerol skeleton (Wang et al. 2002). The fatty acyls can come from de novo synthesis or through fixation of inorganic carbon, and the fatty acyls existing in other cellular lipids also can be reutilized. Phospholipids and galactolipids are the main components of polar lipids in P. tricornutum (Alonso et al. 2000) and they can be hydrolyzed by phospholipase and galactolipase, respectively (Lion et al. 2006), to release fatty acids and the glycerol skeleton for subsequent synthesis of TAGs. Takagi et al. (2000) proposed that nitrogen limitation activates acyl hydrolase, which can degrade glycolipids, stimulate the hydrolysis of phospholipids, and activate diacylglycerol acyltransferase, which converts fatty acyl-CoA into TAGs.
In Chlamydomonas reinhardtii, a phospholipid/diacylglycerol acyltransferase can catalyze TAG synthesis via two pathways: transacylation of diacylglycerol (DAG) with acyl groups from phospholipids and galactolipids and DAG/DAG transacylation (Yoon et al. 2012). A galactoglycerolipid lipase has also been shown to be required for TAG accumulation following nitrogen deprivation in C. reinhardtii (Li et al. 2012). So, in P. tricornutum CCMP632, these mechanisms were all able to function to initiate the synthesis of TAGs independently of de novo synthesis of fatty acids. The degradation of the membrane lipid can also be caused by the formation of reactive oxygen species (ROS). But, in this study, the O2 supply was limited, which decreased the probability of ROS formation. This may explain why obvious TAG accumulation under carbon limitation was not followed by obvious degradation of PLs, as the results show in Fig. 2. While under carbon sufficiency, the production of O2 by photosynthesis would promote this process.
With TAG accumulation, the fatty acid profile was also changed in P. tricornutum. C16:0 accumulated, while C16:3 and C20:5 were degraded or transformed. So, the chain length and saturation of double bonds could both be adjusted, although under carbon limitation, these processes were inhibited. Fatty acid elongase and desaturase are thought to influence and control these processes in cells. Some genes encoding these enzymes have been identified and cloned in some microalgae (Meyer et al. 2004; Petrie et al. 2010). In addition, the proportion of C14:0 was found to be decreased after incubation under carbon sufficiency, while it accumulated under carbon limitation. In the psychrophilic bacterium Micrococcus cryophilus, it was found that C14 could be an intermediate of interconversion between C16 and C18 fatty acids to control phospholipid acyl chain length (Sandercock and Russell 1980). It may be possible that C14:0 could function as a precursor for synthesizing C16:0 fatty acids. So, with TAG accumulation, other fatty acids such as C20:5 can be oxidized to C14:0, but carbon limitation stops elongation.
Enzyme specificity is also thought to be a factor in the regulation of fatty acid chain length (Greenspan et al. 1970; Millar and Kunst 1997). As shown in Fig. 6, TAGs chose more C16:0 as their side chain than PLs, and more C16:3 and C20:5 accumulated in PLs. Thus, the directed alteration of fatty acid profiles may be caused by TAG synthesis and its related enzymes. Under nitrogen and phosphorus limitation, P. tricornutum tended to accumulate TAGs in cells, which could cause a series of changes.
Goldman and Graham (1981) showed that the main influence of inorganic carbon limitation appears to be not on the chemical structure of the biomass but rather on cell size; higher steady-state growth rates lead to bigger cells, although an abundant supply of CO2 was shown to be favorable for TAG accumulation. However, even with continuous aeration with 5 % CO2, TAGs only started to accumulate at the late stage of cultivation when nitrogen had been depleted (Gardner et al. 2012). Therefore, nitrogen or phosphorus metabolism regulates the synthesis of TAG, while carbon metabolism promotes it by providing enough substrates. There should be a regulating mechanism controlling this process.
References
Alonso DL, Belarbi EH, Fernandez-Sevilla JM, Rodriguez-Ruiz J, Grima EM (2000) Acyl lipid composition variation related to culture age and nitrogen concentration in continuous culture of the microalga Phaeodactylum tricornutum. Phytochemistry 54:461–471
Ben Amotz A, Tornabene TG, Thomas WH (1985) Chemical profile of selected species of microalgae with emphasis on lipids. J Phycol 21:72–81
Bigogno C, Khozin-Goldberg I, Boussiba S, Vonshak A, Cohen Z (2002) Lipid and fatty acid composition of the green oleaginous alga Parietochloris incisa, the richest plant source of arachidonic acid. Phytochemistry 60:497–503
Chen L, Liu TZ, Zhang W, Chen XL, Wang JF (2012) Biodiesel production from algae oil high in free fatty acids by two-step catalytic conversion. Bioresour Technol 111:208–214
Chisti Y (2007) Biodiesel from microalgae. Biotechnol Adv 25:294–306
Chiu SY, Kao CY, Tsai MT, Ong SC, Chen CH, Lin CS (2009) Lipid accumulation and CO2 utilization of Nannochloropsis oculata in response to CO2 aeration. Bioresour Technol 100:833–838
Dean AP, Sigee DC, Estrada B, Pittman JK (2010) Using FTIR spectroscopy for rapid determination of lipid accumulation in response to nitrogen limitation in freshwater microalgae. Bioresour Technol 101:4499–4507
Fedosov SN, Brask J, Xu XB (2011) Analysis of biodiesel conversion using thin layer chromatography and nonlinear calibration curves. J Chromatogr A 1218:2785–2792
Gardner RD, Cooksey KE, Mus F, Macur R, Moll K, Eustance E, Carlson RP, Gerlach R, Fields MW, Peyton BM (2012) Use of sodium bicarbonate to stimulate triacylglycerol accumulation in the chlorophyte Scenedesmus sp. and the diatom Phaeodactylum tricornutum. J Appl Phycol 24:1311–1320
Goldman JC, Graham SJ (1981) Inorganic carbon limitation and chemical composition of two freshwater green microalgae. Appl Environ Microbiol 41:60–70
Gordillo FJL, Goutx M, Figueroa FL, Niell FX (1998) Effects of light intensity, CO2 and nitrogen supply on lipid class composition of Dunaliella viridis. J Appl Phycol 10:135–144
Greenspan MD, Birge CH, Powell G, Hancock WS, Vagelos PR (1970) Enzyme specificity as a factor in regulation of fatty acid chain length in Escherichia coli. Science 170:1203–1204
Guan W, Zhao H, Lu X, Wang C, Yang M, Bai F (2011) Quantitative analysis of fatty acid-based biofuels produced by wild-type and genetically engineered cyanobacteria by gas chromatography–mass spectrometry. J Chromatogr A 1218:8289–8293
Guillard RRL (1975) Culture of phytoplankton for feeding marine invertebrates. In: Smith WL, Chanley MH (eds) Culture of marine invertebrate animals. Plenum Press, New York, pp 26–60
Guillard RR, Ryther JH (1962) Studies of marine planktonic diatoms. I. Cyclotella nana Hustedt, and Detonula confervacea (Cleve) Gran. Can J Microbiol 8:229–239
Illman AM, Scragg AH, Shales SW (2000) Increase in Chlorella strains calorific values when grown in low nitrogen medium. Enzyme Microb Technol 27:631–635
Lion U, Wiesemeier T, Weinberger F, Beltran J, Flores V, Faugeron S, Correa J, Pohnert G (2006) Phospholipases and galactolipases trigger oxylipin-mediated wound-activated defence in the red alga Gracilaria chilensis against epiphytes. Chembiochem 7:457–462
Liu CP, Lin LP (2001) Ultrastructural study and lipid formation of Isochrysis sp. CCMP1324. Bot Bull Acad Sinica 42:207–214
Li X, Moellering ER, Liu B, Johnny C, Fedewa M, Sears BB, Kuo MH, Benning C (2012) A galactoglycerolipid lipase is required for triacylglycerol accumulation and survival following nitrogen deprivation in Chlamydomonas reinhardtii. Plant Cell 24:4670–4686
Meyer A, Kirsch H, Domergue F, Abbadi A, Sperling P, Bauer J, Cirpus P, Zank TK, Moreau H, Roscoe TJ, Zahringer U, Heinz E (2004) Novel fatty acid elongases and their use for the reconstitution of docosahexaenoic acid biosynthesis. J Lipid Res 45:1899–1909
Millar AA, Kunst L (1997) Very-long-chain fatty acid biosynthesis is controlled through the expression and specificity of the condensing enzyme. Plant J 12:121–131
Petrie JR, Liu Q, Mackenzie AM, Shrestha P, Mansour MP, Robert SS, Frampton DF, Blackburn SI, Nichols PD, Singh SP (2010) Isolation and characterisation of a high efficiency desaturase and elongases from microalgae for transgenic LC-PUFA production. Mar Biotechnol 12:430–438
Qian KX, Borowitzka MA (1993) Light and nitrogen deficiency effects on the growth and composition of Phaeodactylum tricornutum. Appl Biochem Biotechnol 38:93–103
Rodolfi L, Chini Zittelli G, Bassi N, Padovani G, Biondi N, Bonini G, Tredici MR (2009) Microalgae for oil: strain selection, induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactor. Biotechnol Bioeng 102:100–112
Sandercock SP, Russell NJ (1980) The elongation of exogenous fatty acids and the control of phospholipid acyl chain length in Micrococcus cryophilus. Biochem J 188:585–592
Sheehan, J., Dunahay, T., Benemann, J. and Roessler, P.G. (1998) US Department of Energy’s Office of Fuels Development, July 1998. A look back at the US Department of Energy’s Aquatic Species Program—biodiesel from algae, Close out report TP-580- 24190. Golden, CO: National Renewable Energy Laboratory.
Schenk PM, Thomas-Hall SR, Stephens E, Marx UC, Mussgnug JH, Posten C, Kruse O, Hankamer B (2008) Second generation biofuels: high-efficiency microalgae for biodiesel production. Bioenergy Res 1:20–43
Solovchenko AE, Khozin-Goldberg I, Cohen Z, Merzlyak MN (2009) Carotenoid- to chlorophyll ratio as a proxy for assay of total fatty acids and arachidonic acid content in the green microalgae Parietochloris incisa. J Appl Phycol 21:361–366
Takagi M, Watanabe K, Yamaberi K, Yoshida T (2000) Limited feeding of potassium nitrate for intracellular lipid and triglyceride accumulation of Nannochloris sp. UTEX LB1999. Appl Microbiol Biotechnol 54:112–117
Tang DH, Han W, Li PL, Miao XL, Zhong JJ (2011) CO2 biofixation and fatty acid composition of Scenedesmus obliquus and Chlorella pyrenoidosa in response to different CO2 levels. Bioresour Technol 102:3071–3076
Wang JY, Zhu SG, Xu CF (2002) Biochemistry. Higher Education Press, Beijing
Xin L, Hu HY, Ke G, Sun YX (2010) Effects of different nitrogen and phosphorus concentrations on the growth, nutrient uptake, and lipid accumulation of a freshwater microalga Scenedesmus sp. Bioresour Technol 101:5494–5500
Yamaberi K, Takagi M, Yoshida T (1998) Nitrogen depletion for intracellular triglyceride accumulation to enhance liquefaction yield of marine microalgal cells into a fuel oil. J Mar Biotechnol 6:44–48
Yoon K, Han D, Li Y, Sommerfeld M, Hu Q (2012) Phospholipid:diacylglycerol acyltransferase is a multifunctional enzyme involved in membrane lipid turnover and degradation while synthesizing triacylglycerol in the unicellular green microalga Chlamydomonas reinhardtii. Plant Cell 24:3708–3724
Acknowledgments
This work was supported by the National Natural Science Foundation of China (No. 30900040) and Key Technology R&D Program of Shandong Province, China (No. 2008GG20007002).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Peng, X., Liu, S., Zhang, W. et al. Triacylglycerol accumulation of Phaeodactylum tricornutum with different supply of inorganic carbon. J Appl Phycol 26, 131–139 (2014). https://doi.org/10.1007/s10811-013-0075-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10811-013-0075-7