
Abstract
Gigartina skottsbergii is a commercially important carrageenan producer that has been suffering severe extraction pressure in Chile’s Magellan Region and Cape Horn Archipelago since 1998. In order to create baseline information for its cultivation and repopulation, we studied the effects of agricultural fertilizers on growth of G. skottsbergii early developmental stages. The culture media utilized were: a) seawater + Bayfoland, b) seawater + Superphosphate, c) seawater + Urea, d) seawater + Provasoli and e) seawater as a control. The culture conditions were: a) 12L:12D photoperiod; b) temperature 8 ± 1°C and c) irradiance at 45 µmol photons m−2 s−1. After 60 days, higher relative growth rates between treatments were observed; the treatments that included Bayfoland and Provasoli showed greater growth (382 ± 55 and 378 ± 50 µm, respectively,) compared to Superphosphate (88 ± 16 µm), control (78 ± 10 µm) and Urea (70 ± 11 µm) treatments, after 81 days. The Urea treatment and the control had inhibitory effects on G. skottsbergii germlings growth and survival, as evidenced by progressive loss of pigmentation and death after 60 days. These results showed that Bayfoland was an excellent alternative to develop cultures.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Over the last several years, there has been a global increase in the demand for red algae in the carrageenan and agar industries (Bixler 1996; Zemke-White and Ohno 1999; Buschmann et al. 2001; McNeill et al. 2003; Buschmann et al. 2004). About 20% of the world demand for carrageenan has been supplied by extractive industries in Chile (Avila et al. 2001a, b). However, these local carrageenan industries depend entirely on the harvesting of natural beds of Gigartina skottsbergii (Buschmann et al. 2001), and other carrageenan producing red algae such as Sarcothalia crispata and Mazzaella laminarioides; all are harvested as a single group, known as “lugas” (Marin et al. 2002; Avila et al. 2001a, b). Since, at the global level, the cultivation of commercially important algae represents nearly 22% of all red algae used (Zemke-White and Ohno 1999; McNeill et al. 2003), culture techniques and repopulation methods are an important alternative for the recolonization of natural algae beds to ensure the supply of primary material for the hydrocolloid industry.
Due to its high level of carrageenan content (>70% of dry weight) (Piriz and Cerezo 1991), Gigartina skottsbergii is considered to be the most important carrageenan producing red alga commercially in Chile. Export volumes of carrageenan have increased from 26 tons in 1989 to 3,772 ton in 2005, totally US$ 29.5 million in value. The Magellan Region contributes approximately 9% of the total biomass produced in Chile and has over the last five years become the third most important port of landing for the resource (Zamorano, Pers. comm.). There exists a great internal demand and significantly increasing extraction of the species, which constitutes an alternative resource for artisenal fisheries, especially during the closed seasons for sea urchin and king crab and during outbreaks of red tide, a characteristic phenomenon of the region (Avila et al. 2001b). This situation has provoked an over-exploitation and in some cases the collapse of natural populations (Avila et al. 1999; Buschmann et al. 2004; Romo et al. 2006). The growing demand for this resource and the apparent deterioration of natural populations makes evident the importance of developing a production technology to allow the acceleration of some aspects of the developmental process, such as growth (Avila et al. 1999; Romo et al. 2006). In addition, the recently declared Cape Horn Biosphere Reserve has as a primary objective the establishment of research and demonstration projects that focus on the sustainable use of biodiversity for the betterment of social wellbeing in the archipelago (Rozzi et al. 2006).
Several endeavours on the part of researchers from diverse institutions, financed through a variety of sources, have allowed the generation of background information on the ecology, biology, physiology, and cultivation of G. skottsbergii in its distribution south of 40°S (Westermeier et al. 1999; Buschmann et al. 1999, 2001; Avila et al. 1999, 2003; Romo et al. 2001, 2006; Buschmann et al. 2004; Faugeron et al. 2004; Martínez et al. 2005). However, no prior studies exist on the effects of agricultural fertilizers on the development of G. skottsbergii in controlled systems. The objective of this study, therefore, was to determine the effect of traditional agricultural fertilizers on the growth and survival rates of the initial developmental stages of G. skottsbergii under laboratory conditions, which renders production more efficient, lowers costs associated with repopulation, and develops techniques valuable for the sustainable use of this resource.
Material and methods
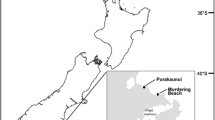
Mature tetrasporic fronds of G. skottsbergii (Fig. 1) were collected by scuba diving from natural populations at Santa Ana Point (53°18’11.4’’S; 70°23’30, 5’’W), 70 km southwest from the city of Punta Arenas on the Strait of Magellan (Fig. 2). The material was transported in seawater inside hermetic containers to the laboratory at the Center for Aquaculture and Marine Resources, Department of Natural Sciences and Resources, University of Magallanes (UMAG). Once in the laboratory the material was washed with filtered, sterilized seawater to eliminate epiphytes and epibionts that could affect the subsequent cultures.

The life history of Gigartina skottsbergii. (Mod. Romo et al. 2004)

Geographic location of collection site of Gigartina skottsbergii at Santa Ana Point, Strait of Magellan, Chile
Tetraspore release
Liberation of tetraspores was induced by stress through dehydration of the fronds at ambient temperature for approximately 45 min, after which the fronds were rehydrated in sterilized and filtered (0.2 µm) seawater. Once the tetraspores were released, the fronds were removed from the water and the resulting suspension was filtered through a sieve in order to catch the remaining tissues and impurities, obtaining a homogenous density of approximately 2,000 tetraspores mL−1 to use as an inoculation solution on glass microscope slides.
Cultivation methods used
Five treatments were performed: three commercial fertilizers (Bayfoland 250SL, Superphosphate and Urea, Table 1), a laboratory medium (Provasoli) and a control (seawater without additives). Superphosphate and Urea were used as phosphate and nitrogen sources, respectively. Both are sold in their solid state, and for this reason it was necessary to prepare stock solutions. The concentrations of each nutrient were adjusted to the corresponding values of nitrogen and phosphorus as has been used in Gracilaria chilensis (Alveal et al. 1991): 0.06 g L−1of sodium nitrate (10 mg L−1 of nitrogen) and 0.01 g L−1 of sodium phosphate (3 mg L−1 of phosphorus). Bayfoland is a liquid commercial foliar fertilizer and was used at a concentration of 0.1 ml L−1, corresponding to 1/3 of the required minimum concentration for the cultivation of initial growth stages. The quantities of different agricultural fertilizers required to achieve the indicated concentrations of nitrogen and phosphorus are shown in Table 2.
The cultures were maintained inside a temperature-controlled room at 8 ± 1°C and 45 μmol photons m−2 s−1 PAR (provided by Philips TLT 20W/54 daylight fluorescent tubes) at 12:12h light-dark cycle. The culture media for each treatment was renewed weekly.
Experimental design
Slides inoculated with tetraspores were placed in 250 mL capacity Petri dishes. Fifteen experimental units (dishes) were prepared, distributed in the 5 treatments of 3 replicates each. The effects of the distinct nutrient solutions on the development of G. skottsbergii tetraspores were evaluated through the mortality and growth of 45 individuals of each experimental unit. Weekly and during the 81 days of cultivation, 45 individuals were photographed using a microscope integrated imaging system connected to an image analyzing software (Imagen Pro-Plus version 4.1), providing data on the increase in diameter of discs of G. skottsbergii.
Growth rate and data analyses
Increase of basal diameter was used as the parameter for estimating growth rates. The relative growth rate of G. skottsbergii discs was estimated using the formula proposed by Hansen (1980):
Where:
- RGR (%):
-
relative growth rate expressed in daily percentage
- t0 :
-
initial diameter of disc (µm)
- tf :
-
final diameter of disc (µm)
- T:
-
time in days.
Subsequently, the values for the growth rates and mortality percentages were transformed by applying arc-sine (Zar 1999) to achieve normal distributions and then subjected to an analysis of variance analysis (ANOVA), using the software program Statistica 5.0. When significant differences were found, a Newman-Keul test was applied a posteriori (Zar 1999). All conclusions were based on a 95% confidence level (p < 0.05).
Results
Growth rates
Gametophytes of G. skottsbergii cultivated with different nutrient solutions showed different responses in growth during their initial developmental stages. At the end of day 81, the greatest increase in disc diameter was obtained with Bayfoland (381.87 ± 54.99 µm) and Provasoli (377.88 ± 49.92 µm). Growth was less in the treatments with Superphosphate (88.36 ± 16.27 µm), in the control solution (77.65 ± 9.93 µm) and with Urea (70.06 ± 10.88 µm) (Fig. 3).

Diameter increase (µm ± SD) of G. skottsbergii discs during 81 days of culture using different nutrient solutions
Therefore the highest growth rates were observed in the gametophytes cultivated with Bayfoland (6.19 ± 0.38% day−1) and Provasoli (5.70 ± 0.035% day−1), whereas the lowest growth rates were observed in the control and Urea treatments (3.76 ± 0.07% day−1 and 3.27 ± 0.23% day−1, respectively) (Fig. 4). The differences between all the treatments utilized were statistically significant (p = 0.000) (Table 3).

Daily growth rate (% day−1 ± SD) of G. skottsbergii discs during the first 35 days of the study in different treatments. Different letters refer to significant differences between mean values with Newman-Keuls post-hoc test (p < 0.05)
The a posteriori test applied to the interaction between the different treatments used and the time of cultivation, considering weekly periods, showed that during the first week, the gametophytes cultured with Provasoli had a higher growth rate than those under the other nutrient regimes (Fig. 5). During the second week, significant differences were observed in growth under Provasoli and Superphosphate treatments (p = 0.021) and Urea (p = 0.026) (Fig. 5, Table 4).

Growth rates (% day−1 ± SD) of G. skottsbergii disks during the first five weeks of culture (n = 3). (*) Indicates significant differences (p < 0.05)
In the third week, the differences observed between the treatments were only in Bayfoland v/s the other cultures (control, p = 0.043; Superphosphate, p = 0.028; Urea, p = 0.000) (Fig. 5, Table 4). During the fourth week, the Provasoli and Bayfoland treatments showed significant differences in growth rate related to the other treatments (Fig. 5, Table 4).
Finally, beginning in the fifth week of the study, differences in growth rates became evident, mainly between the treatments Provasoli and Bayfoland compared to control, Superphosphate and Urea treatments (Fig. 5, Table 4). The a posteriori test indicated that the differences derived mostly from the control (least growth) versus Bayfoland (p = 0.033) and Provasoli (p = 0.021), Urea versus Provasoli (p = 0.011) and Bayfoland (p = 0.016), which showed the greatest growth at the end of the study (Table 4). The variance test used on the weekly growth rates showed that the nutrient solutions used, the time under cultivation, and the interaction between them, acted in a synergistic manner on the growth of the gametophytes.
A more detailed analysis of the behavior of the growth rates of different treatments (control, Provasoli, Bayfoland, Superphosphate, and Urea) during the five weeks of the study indicated that, although the growth rates of gametophytes cultivated with Bayfoland and Provasoli did not show significantly greater growth during the first 7 days of cultivation, they began in the second and third weeks to show better developmental responses in relation to the other treatments, recording rates of greater than 8% daily growth as compared with growth rates that did not surpass 6% in the other treatments. ANOVA carried out on the growth results of the Provasoli and Bayfoland treatments, the solutions that noticeably promoted the growth of G. skottsbergii gametophytes, showed no statistically significant difference (p = 0.431) between the growth rates of these two treatments (Table 3).
Survival
Beginning with day 39 and day 46, respectively, mortality was observed in the Urea (45%) and control treatments (11%). At 67 days of cultivation, all of the gametophytes cultured under the control treatment, Superphosphate, and Urea had died. The treatments using Provasoli and Bayfoland had disc survival rates of 100% on the 81st (final) day of the experiment (p = 0.000) (Table 3). The a posteriori test indicated that the observed differences were between the treatments of Bayfoland and Provasoli (p = 1.00) and cultures enriched with Superphosphate (p = 0.000) and Urea (p = 0.000) (Table 5).
Discussion
Physiological explanations for differential response to cultivation media
There is a tendency to think of all algae as having similar nutritional requirements, but the heterogeneity of seaweeds makes such generalization difficult (Lobban and Harrison 1994). A wide variety of physiological and biological factors, such as inter-seaweed variability, nutritional history, type of tissue, life history stage, age, the surface area to volume ratio of the thallus, and morphology (Harrison et al. 1986; Harrison and Hurd 2001), will influence growth and uptake of nutrients.
Nitrogen and phosphorus are two nutrients that limit seaweeds growth and yields in most natural environments (Harrison and Hurd 2001). Nitrate and ammonia are important environmentally available nitrogen sources for the growth of marine algae. After these ions are absorbed from the environment, they are generally incorporated into amino acids and proteins, or used in the synthesis of pigments such as phycoerithrin, which is considered to be a store of available nitrogen (Lapointe 1985; Ryther et al. 1981). However, in many marine algae, nitrogen in the form of ammonia is frequently absorbed and assimilated more rapidly than nitrate and Urea (De Boer 1981). At the same time, it has also been found that Urea is an excellent source of nitrogen for many marine algae (Nars et al. 1968; Probyn and Chapman 1982; Thomas et al. 1985). However, experiments carried out by Buschmann et al. (2004), showed that G. skottsbergii achieves most of its growth rate with the addition of ammonia, rather than nitrate.
In our experiments, G. skottsbergii germlings grew more rapidly when cultivated in a solution of Bayfoland 250 SL and Provasoli, while growth rates in Urea and Superphosphate were significantly lower. These differences may be due to the chemical composition of each of the nutrient solutions used. A comparison of the solutions of Urea and Superphosphate with those of Bayfoland and Provasoli reveals that the latter solutions contain a broad variety of macro and micronutrients, namely all those required for the development of plants. In fact, the Provasoli solution was formulated specifically for the enrichment of media for cultivation of seaweeds and likely for this reason produced favorable growth in comparison with Urea and Superphosphate. On the other hand, Bayfoland has been formulated for the growth and development of terrestrial plants, and contains macro (N, P and K) and micronutrients (Fe, Cu, Mo, Ni, Mn and Bo among others), thus preventing nutritional problems in plants and making it also an effective medium for the cultivation of G. skottsbergii.
For their part, Urea and Superphosphate provide N (46%) and P (48%). Urea is an excellent N source for some seaweeds (e.g., kelp), but other seaweeds show reduced growth on Urea (De Boer 1981; Navarro-Angulo and Robledo 1999). While it is true that P and N are considered essential limiting elements for growth of algae (Lobban and Harrison 1994), in this study, their addition via Urea and Superphosphate did not increase growth rates in the G. skottsbergii germlings, possibly due to the absence of other elements which are present in Bayfoland and Provasoli.
This decrease in growth in two treatments, even in the presence of limiting nutrient, could be due to saturation of the uptake capacity for N and P. Some seaweeds do not show such saturation at high environmental nutrient concentration (as described by the Michaelis-Menten Equation), and their uptake rate increases linearly with high nutrient additions (Harrison et al. 1986; Taylor et al. 1998). However, here G. skottsbergii may need other elements and nutrients or have difficulty absorbing and assimilating Urea and Superphosphate. Nitrate is not as easily assimilated as ammonia, and there is a waste of energy in its capture and assimilation (Hurd 2000). Also, since our experiments were done without aeration, it must be considered that water motion is extremely important for seaweeds in natural conditions, since it determines the thickness of velocity and diffusion boundary layers (DBL’s) around the thallus and hence the movement of ions and gases to and from the thallus surface (Hurd 2000). Nonetheless, an adequate nutrient supply is important in the early stages (germlings) of G. skottsbergii development, as it determines future growth and reproduction processes.
References
Avila M, Candia A, Núñez M, Romo H (1999) Reproductive biology of Gigartina skottsbergii (Gigartinaceae, Rhodophyta) from Chile. Hydrobiologia 398/399:149–157
Avila M, Pavez H, Candia A, San Martín R, Cáceres J (2001a) Effects of temperature and Irradiance on the germination and growth of Sarcothalia crispata in southern Chile. In: Chapman ARO, Anderson RJ, Vreeland VJ, Davison I (eds) Proceedings of the 17th International Seaweed Symposium. Oxford Univ Press, pp 27–33
Avila M, Candia A, Romo H, Pavez H, Torrijos C (2001b) Exploitation and cultivation of Gigartina skottsbergii in southern Chile. In: Chapman ARO, Anderson RJ, Vreeland VJ, Davison I (eds) Proc 17th International Seaweed Symposium. Oxford Univ Press, pp 137–143
Avila M, Candia A, Romo H, Pavez H, Torrijos C (2003) Exploitation and cultivation of Gigartina skottsbergii in southern Chile. In: Chapman ARO, Anderson RJ, Vreeland VJ, Davison IR (eds) Proc 17th International Seaweed Symposium, pp. 137–143
Bixler JH (1996) Recent developments in manufacturing and marketing carrageenan. Hydrobiology 362/327:35–57
Buschmann AH, Correa JA, Westermeier R (1999) Recent advances in the understanding of the biological basis for Gigartina skottsbergii Rhodophyta cultivation in Chile. Hydrobiologia 398/399:427–434
Buschmann AH, Correa J, Westermeier R, Hernandez M, Norambuena R (2001) Red algal farming in Chile: a review. Aquaculture 194:203–220
Buschmann AH, Varela D, Cifuentes M, Hernandez Gonzalez MC, Henrıquez L, Westermeier R, Correa JA (2004) Experimental indoor cultivation of the carrageenophytic red alga Gigartina skottsbergii. Aquaculture 241:357–370
Conway HL, Harrison P, Davis CO (1976) Marine diatoms grown in chemostats under silicate or ammonium limitation. II. Transient response of Skeletonema costatum to a single addition of the limiting nutrients. Mar Biol 35:187–199
De Boer JA (1981). Nutrients. In: Lobban CS, Wynne MJ (eds) The Biology of seaweeds. Oxford Blackwell Scientific, pp 356–391
Faugeron S, Martinez EA, Correa J, Cardenas L, Viard F, Destombe C, Valero M (2004) Reduced genetic diversity and increased population differentiation in peripheral and overharvested populations of Gigartina skottsbergii (Rhodophyta, Gigartinales) in southern Chile. J Phycol 40:454–462
Hansen JE (1980) Physiological considerations in the mariculture of red algae. In: Abbott IA, Foster MS, Eklund LF (eds) Pacific Seaweed Aquaculture. Publications of the California Sea Grant. Coll. Program, La Jolla, pp 80–91
Harrison PJ, Hurd CL (2001) Nutrient physiology of seaweeds: application of concepts to aquaculture. Cah Biol Mar 42:71–82
Harrison PJ, Druehl LD, Lloyd KE, Thompson PA (1986) Nitrogen uptake kinetics in three year-classes of Laminaria groenlandica (Laminariales: Phaeophyta). Mar Biol 93:29–35
Hein M, Pedersen MF, Sand-Jensen K (1995) Size-dependent nitrogen uptake in micro and macroalgae. Mar ecol Prog ser 118:247–253
Hurd CL (2000) Water motion, marine macroalgal physiology and production. J Phycol 36:453–472
Lapointe B (1985) Strategies for pulse nutrient supply to Gracilaria cultures in the Florida Keys: interactions between concentration and frequency of nutrient pulses. J Exp Mar Biol Ecol 93:211–221
Lobban CS, Harrison PJ (1994) Seaweed Ecology and Physiology. Cambridge Univ Press, Cambridge, p 366
Marin SL, Westermeier R, Melipillan J (2002) Simulation of alternative management strategies for red algae, luga roja, (Gigartina skottsbergii Setchell and Gardner) in southern Chile. Ecol Mod 154:121–133
Martínez EA, Correa J, Faugeron S, Mansilla A, Avila M, Camus P (2005) Demographic and genetic survey of the red seaweed Gigartina skottsbergii Setchell et Gardner (Rhodophyta, Gigartinaceae) along its distribution range in the southern Pacific. Cienc Tecnol Mar 28(1):53–63
McNeill SE, Page M, Falshaw R (2003) Field trials to optimize the timing and frequency of pruning for cultivation of a New Zealand carragenophyte, Gigartina atropurpurea. J Appl Phycol 15:391–405
Navarro-Angulo L, Robledo D (1999) Effects of nitrogen source, N: P ratio and N-pulse concentration and frequency on the growth of Gracilaria cornea (Gracilariales, Rhodophyta) in culture. Hydrobiologia 398/399:315–320
Nars AH, Bekheet I, Ibrahim RK (1968) The effects of different nitrogen and carbon source on amino acids synthesis in Ulva, Dictyota and Pterocladia. Hydrobiologia 31:7–16
Pedersen MF (1994) Transient ammonium uptake in the macroalgae Ulva lactuca (Chlorophyta): Regulation, and the consequences for choice of measuring technique. J Phycol 30:980–986
Piriz M, Cerezo A (1991) Seasonal variation of carrageenans in tetrasporic, cystocarpic and “sterile” stages of Gigartina skottsbergii S. et G. (Rhodophyta, Gigartinales). Hydrobiology 226:65–69
Probyn TA, Chapman ARO (1982) Nitrogen uptake characteristics of Chordaria flagelliformis (Phaeophyta) in batch mode and continuous mode experiments. Mar Biol 71:129–133
Romo H, Paula E (1995) Métodos experimentales para el cultivo de Porphyra. In: Alveal K, Ferrario M, Oliveira y E, Sar E (eds) Manual de Métodos Ficológicos. Universidad de Concepción, Editorial Anibal S.A., Chile, pp 551–562
Romo H, Avila M, Candia A (2001) Manual de Técnicas de Cultivo y Repoblación de “Luga Roja” (Gigartina skottsbergii). Proyecto FONDEF D97I1064. Universidad de Concepción-IFOP, Chile, p 32
Romo H, Avila M, Candia A (2004) Manual de Técnicas de Cultivo y Repoblación de “Luga Roja” (Gigartina skottsbergii). Proyecto FONDEF D97I1064 y DOOI1109. Universidad de Concepción-IFOP, Chile, p 32
Romo H, Avila M, Nuñez M, Perez R, Candia A, Aroca G (2006) Culture of Gigartina skottsbergii (Rhodophyta) in southern Chile. A pilot scale approach. J Appl Phycol 18:307–314
Rozzi R, Massardo F, Anderson C, Berghoefer A, Mansilla A, Mansilla M, Plana J, Berghoefer U, Barros E, Araya P (2006) La RB Cabo de Hornos. Ediciones de la Universidad de Magallanes, Punta Arenas, Chile, p 260
Ryther JH, Corwin N, DeBusk TA, Williams TD (1981) Nitrogen uptake and storage by the red alga Gracilaria tikvahiae (McLachlan, 1979). Aquaculture 26:107–115
Taylor RB, Peek JT, Rees TAV (1998) Scaling of ammonium uptake by seaweeds to surface area: volume ratio: Geographical variation and the role of uptake by passive diffusion. Mar Ecol Prog Ser 169:143–148
Thomas TE, Harrison PJ, Taylor EB (1985) Nitrogen uptake and growth of the germlings and mature thallus of Fucus distichus. Mar Biol 84:267–272
Vallentinus I (1984) Comparisons of nutrients uptake rates for Baltic macroalgae with different thallus morphologies. Mar Biol 80:215–225
Westermeier R, Aguilar A, Sigel J, Quintanilla J, Morales J (1999) Biological basis for the management of Gigartina skottsbergii Gigartinales, Rhodophyta in southern Chile. Hydrobiologia 398/399:137–147
Zar JH (1999) Biostatistical analysis, 4th edn. Prentice- Hall, Inc, New Jersey, p 663
Zemke-White WL, Ohno M (1999) World seaweed utilization: An end-of-century summary. J Appl Phycol 11:369–376
Acknowledgements
We gratefully acknowledge Dr. Christopher Anderson for valuable suggestions and improvements of this manuscript. The manuscript also has been improved with the suggestions of two anonymous reviewers. This work was financed in part by Project PRN 00251 (University of Magallanes) to AM and Project Fondef D03I1164 and Project 010201010085 (University Arturo Prat) to MA.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Mansilla, A., Palacios, M., Navarro, N.P. et al. Growth and survival performance of the gametophyte of Gigartina skottsbergii (Rhodophyta, Gigartinales) under defined nutrient conditions in laboratory culture. J Appl Phycol 20, 889–896 (2008). https://doi.org/10.1007/s10811-007-9279-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10811-007-9279-z