
Abstract
Recent human population growth in North America’s southern Appalachian Mountains has raised concerns about the effects of ex-urban development on its streams and their at-risk biota. The few studies to examine effects of land-use change in this region found that many headwater species require well-forested watersheds. Alasmidonta raveneliana is endemic to montane streams in North Carolina and Tennessee and during the past 20 years populations have declined alarmingly in several former strongholds. In 2015, we conducted surveys for A. raveneliana and quantified in-stream habitat and land-use at 25 current or historically occupied sites in the Nolichucky Drainage. We found that A. raveneliana populations were only detected at sites with well-forested watersheds and low proportions of fine substrates. Thin-sections revealed that A. raveneliana is fast-growing and short-lived (max age ~ 12 years) which may be an adaptation to unpredictable headwater streams. However, like many headwater specialists, A. raveneliana appears sensitive to changes in forest cover. Our results suggest that subtle changes in stream habitats linked to land-use change are a significant threat to the long-term survival of this species. Protection of forested headwaters and restoration of riparian zones appear to be the best strategies for conserving populations of this critically endangered freshwater mussel.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Stream ecologists have long noted the importance of interactions between aquatic systems, associated terrestrial landscapes and their role in the persistence of sensitive stream biota (Ellis, 1936). Changes to land-use and land cover (LULC) at both the watershed and reach scales may strongly influence stream hydrologic and thermal attributes as well as in-stream habitat and channel geomorphology (Harding et al., 1998; Gergel et al., 2002; Hopkins & Roush, 2013). The spatial extent and frequency of LULC disturbance may also profoundly affect the composition of benthic invertebrate and fish communities (Snyder et al., 2003; Allan, 2004; Pan et al., 2004; King et al., 2005; Ahern et al., 2005; Weijters et al., 2009; Simeone et al., 2021). Thus mitigating the impacts of disturbance and development has become an important tool in the toolbox of conservation managers.
Freshwater mussels are acutely sensitive to changes in land-use and stream habitat conditions (Ellis, 1936; Brim-Box & Mossa, 1999; Hopkins, 2009). Sedimentation from forestry, agriculture, or other disturbances may occlude interstitial spaces in heterogeneous gravel riffles and runs resulting in the formation of hardpan streambeds (Gordon et al., 2004). Furthermore, fines may foul the gills and other feeding structures of mussels and other benthic filter feeders (Ellis, 1936; Kat, 1982; Aldridge et al., 1987). Clearing of upstream forestlands often results in changes to sediment dynamics and flow regimes in downstream reaches. Fine sediments alter freshwater mussel habitats at both the micro- and macro-scale. Hopkins (2009) found that altered LULC may supersede the effects of local surface geology and is contributing to mussel extirpations in southern Appalachian watersheds. Although altered sediment composition can lead to dramatic changes in the size and distribution of mussel populations, the effects of sedimentation may take years or decades to become noticeable (Brim-Box & Mossa, 1999). Additionally, it is difficult to link aquatic habitat degradation to land-use change because impacts of landscape-level disturbances may take decades to become detectable (Brim-Box & Mossa, 1999; Allan, 2004; Maloney et al., 2008).
Episodic flooding and associated channel alteration are linked to LULC changes in many systems and are important stressors for many benthic organisms, including mussels. Moreover, the effects of geomorphic change are evident at multiple scales (DiMaio & Corkum, 1995; Gangloff & Feminella, 2007). Many benthic organisms survive the scouring effects of spates by utilizing hydraulic refugia that experience lower-intensity shearing and turbulent forces and exhibit higher levels of substrate stability relative to other parts of the stream (Vannote & Minshall, 1982; Lancaster & Hildrew, 1993; Strayer & Ralley, 1993; Strayer, 1999; Fuller et al., 2010; Pandolfo et al., 2010). However, adult freshwater mussels are relatively sedentary and are generally unable to move quickly enough to avoid high-flow events. Conversely, long-distance displacement of mussels by high-flow events frequently results in burial or lethal stranding (Vannote & Minshall, 1982; Brim-Box & Mossa, 1999; Hastie, 2011). Moreover, mussels may become stranded on sand bars where they are subject to high rates of predation or desiccation when waters recede (Brown & Banks, 2001). Similarly, deep burial by mobile sediments may also result in mass mortalities of mussels as is evidenced by fossil and sub-fossil ‘death assemblages’ (i.e., evidence for mass burial of large aggregations of freshwater bivalves in riverine strata; Vannote & Minshall, 1982; Cummins, 1994).
Streams draining the southern Appalachian Mountains are among North America’s most ancient and unique freshwater ecosystems (Lydeard & Mayden, 1995; Benz & Collins, 1997; Parmalee & Bogan, 1998; Williams et al., 2017). Much of the region was heavily logged in the late 19th and early twentieth centuries with devastating impacts to streams, and presumably wildlife (Glenn, 1911). Over the past century, however, forests across the southern Appalachians in western North Carolina, eastern Tennessee, northern Georgia, and South Carolina have regenerated (sensu Harding et al., 1998). Although the region has seen increased urban and ex-urban development (Paul & Meyer, 2001; Scott, 2006), secondary forest cover remains high thanks to a large network of National Parks (e.g., Great Smokey Mountains NP), National Forests (e.g., Pisgah, Nantahala, Cherokee NFs) and state-managed lands (e.g., state forests, wildlife management areas). These protected headwaters support a large number of endemic, imperiled species, and as such play a critical role in numerous state and federal endangered species management plans (USFWS, 2002, 2017). Although numerous studies have examined the effects of changing LULC on many southeastern US Piedmont and Coastal Plain streams, there have been surprisingly few studies examining how recent changes to LULC have affected montane streams in the southern Appalachian Mountains (but see Hopkins, 2009; Shea et al., 2013).
Here we examine how variation in land-use and in-stream habitat parameters influence the distribution, abundance, and demographic structure of Appalachian elktoe [Alasmidonta raveneliana (I.Lea, 1834)], a federally endangered freshwater mussel. We tested the hypothesis that A. raveneliana distribution and site occupancy are related to land-use at multiple scales. We also examined the role of both landscape and reach scale physical stream habitat parameters (e.g., substrate, flow) in order to attain insights into interactions among these factors that contribute to mussel occurrence within a Tennessee River Drainage headwater river network, the upper Nolichucky Drainage. These observations may help inform subsequent habitat conservation and management strategies designed to mitigate ongoing declines of A. raveneliana and help guide management and recovery of this endemic freshwater mussel. Additionally, A. raveneliana is the furthest upstream occurring mussel species found in the Nolichucky Drainage. Protection and conservation of this species and its unique habitat requirements would contribute to the enhancement and protection of other endemic fauna and natural habitat downstream.
Methods
Focal species
Alasmidonta raveneliana is endemic to upper Tennessee River Drainage streams draining the Blue Ridge Physiographic Province in western North Carolina and eastern Tennessee (Clarke, 1981). Alasmidonta raveneliana was listed as an endangered species under the US Endangered Species Act in 1994 because it occurred primarily as small, isolated populations across a restricted range (USFWS, 1994). During the latest 5-year review conducted by the US Fish and Wildlife Service (Service) A. raveneliana was considered to be declining (USFWS, 2017). Seven isolated populations currently occur in 10 streams in the French Broad and Little Tennessee River drainages in North Carolina. However, only 5 of these populations are believed to be stable and reproducing (USFWS, 2017). The Nolichucky River and its tributaries the Cane, North Toe, South Toe, and Toe rivers support one of the largest populations of A. raveneliana in the French Broad Drainage. Large populations are also found in the Tuckasegee, the Pigeon, and the Little River in western North Carolina. The Little Tennessee River historically contained the largest populations prior to ~ 2005 but they are now largely extirpated from that system (USFWS, 2017). Hypothesized mechanisms for the collapse of the Little Tennessee population include land use-mediated changes to water temperature and quality, the effects of invasive species, disease, flooding associated with remnants of hurricanes and climate change (Fraley & Simmons, 2006; Jarvis, 2011; Levine et al., 2015; Pandolfi, 2016; USFWS, 2017).
Study sites
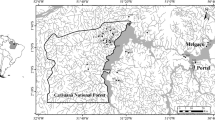
The Nolichucky River drains ~ 2921 km2 (~ 1600 km2 in North Carolina) of primarily upland habitat in western North Carolina and eastern Tennessee. Its headwaters originate on the slope of Mount Mitchell (elevation 2037 m), the highest point in North America east of the Mississippi River. The Nolichucky River is formed by the confluence of the mainstem Toe and Cane rivers ~ 13 km northwest of Burnsville, North Carolina (elevation ~ 860 m). Along with its primary tributaries, the Cane, North and South Toe rivers, the Nolichucky drains portions of the Blue Ridge Physiographic Province in western North Carolina. The region is largely rural. Much of the headwaters are steep and forested but small agricultural and mining communities are found in the lowlands. According to 2011 LULC statistics, ~ 84% of the watershed was forested, 6.7% was urban and impervious surfaces, and 7.6% was cropland (www.streamstats.usgs.gov). Study sites were located within the North Carolina counties of Yancey, Mitchel, and Avery (Fig. 1). According to the 2019 U.S. Census, human populations in the three counties in the upper Nolichucky watershed were 18,609 in Yancey County, 14,964 in Mitchel County, and 17,577 in Avery County (www.census.gov/quickfacts/map).

Map of study localities in the Nolichucky drainage in North Carolina. Solid circles represent sites with extant A. raveneliana populations, whereas open circles indicate non-occupied sites during 2015 surveys
Alasmidonta raveneliana was historically present at or had the potential to occupy all of our study sites in the Nolichucky Drainage; however, few historical (i.e., pre-1990) mussel survey data are available for this drainage. Only one other native freshwater mussel, the Wavy-rayed lampmussel (Lampsilis fasciola Rafinesque, 1820) is known from this drainage. The Asian clam [Corbicula fluminea (O.F.Müller, 1774)] also occurs at very low densities in the lower South Toe River and at moderate densities in the lower Cane, North Toe, mainstem Toe, and Nolichucky rivers (GP, personal observation).
Mussel surveys
We used timed searches to quantify freshwater mussel abundance at 25 sites across the upper Nolichucky River Drainage in North Carolina (Table 1, Fig. 1). At each site, we established a 150-m study reach that was divided into 10-m sections (n = 15 sections per site). Experienced personnel searched the entire wetted area of each 10-m section using visual-tactile survey methods (i.e., searchers examined the streambed via snorkeling and lightly disturbed finer substrates to a depth of 10–15 cm) to detect any mussels present. We conducted surveys during summer months (e.g., June–August) when A. raveneliana are believed to be actively feeding and located at the surface of the streambed. Mussels were identified and measured (total length) before being returned to their point of capture (Fig. 2). Mussel surveys were conducted with assistance from biologists with the North Carolina Wildlife Resources Commission (NCWRC), the US Fish and Wildlife Service (Service), and the US Forest Service (USFS).

Size distribution of 327 A. raveneliana encountered alive at 12 of 25 sites in the Nolichucky River Drainage during 2015 surveys. The mean total length of mussels was 57.5 mm and ranged from 21 to 87 mm
In-stream habitat
We quantified physical habitat parameters at summer low-flow conditions along 16 cross-channel transects within each study site. We used a Marsh McBurney Flow Mate (HACH, Loveland, CO) flow meter to quantify stream depth and mid-water column current velocity at 5 equidistant increments across each transect (n = 80 depth and flow measurements per site). We used a modified Wolman Pebble count (Wolman, 1954) to quantify substrate composition and collected 25 randomly selected substrate particles along each cross-channel transect (n = 400 substrate measurements per site). We recorded the maximum diameter of all lithic substrate particles and characterized non-lithic substrates including wood, organic matter, bedrock, boulder [i.e., particles > 2 m diameter], silt, and sand. To control for uncertainties associated with differentiating small diameter particles in the field, we summed all of the sand and silt particles detected to obtain an estimate of the proportion of fine sediments (< 2 mm) present within each site.
We estimated bank-full channel depth using a telescoping stadia rod. Measurements were made from the surface of the base flow water level to the height of the lowest stream bank. Nine bank-full height measurements were taken at each site and we attempted to get measurements at the upstream and downstream ends of the site as well as mid-site (Gangloff & Feminella, 2007). The mean bank-full depth for each site was estimated by adding the mean height of stream banks to the mean channel water depth under base flow conditions.
Land-use analysis
Land-use based on 1992 and 2011 data were computed for both the entire catchment upstream of each study site as well as the riparian zone within 100-m of the active channel for the entire upstream catchment (Hopkins & Roush, 2013). We also calculated the change in land-use from 1992 to 2011 for all (n = 25) study sites at the same catchment and riparian scales using ArcGIS (v10.3; ESRI Redlands, CA) and the ArcHydro toolset following a slightly amended protocol from Merwade (2012). In contrast to Merwade (2012), we used a much smaller (n = 1000) cell convergence requirement to initiate streams. The smaller threshold was selected to better represent first order streams in the drainage networks (Pandolfi, 2016). National Land Cover Data were downloaded from the Multi-Resolution Land Characteristics Consortium (www.mrlc.gov).
Drainages were delineated from a 30-m resolution (i.e., each 30 × 30 m cell equals a single land-use type) digital elevation models (DEMs) downloaded from Earth Explorer (www.earthexplorer.usgs.gov). US Geological Survey land-use categories were simplified prior to statistical analysis. We combined deciduous, evergreen and mixed forest classifications into a single category representing forest cover. US Geological Survey categories for low, moderate, and highly disturbed areas were combined into one category, disturbed land cover. The agricultural land-use category used in this analysis was calculated by combining coverage data for row crop and pasture/hay land-use classes (Pandolfi, 2016). Therefore, our analysis contained three land-use categories: forest cover, disturbed land cover, and agriculture land cover.
The USFWS generated a seamless statewide North Carolina DEM by aggregating individual 20 ft resolution DEMs which were obtained from the North Carolina Floodplain Mapping Program (https://flood.nc.gov/ncflood/mappingprogram.html). The resulting statewide DEM had a pixel size of 6.1 m2 and elevation was represented at 1.27 cm intervals. To determine slope, a satellite image of the study area was overlain on the DEM in ArcGIS v10.3.1 (Esri, Redlands CA 2015) and the Interpolate Line Tool was used to determine the slope down the center of each 150-m survey reach (Pandolfi, 2016). We computed stream power by multiplying the average stream bank-full height (estimated stream bank-full + stream base flow depth) by the stream gradient obtained via ArcGIS for each site (Gordon et al., 2004; Gangloff & Feminella, 2007).
Shell sectioning
Alasmidonta raveneliana shells (n = 89) were collected opportunistically during mussel surveys in 2014 and 2015 at several sites in the South Toe River. Because A. raveneliana is a federally endangered species, we were unable to sacrifice living individuals and instead used fresh-dead shells. Shells in good condition (i.e., those with intact periostracum and nacre layers) indicate that these animals likely died within the previous year as average alkalinity levels are low (Ca2+ = 4.03 mg/l) and shells quickly dissolve in this stream (USEPA, 2018). Thin-sectioning methods followed previously published methodology (Neves & Moyer, 1988; Haag & Commens-Carson, 2008; Singer & Gangloff, 2011; Pandolfi, 2016). Shells were independently aged by two reviewers (GP and MG) using an Olympus SZ61 0.67x-4.5 × stereo-microscope. Shells for which different age estimates were obtained were averaged prior to length-at-age analysis.
Statistical analyses
We recorded the total numbers of mussels and search times for each 10-m section surveyed. We then calculated catch-per-unit-effort (CPUE) for each site by dividing the number of mussels by the total search time (person hours) per section. Additionally, we calculated frequency of occurrence (FOO) for A. raveneliana as the proportion of sections within each site in which mussels were detected. We used total number of mussels, mussel CPUE and mussel FOO as dependent variables in subsequent GLM models.
Statistical analyses were conducted in SPSS software (IBM Corp. Version 22, Armonk NY) and GNU R (R Development Core Team, 2008). We computed site-scale means for all habitat parameters and used these values in models (Table 2). We used a principal component analysis (PCA) to identify co-linear habitat variables and reduce the number of parameters in generalized linear models (GLMs). We examined the ability of stream habitat variables that loaded strongly on PC axes to independently predict A. raveneliana presence in GLMs. All possible interactions between PCs and A. raveneliana population attributes were investigated using GLMs. Spearman correlations were used to investigate links between forest cover at the watershed and riparian scales, fine sediment coverage and A. raveneliana abundance (as FOO).
We used the von Bertalanffy growth equation to create a growth curve for shells collected from the South Toe River (von Bertalanffy, 1938; Anthony et al., 2001) using GNU R and the FSA: Fisheries Stock Analysis R package version 0.8.6 (R Core Development Team, 2008; Ogle, 2016). The von Bertalanffy growth equation \((L_{t} = L_{\infty } (1 - e^{{ - K(t - t_{{\text{o}}} )}} )\), where Lt is the shell at a given age (t), L∞is the theoretical shell length-at-age infinity, K is a fitted constant showing the rate of Lt approaching L∞ over time, and to is the theoretical age when the shell length is equal to 0), was used to determine the length at age and for 89 A. raveneliana shells ranging in length from 34.5 to 91 mm and we used a length-at-age infinity estimate of 109.4 mm.
Results
Alasmidonta raveneliana distribution and abundance
We found a total of 327 A. raveneliana and 29 Lampsilis fasciola during 2015 surveys in the Nolichucky Drainage. Living mussels and A. raveneliana were detected at roughly half (12) of the 25 sites sampled (Fig. 1, Table 1). Alasmidonta raveneliana populations were found at all 6 sites surveyed in the South Toe River and at 3 sites in both the Cane and mainstem Toe Rivers. No living mussels or shells were found at 6 sites in the North Toe River. The vast majority (> 94%) of individual A. raveneliana encountered during 2015 surveys were detected in one stream, the South Toe River and 217 (66% of all A. raveneliana detected) were encountered at a single South Toe site.
Habitat parameters
Principal component analyses revealed that four PCs explained 77% of the total variation in physical habitat conditions among sites in the Nolichucky River Drainage (Table 2). Habitat PC1 explained 33.9% of overall habitat variation. Stream depth, width, and % boulder loaded positively on PC1, whereas organic matter loaded negatively. PC2 explained 20.1% of physical habitat variation. Mean velocity and % wood loaded negatively on PC2, whereas median substrate and % bedrock and width loaded positively on PC2. Habitat PC3 explained 12.7% of variation in the dataset with stream power and the proportion of boulders loading positively, whereas mean velocity and bedrock had negative loadings on PC3. Finally, PC4 explained only 10.3% of habitat variability and stream power, mean velocity, and % bedrock loaded positively and substrate size loaded negatively on this component (Table 2).
Land-use, habitat, and mussels
The frequency of A. raveneliana occurrence within transects was unrelated to land-use at the watershed scale but was significantly positively correlated with forest cover within riparian zones (i.e., 0–100 m from the stream edge, P < 0.0001, rs = 0.71, n = 25, Fig. 3). Additionally, concentrations of fine sediments were significantly higher at sites where A. raveneliana was not detected compared to sites where they were present (P = 0.011, n = 25, Fig. 4). Habitat PC2 (substrate size, proportion bedrock, proportion wood) was significantly associated with the presence of A. raveneliana (rs = 0.46, P = 0.022, n = 25). The percent of fine substrate present within a site was negatively related to watershed-scale forest cover (rs = − 0.54, P = 0.006, n = 25, Fig. 5). Further, A. raveneliana presence was significantly higher at sites with lower levels (< 30%) of fine sediment (< 2 mm, P < 0.011, n = 25, Fig. 6) suggesting that broad-scale land-use may influence the overall availability of fines within a sub-catchment but that local substrate and habitat conditions are still ultimately dependent on levels of riparian disturbance in the Nolichucky Drainage.

Scatter plot of the relationship between percent 2011 riparian forest cover and the frequency of occupied (FOO) sections (n = 16 per site) within the 150-m focal area in which A. raveneliana were detected (Spearman correlation, P < 0.0001, r = 0.711, n = 25)

Comparison of mean (± 95% confidence interval) land-use PC2 scores for sites where A. raveneliana was absent and present in the Nolichucky River Drainage during 2015 surveys. Streams with wider channels, higher flow velocities, larger median substrate diameters more bedrock, and fewer fines or organic matter were more likely to support A. raveneliana compared to streams with lower flows, smaller particles, and more organic matter

Relationship between the percent of 2011 forest cover at the watershed scale and the percentage of fine substrates (< 2 mm diameter) observed during Wolman pebble counts at sites in the Nolichucky Drainage in 2015 (Spearman correlation, P < 0.006, r = − 0.537, n = 25)

Relationship between A. raveneliana presence and the percent of fine substrates (< 2 mm diameter) at sites in the Nolichucky Drainage in 2015. Sites with lower amounts of fine sediment (silt + sand) are significantly more likely to have A. raveneliana present (P < 0.011, n = 25). Error bars represent 95% confidence intervals
Regression analysis
Generalized linear regression models found that Habitat PC1 and PC2 produced the most informative model of A. raveneliana presence (AICc = 32.37, Wi = 0.40, X2 = 9.39, P = 0.01, Table 3). Although PC1 primarily explains variation in stream size, PC2 loaded strongly on stream habitat variables appearing to be more positively associated with A. raveneliana presence. The second most informative model included PC2 alone predicting A. raveneliana presence (AICc = 34.36, Wi = 0.15, X2 = 4.81, P = 0.03, Table 3).
Shell thin-sections
Analysis of length-at-age data indicates that A. raveneliana are short-lived and fast-growing mussels. The mean age of sectioned mussels was 5.6 years and the median was 5.5 years. Shell lengths begin to asymptote at around age 8 and annual growth rates for all mussels ages 8 + appeared relatively slow. The youngest shells sectioned were 3 years old. The mean length of age 3 shells was 49.4 mm and the smallest age 3 shell was 34.5 mm long. No animals with ages > 11 y were encountered and the largest individual available for thin-sectioning was 91 mm (Fig. 7). Erosion of larger shells precluded reliable age and growth analysis but this is near the maximum reported size for Appalachian elktoe.

von Bertalanffy length-at-age data representing A. raveneliana shells thin-sections collected during 2014–2015 from 4 sites in the South Toe River (n = 89 shells). Shells ranged in length from 34.5 to 91 mm and a length-at-age infinity estimate of 109.4 mm was used
Discussion
Results of this study suggest that A. raveneliana populations, like those of many headwater specialists, are dynamic and extremely sensitive to riparian and catchment-scale land-use changes. Although the proportion of forest cover was relatively high (> 70%) at all sites in this sparsely populated watershed, A. raveneliana distributions exhibit a high degree of both spatial patchiness (i.e., a preference for run-glide mesohabitats with stable substrates and low concentrations of fine sediments) and temporal variability. This is likely indicative of this species’ extreme sensitivity to subtle differences in habitat quality. Changes to instream habitat resulting from land-use change at the 100-m riparian scale appear to have contributed to the local extirpation and fragmentation of populations across the Nolichucky Drainage. These results have potentially profound implications for managing and conserving populations of A. raveneliana and other headwater specialists in this region because they suggest that even streams with heavily forested and well-protected watersheds are not immune to the impacts of seemingly inconsequential land-use change.
In western North Carolina, as in much of the southern Appalachians, headwater streams are intimately linked to associated upland and riparian ecosystems. These watersheds and biotic communities rely on terrestrial habitats for key resources including organic matter and nutrients as well as their role in creating stream geomorphic features (Vannote et al., 1980; Kreutzweiser & Capell, 2001; Pugh et al., 2016, 2020). Land-use modification may potentially affect a broad suite of stream habitat or water quality parameters. Deforestation may lead to increased sediment inputs to streams and increased intensity and frequency of high-flow events (Naiman & Decamps, 1997; Harding et al., 1999; Gulis & Suberkropp, 2003; Strayer & Smith, 2003; Arthington et al., 2009). Sand and silt can occlude interstitial spaces and decrease juvenile mussel survivorship and impair particle assimilation abilities of these filter-feeders (Henley et al., 2000). Siltation of streams has long been linked with mussel declines in the southeastern United States (Kunz, 1898; Ellis, 1936; Brim-Box & Mossa, 1999; Hegeman et al., 2014; Simeone et al., 2021).
Alasmidonta raveneliana distributions provide a salient example of the links between land-use and sedimentation. Populations in the Nolichucky Drainage appear to be very sensitive to changes in sediment composition at the local scale. For example, although we found A. raveneliana in all 4 study streams, occupancy rates at nearly all sites were low and most populations outside the South Toe watershed appeared to be very small. Alasmidonta raveneliana were also not detected at numerous historical and recently occupied sites in this study (Fraley & Simmons, 2006). This is likely due to its sensitivity to environmental changes associated with riparian disturbance including sedimentation as well as changes to conductivity and potentially temperature. Although this species’ life history traits (e.g., fast-growing, short-lived, and early to reproduce) appear to be adaptive for the uncertainties of living in headwaters (sensu Haag & Warren, 1998), the cumulative impacts of land-use change and habitat alteration may have fragmented populations to the extent that re-colonization may no longer be possible in this watershed. It appears that long-term substrate and habitat stability are needed for A. raveneliana populations to establish large, reproducing populations.
Although prior surveys found large numbers of smaller individuals at numerous sites across the drainage, we detected relatively few individuals < 40 mm during 2015 surveys, seemingly suggesting limited recent recruitment. However, analyses of shell thin-sections indicated that A. raveneliana are relatively short-lived and surprisingly (given the cool water temperatures and low dissolved ion concentrations characteristic of the South Toe River) fast-growing mussels. Moreover, examination of shell thin-sections indicates that numerous year classes are present in the South Toe River, suggesting that populations in this stream are stable (Fig. 7). The relatively low number of smaller mussels (< 40 mm) detected may reflect a bias associated with visual-tactile searches or the relatively rapid growth rates exhibited by A. raveneliana (Fig. 2). Similar attributes appear to be widespread among other headwater-adapted mussel taxa (e.g., Alasmidonta, Anodontoides, Strophitus, Villosa spp., Haag & Warren, 1998; Haag, 2012).
The vast majority (95%) of A. raveneliana detected during this study were found in one stream, the South Toe River. Interestingly, surveys failed to detect A. raveneliana populations in this stream prior to 1998 suggesting recent colonization of the South Toe River (USFWS, 2017). Furthermore, A. raveneliana populations in the South Toe River appear to have shifted their range upstream during the last two decades (USFWS, 2017). Protection of the South Toe Watershed may be critical to the survival of Nolichucky Drainage A. raveneliana populations as few other viable populations remain in the drainage. Based on 2011 land cover data, the South Toe River drains the least-impacted watershed in the Nolichucky Drainage. This is likely why it remains a key stronghold for populations of A. raveneliana and numerous other sensitive species present in this system including Brook char [Salvelinus fontinalis (Mitchill, 1814)], Eastern hellbender [Cryptobranchus alleganiensis (Daudin, 1803)], and Blotch-sided logperch (Percina burtoni Fowler, 1945), (NCWRC, 2005).
In addition to the effects of chronic stressors including land-use change and sedimentation, more discrete impacts may have profound implications for the persistence of A. raveneliana populations. Other observations indicate that A. raveneliana populations fluctuate dramatically in abundance and age class structure from year to year and that catastrophic flooding and bedload movements as well as more chronic disturbances appear to play a role in these changes (Fraley & Simmons, 2006; Thompson, 2020; Gangloff unpublished data). Substantial recent changes in A. raveneliana occurrence and abundance may explain why we found relatively strong associations between forest cover and A. raveneliana occurrence despite having sampled a relatively small (n = 25) number of sites. The majority of A. raveneliana were found in the South Toe River and substantial changes to mussel abundance were observed between 2014 and 2015 at several sites in this stream. We suspect that this may be due to increased inputs of fine sediments and habitat changes resulting from an ongoing large-scale highway construction project. We observed a > 50% decline in A. raveneliana abundance and a 63% decline in CPUE from 2014 (18 mussels, 6 mussels per hour) to 2015 (9 mussels, 2.3 mussels per hour) at two sites in the South Toe River. These anecdotal observations are from two survey sites approximately 300 m and 2700 m downstream of the Highway 19 E construction project, respectively. This observation provides support for the hypothesis that fine sediments negatively influence A. raveneliana abundance and suggests that changes may be rapid, underscoring the need to strengthen protection and restoration of riparian buffer zones in streams with A. raveneliana populations.
Management implications
In the Nolichucky Drainage, A. raveneliana populations appear to be stable and reproducing only in the South Toe River. Because populations in other Nolichucky Drainage streams appear to be very small or declining, the South Toe is now likely the major population stronghold in this drainage. Further it may be one of only 4 currently stable A. raveneliana populations remaining globally (2 others also occur in French Broad River tributaries). Continued monitoring of this population (including quantitative surveys to detect recruits and obtain more complete demographic data) is needed to track population trends and ensure that current management strategies do not need to be augmented. Additionally, float surveys of more remote reaches of the mainstem Toe and North Toe Rivers are needed to help determine whether habitats suitable for A. raveneliana exist in other sections of the drainage. The headwaters of the Cane and South Toe rivers may presently be too cool to support large A. raveneliana populations but habitat appears excellent and these streams may provide opportunities for colonization in the future. However, steep channel gradients and a lack of suitable substrates may limit the upstream extent of mussel populations in these streams.
Results of this and other studies suggest that, although mechanistic links between the changes to land-use at the watershed and/or riparian scales and mussel populations may not always be directly apparent. Indirect changes including increased levels of fine sediment and associated dissolved ion inputs and potential changes to stream thermal regimes may have profound consequences for populations of sensitive headwater mussels (Pandolfi, 2016). Ongoing research is examining how juvenile growth and survival vary among A. raveneliana populations in the South Toe River in order to provide insights into how habitat conditions influence mussel populations in this system.
Data availability
Requests for underlying data should be directed to the corresponding author and will be granted under resonable request.
References
Ahearn, D. S., R. W. Sheibley, R. A. Dahlgren, M. Anderson, J. Johnson & K. W. Tate, 2005. Land use and land cover influence on water quality in the last free-flowing river draining the western Sierra Nevada, California. Journal of Hydrology 313: 234–247.
Aldridge, W. W., B. S. Payne & A. C. Miller, 1987. The effects of intermittent exposure to suspended solids and turbulence on three species of freshwater mussels. Environmental Pollution 45: 17–28.
Allan, J. D., 2004. Landscapes and riverscapes: the influence of land use on stream ecosystems. Annual Review of Ecology, Evolution, and Systematics 35: 257–284.
Anthony, J. L., D. H. Kessler, W. L. Downing & J. A. Downing, 2001. Length-specific growth rates in freshwater mussels (Bivalvia: Unionidae): extreme longevity or generalized growth cessation? Freshwater Biology 46: 1349–1359.
Arthington, A. H., R. J. Naiman, M. E. McCain & C. Nilsson, 2009. Preserving the biodiversity and ecological services of rivers: new challenges and research opportunities. Freshwater Biology 55: 1–16.
Benz, G. W. & D. E. Collins (eds), 1997. Aquatic Fauna in Peril; the Southeastern Perspective. Southeast Aquatic Research Institute Special Publication 1. Lenz Design and Communications, Decatur, Georgia.
Brim-Box, J. & J. Mossa, 1999. Sediment, land use and freshwater mussels: prospects and problems. Journal of the North American Benthological Society 18: 99–117.
Brown, K. M. & P. D. Banks, 2001. The conservation of unionid mussels in Louisiana Rivers: diversity, assemblage composition and substrate use. Aquatic Conservation- Marine and Freshwater Ecosystems 11: 189–198.
Clarke, A. H., 1981. The Tribe Alasmidontini (Unionidae: Anodontinae), Part I: Pegias, Alasmidonta, and Arcidens. Smithsonian Contributions to Zoology 399: 1–75.
Cummins, R. H., 1994. Taphonomic processes in modern freshwater molluscan death assemblages: implications for the freshwater fossil record. Paleogeography, Paleoclimatology, Paleoecology 108: 55–73.
Di Maio, J. & L. D. Corkum, 1995. Relationship between the spatial distribution of freshwater mussels (Bivalvia: Unionidae) and the hydrological variability of rivers. Canadian Journal of Zoology 73: 663–671.
Ellis, M. M., 1936. Erosion silt as a factor in aquatic environments. Ecology 17: 29–42.
Fuller, R. L., C. Griego, J. D. Muehlbauer, J. Dennison & M. W. Doyle, 2010. Response of stream macroinvertebrates in flow refugia and high-scour areas to a series of floods: a reciprocal replacement study. Journal of the North American Benthological Society 29: 750–760.
Fraley, S. & J. Simmons, 2006. An assessment of selected rare mussel populations in western North Carolina following extraordinary floods of September 2004. NC Department of Environment and Natural Resources Permit Report, Raleigh, NC.
Gangloff, M. M. & J. W. Feminella, 2007. Stream channel geomorphology influences mussel abundance in southern Appalachian streams, USA. Freshwater Biology 52: 64–74.
Gergel, S. E., M. G. Turner, J. R. Miller, J. M. Melack & E. H. Stanley, 2002. Landscape indicators of human impacts to riverine systems. Aquatic Science 64: 118–128.
Gordon, N. D., T. A. McMahon, B. L. Finlayson, C. J. Gippel & R. J. Nathan, 2004. Stream Hydrology: An Introduction for Ecologists, 2nd ed. Wiley, West Sussex:
Glenn, C. L., 1911. Denudation and Erosion in the Southern Appalachian Region and the Monongahela Basin. Washington Government Printing Office. United States Geological Survey.
Gulis, V. & K. Suberkropp, 2003. Leaf litter decomposition and microbial activity in nutrient-enriched and unaltered reaches of a headwater stream. Freshwater Biology 48: 123–134.
Haag, W. R., 2012. North American Freshwater Mussels: Natural History, Ecology and Conservation, Cambridge University Press, Cambridge:
Haag, W. R. & A. M. Commens-Carson, 2008. Testing the assumption of annual shell ring deposition in freshwater mussels. Canadian Journal of Fisheries and Aquatic Sciences 508: 493–508.
Haag, W. & M. Warren, 1998. Role of ecological factors and reproductive strategies in structuring freshwater mussel communities. Canadian Journal of Fisheries and Aquatic Systems 52: 78–91.
Harding, J. S., E. F. Benfield, P. V. Bolstad, G. S. Helfman & E. B. D. Jones, 1998. Stream biodiversity: the ghost of land use past. Proceedings of the National Academy of Sciences 95: 14843–14847.
Harding, J. S., R. G. Young, J. W. Hayes, K. A. Shearer & J. D. Stark, 1999. Changes in agricultural intensity and river health along a river continuum. Freshwater Biology 42: 345–357.
Hastie, L., 2011. Are Scottish freshwater pearl mussel populations recruiting normally? Toxicology and Environmental Chemistry 93: 1748–1763.
Hegeman, E. E., S. W. Miller & K. E. Mock, 2014. Modeling freshwater mussel distribution in relation to biotic and abiotic habitat variables at multiple spatial scales. Canadian Journal of Fisheries and Aquatic Sciences 71: 1483–1497.
Henley, W. F., M. A. Patterson, R. J. Neves & D. A. Lemly, 2000. Effects of sedimentation and turbidity on lotic food webs: a concise review for natural resource managers. Reviews in Fisheries Science 8: 125–139.
Hopkins, R. L., 2009. Use of landscape pattern metrics and multiscale data in aquatic species distribution models: a case study of a freshwater mussel. Landscape Ecology 24: 943–955.
Hopkins, R. L. & J. C. Roush, 2013. Effects of mountaintop mining on fish distributions in central Appalachia. Ecology of Freshwater Fish 22: 578–586.
Jarvis, J. D., 2011. Water quality in the upper Little Tennessee River and its potential effects on the Appalachian elktoe mussel (Alasmidonta reveneliana). Master’s Thesis. Western Carolina University. https://libres.uncg.edu/ir/wcu/clist.aspx?id=4604.
Kat, P. W., 1982. Effects of population density and substratum type on growth and migration of Elliptio complanata (Bivalvia: Unionidae). Malacological Review 15: 119–127.
King, R. S., M. E. Baker, D. F. Whigham, D. E. Weller, T. E. Jordan, P. F. Kazyak & M. K. Hurd, 2005. Spatial considerations for linking watershed land cover to ecological indicators in streams. Ecological Applications 15: 137–137.
Kreutzweiser, D. & S. Capell, 2001. Fine sediment deposition in streams after selective harvesting without riparian buffers. Canadian Journal of Forest Research 31: 2134–2142.
Kunz, G. F., 1898. A brief history of the gathering of fresh-water pearls in the United States. Bulletin of the United States Fish Commission 17: 321–330.
Lancaster, J. & A. Hilldrew, 1993. Flow refugia and the microdistribution of lotic macroinvertebrates. Canadian Journal of Fisheries and Aquatic Sciences 12: 385–393.
Levine, J., S. Salger, L. Borst, M. Law, C. Eads, C. Osburn, M. M. Gangloff, S. Sumner & B. Sauls, 2015. Investigation of Factors Contributing to the Decline in the Nutritional Health of Alasmidonta raveneliana in the Little Tennessee River, Franklin, North Carolina. NCDOT Project Report 2013-2012-37. FHWA/NC/2009-08. Raleigh, NC.
Lydeard, C. & R. L. Mayden, 1995. A diverse and endangered aquatic ecosystem of the Southeast United States. Conservation Biology 9: 800–805.
Maloney, K. O., J. W. Feminella, R. M. Mitchell, S. A. Miller, P. J. Mulholland & J. N. Hosuer, 2008. Land-use legacies and small streams: identifying relationships between historical land use and contemporary stream conditions. Journal of the North American Benthological Society 27: 280–294.
Merwade, V., 2012. Watershed and stream network delineation using ArcHydro tools. School of Civil Engineering, Perdue University, Perdue, IN 1: 1–22.
Naiman, R. J. & H. Decamps, 1997. The ecology of interfaces: riparian zones. Annual Review of Ecology and Systematics 28: 621–658.
Neves, R. J. & S. N. Moyer, 1988. Evaluation of techniques for age determination of freshwater mussels (Unionidae). American Malacological Bulletin 6: 179–188.
North Carolina Wildlife Resources Commission, 2005. North Carolina Wildlife Action Plan, Raleigh, NC:
Ogle, D. H., 2016. FSA: Fisheries stock analysis. R package 0.8.6.
Pan, Y., A. Herlihy, P. Kaufmann, J. Wigington, J. van Sickle & T. Moser, 2004. Linkages among landuse, water quality, physical habitat conditions and lotic diatom assemblages: a multi-spatial scale assessment. Hydrobiologia 515: 59–73.
Pandolfi, G. P. 2016. Effects of climate, land use and in-stream habitat on Appalachian elktoe (Alasmidonta raveneliana) in the Nolichucky River drainage, North Carolina. Master’s Thesis. Appalachian State University. https://libres.uncg.edu/ir/asu//listing.aspx?id=21052.
Pandolfo, T. J., W. G. Cope, C. Arellano, R. B. Bringolf, M. C. Barnhart & E. Hammer, 2010. Upper thermal tolerances of early life stages of freshwater mussels. Journal of the North American Benthological Society 29: 959–969.
Parmalee, P. W. & A. E. Bogan, 1998. The Freshwater Mussels of Tennessee, 1st ed. The University of Tennessee Press, Knoxville:
Paul, M. & J. Meyer, 2001. Streams in the urban landscape. Annual Review of Ecology and Systematics 33: 333–365.
Pugh, M. W., M. Hutchins, M. Madritch, L. Siefferman & M. M. Gangloff, 2016. Land-use and local physical and chemical habitat parameters predict site occupancy by hellbender salamanders. Hydrobiologia 1: 105–116.
Pugh, M. W., G. P. Pandolfi, T. F. Franklin & M. M. Gangloff, 2020. Influences of in-stream habitat and upstream land-use on site occupancy of the Kanawha darter (Etheostoma kanawhae): a narrowly distributed species from the New River (Upper Kanawha Basin). Aquatic Conservation: Marine and Freshwater Ecosystems 31: 494–503.
R Development Core Team, 2008. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna.
Scott, M. C., 2006. Winners and losers among stream fishes in relation to land use legacies and urban development in the southeastern US. Biological Conservation 127: 301–309.
Shea, C. P., J. T. Peterson, M. J. Conroy & J. M. Wisniewski, 2013. Evaluating the influence of land use, drought and reach isolation on the occurrence of freshwater mussel species in the lower Flint River Basin, Georgia (U.S.A.). Freshwater Biology 58: 382–395.
Simeone, D., C. H. Tagliaro & C. R. Beasley, 2021. Novel insights into habitat suitability for Amazonian freshwater mussels linked with hydraulic and landscape drivers. Ecology and Evolution 11: 11786–11798.
Singer, E. E. & M. M. Gangloff, 2011. Effects of a small dam on freshwater mussel growth in an Alabama (U.S.A.) stream. Freshwater Biology 56: 1904–1915.
Snyder, C. D., J. A. Young, R. Villella & D. P. Lemarié, 2003. Influences of upland and riparian land use patterns on stream biotic integrity. Landscape Ecology 18: 647–664.
Strayer, D. L., 1999. Use of flow refuges by unionid mussels in rivers. Journal of the North American Benthological Society 18: 468–476.
Strayer, D. L. & J. Ralley, 1993. Microhabitat use by an assemblage of stream-dwelling unioaceans (Bivalvia), including two rare species of Alasmidonta. Journal of the North American Benthological Society 12: 247–258.
Strayer, D. L. & D. R. Smith, 2003. A Guide to Sampling Freshwater Mussel Populations, Vol. 8. American Fisheries Society, Monograph, Washington:
Thompson, M., 2020. Influence of Interstitial Sediments on an Endangered Freshwater Mussel Population. Master’s Thesis. Appalachian State University. http://libres.uncg.edu/ir/asu/f/Thompson_Michael_May%202020_Thesis.pdf.
United State Environmental Protection Agency, 2018. Water Quality Data (WQX) web portal. https://www.epa.gov.waterdata/water-quality-data-wqx#portal.
United States Fish and Wildlife Service, 1994. Endangered and threatened wildlife and plants, Appalachian elktoe determined to be an endangered species. Federal Register 59: 60324–60334.
United States Fish and Wildlife Service, 2002. Endangered and Threatened Wildlife and Plants; Designation of Critical Habitat for the Appalachian Elktoe, Final Rule. Federal Register 66: 61016–61040.
United States Fish and Wildlife Service, 2017. Appalachian elktoe 5-Year Review. US Fish and Wildlife Ecological Services Field Office, Asheville.
Vannote, R. L. & G. W. Minshall, 1982. Fluvial processes and local lithology controlling abundance, structure, and composition of mussel beds. Proceedings of the National Academy of Sciences 79: 4103–4107.
Vannote, R. L., G. W. Minshall, K. W. Cummins, J. R. Sedell & C. E. Cushing, 1980. The river continuum concept. Canadian Journal of Fisheries and Aquatic Sciences 37: 130–137.
von Bertalanffy, L., 1938. A quantitative theory of organic growth (Inquiries on growth laws II). Human Biology 10: 181–213.
Weijters, M. J., J. H. Janse, R. Alkemade & J. T. A. Verhhoeven, 2009. Quantifying the effect of catchment land use and water nutrient concentrations on freshwater river and stream biodiversity. Aquatic Conservation: Marine and Freshwater Ecosystems 19: 104–112.
Williams, J. D., A. E. Bogan, R. S. Butler, K. S. Cummings, J. T. Garner, J. L. Harris, N. L. Johnson & G. T. Watters, 2017. A revised list of the freshwater mussels (Mollusca: Bivalvia: Unionoida) of the United States and Canada. Freshwater Mollusk Biology and Conservation 20: 33–58.
Wolman, M., 1954. A Method to Sampling Course River-Bed Material. Transactions of the American Geophysical Union 35: 6.
Acknowledgements
Funding for this research was provided by Grants from the U.S. Fish and Wildlife Service to MG and from the Appalachian State University Office of Student Research and the Graduate Student Association to GP. We thank T. Franklin, S. Geda, D. Mason, W. Pugh, J. Selong, M. Thompson, M. Winebarger (ASU); J. Fridell (USFWS, ret) and A. Goetz (USFWS, ret); S. Fraley and W.T. Russ (NCWRC) for assistance with field sampling and access to historical data. Mark Endries and Doug Newcomb (USFWS) assisted with land-use and statistical analyses. Robert Butler (USFWS, ret), Michael Madritch (ASU), and members of the Gangloff and Siefferman labs at Appalachian State University read earlier versions of this publication and provided helpful comments on the draft.
Disclaimer
The findings and conclusions in this article are those of the author(s) and do not necessarily represent the views of the U.S. Fish and Wildlife Service.
Funding
Funding was provided by grants from the U.S. Fish and Wildlife Service to Dr. Michael Gangloff and from the Appalachian State University Office of Student Research and the Graduate Student Association to Mr. Gary Pandolfi. The datasets generated during and/or analyzed during the current study are available from the corresponding author upon reasonable request. The R Code and publicly obtained datasets utilized in this research are publicly available via the included references.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
There are no conflicting or competing interests among the authors or funding sources.
Ethical approval
This manuscript adheres to the Committee on Publication Ethics (COPE) and follows their guidelines. Human study subjects were not included in this research. Invertebrates (e.g., mussels) are exempt from Institutional Animal Care and Use Committee regulations. However, live animals encountered in this study were handled with care and promptly released unharmed into the wild. Appropriate federal, state, and local permits as well as private landowner access was obtained and adhered to. All authors freely consent to having participated in this research project, data analysis, and manuscript drafting. All authors consent to have this manuscript published as an original research article.
Additional information
Handling editor: Manuel Lopes-Lima
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Pandolfi, G.S., Mays, J.W. & Gangloff, M.M. Riparian land-use and in-stream habitat predict the distribution of a critically endangered freshwater mussel. Hydrobiologia 849, 1763–1776 (2022). https://doi.org/10.1007/s10750-022-04826-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-022-04826-8