
Abstract
We evaluated the association between the El Niño Southern Oscillation (ENSO), an important climatic phenomenon on Earth, with water level variation and fish assemblage attributes (i.e., abundance, species richness and structure of the assemblage) of the Paraná (dammed) and Ivinhema (undammed) rivers, located in the upper Paraná River floodplain, Brazil. Cross-Correlation Function analyses were performed using the Oceanic Niño Index (ONI; which indexed ENSO events) as the influential time series on water level and fish assemblage attributes time series. ONI were positively associated with water level and with most of the fish assemblage attributes tested, but these patterns differed between the two rivers. This indicates that El Niño events affect the structure of the fish assemblages, mainly in the Ivinhema River, which presented more assemblage attributes associated with ONI. In addition, dams may play a regulatory effect over the water level of the Paraná River, which probably explains the different patterns of assemblage attributes in both rivers. Thus, ENSO events are associated with the composition and structure of the fish assemblages in the studied area, and their effects are apparently minimized in the Paraná River due to the presence of upstream dams.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
El Niño Southern Oscillation (ENSO) is a large-scale climatic phenomenon related to variations in the sea surface temperature of the Pacific Ocean (Philander, 1983; Trenberth, 1997), which can lead to several meteorological changes, and it is classified in warm (El Niño) and cold (La Niña) phases. The strength of ENSO events can be measured with the Oceanic Niño Index (ONI), which allows inferences on the occurrence and intensity of El Niño or La Niña events. These events may result in quite different responses across regions and impact both marine and continental ecosystems (Caviedes, 2001; Garcia et al., 2004, Collins et al., 2010; Su et al., 2018; Pereira et al., 2020). They can cause changes in the global hydrological cycle, as well as patterns of precipitation resulting in extreme drought or flood in different parts of the planet (Ropelewski & Halpert, 1987; Philander, 1990; Grimm & Tedeschi, 2008; Henderson et al., 2018).
Environmental changes caused by ENSO events (i.e., El Niño and La Niña) may affect diverse parameters of ecological communities in different temporal and spatial scales (Holmgren et al., 2001; Glynn et al., 2017; Pineda et al., 2019). Regarding fish assemblages, changes in the composition and in patterns of distribution, recruitment, migration, and seasonality of several species have been directly related to these events (Godínez-Domínguez et al., 2000; Garcia et al., 2001; Garcia et al., 2004; Brander, 2007). These effects are well demonstrated for marine environments (Hollowed et al., 2001; Watters et al., 2003; Lehodey et al., 2005; Booth et al., 2011) and estuaries (Mol et al., 2000; Garcia et al., 2003; Possamai et al., 2018), where environmental changes can lead to a delayed response in the recruitment of some species (Moraes et al., 2012). However, few studies explored ENSO effects over freshwater fishes. In this context, historical drought events related to ENSO were associated with high mortality of fishes due to algal blooms in the Fly River system (Swales et al., 1999), as well as with the collapse of a migratory species in the Pilcomayo River floodplain, which depends on high water levels for their reproductive migration process (Smolders et al., 2000).
The hydrological cycle of diverse basins around the world are related to ENSO events, which may influence the frequency of floods, drought, water quality, discharge, and river flow (Chiew et al., 1998; Keener et al., 2010; Su et al., 2018). However, the natural conditions of many rivers – and consequently their biota – have been intensely modified by human activities, especially by the construction of dams (Dudgeon et al., 2006; Agostinho et al., 2008). For example, dams can reduce floods that would be caused by ENSO events, and may intensify drought events downstream (Ponton, 2001). Dams change rivers from lotic to lentic systems, affecting physical and chemical properties of water, nutrient cycling, habitat structure, species distribution and taxonomic and functional composition of communities (Tundisi & Straskraba, 1999; Fernandes et al., 2009; Oliveira et al., 2015). Fish diversity is intensely affected by dams, since these structures may affect the abundance, reproduction, development, recruitment of several species, and they often cause local or even regional extinctions (Terra et al., 2010; Cheng et al., 2015; Moran et al., 2018). These impacts are even more relevant considering migratory species, which depend on floods to fulfill their life cycles (Gubiani et al., 2007; Agostinho et al., 2007, 2016). Migratory species travel over long distances, depend on floods for reproduction, and they also use adjacent habitats to the main channel of rivers (e.g., floodplain lakes or oxbow lakes) as nursery and feeding sites (Baumgartner et al., 2004; Agostinho et al., 2004a, 2008).
In different stretches of the Paraná River (in Brazil, Paraguay and Argentina), it was demonstrated that the interannual variations in flow and water discharge are associated with ENSO events (Berri et al., 2002; Dai et al., 2009; Antico et al., 2018). El Niño events were also linked to intense floods in these regions in recent decades (Camilloni & Barros, 2000; Fernandes et al., 2009). However, in Brazil, there are about 150 reservoirs in the Paraná River and in its main tributaries, which have strong regulatory effects over their hydrological regime, changing historical patterns and, consequently, disturbing aquatic communities (Ward & Stanford, 1995; Agostinho et al., 2007; Stevaux et al., 2009; Agostinho et al., 2016). The Upper Paraná River region holds an extensive floodplain, composed of the Paraná River, its main channel, and regulated and unregulated tributaries (e.g., Paranapanema and Ivinhema rivers, respectively). The floodplain was intensively affected by the construction of the Sérgio Motta dam in its main channel, which halved the floodplain extension area, strongly affecting the water discharge and the water level oscillations (Agostinho et al., 2004a; Gubiani et al., 2007; Stevaux et al., 2009).
Paraná and Ivinhema rivers were recorded to have significant differences considering their fish assemblages, and one of the main factors influencing these differences is the respectively presence and absence of upstream dams (Fernandes et al., 2009; Granzotti et al., 2018; Oliveira et al., 2018). Since dam constructions have important regulatory effects on the water flow in downstream sites, it is possible that these impoundments interfere in the ENSO influence in the Paraná River. Based on this, the main goal of this paper was to evaluate the effects of ENSO (indexed by ONI) in the fish assemblages of the upper Paraná River floodplain. For this, we seek to answer the following questions: (i) Are ENSO events directly associated with water level of the rivers that compose the upper Paraná floodplain?; (ii) Do ENSO events influence fish assemblage attributes (total abundance, abundance of migratory species, species richness and assemblage structure) of Paraná and Ivinhema rivers?; and (iii) Are there different association patterns regarding ENSO events with water level and assemblage attributes in the dammed and undammed rivers?
Material and Methods
Study area
The upper Paraná River floodplain is located in the last lotic stretch of the Paraná River inside Brazil, between the Itaipu Reservoir (downstream) and the Sérgio Motta Dam (locally known as Porto Primavera; upstream). This stretch is approximately 250 km in length and covers an area of 5,268 km2 (Agostinho et al., 2004b). The main tributary in the East margin is the Paranapanema River, which is also intensely dammed (the last dam in this river is the Rosana Reservoir) and contributes to the regulation of the Paraná River water level.
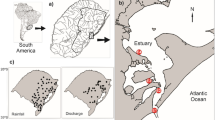
The floodplain consists of three rivers: Ivinhema, Paraná, and Baía. For this study, sampling was carried out in six stations: three in the Paraná River (intensely impacted by the construction of dams) and three in the Ivinhema River (free of dams and in more pristine conditions), including the main channel and adjacent floodplain lakes, connected or not connected to the main channel of the rivers (Fig. 1). Sampling stations are sentinel stations of a Long Term Ecological Research (LTER), Site 6: Ventura Lake, Patos Lake, Ivinhema River channel (all located in the Ivinhema River), and Pau Véio Lake, Garças Lake and Paraná River channel (all located in the Paraná River).

Study area of the upper Paraná River floodplain, where: 1—Ventura Lake; 2—Patos Lake; 3—Ivinhema River; 4—Pau Véio Lake; 5—Paraná River; 6—Garças Lake
Data collection
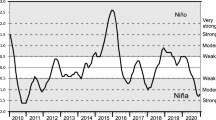
Monthly ENSO data from 1964 to 2018 were obtained from the National Oceanic and Atmospheric Administration (NOAA). ENSO data are given by the Oceanic Niño Index (ONI), for which values greater than or equal to 0.5 represent El Niño events and values less than or equal to -0.5 represent La Niña events (Fig. 2a). The values of ONI are calculated as quarterly means of ERSST.v5 SST anomaly for the Niño 3.4 region (i.e., 5° N–5° S, 120°–170° W; Huang et al., 2017). Values between − 0.5 and 0.5 represent Neutral periods when none of the events had strength to occur (i.e., absence of ENSO events).

Temporal series of the Oceanic Niño Index (ONI) and of the water level used. a Annual variation of ONI values (1964–2018); Dashed lines = thresholds of El Niño and La Niña events. b Daily variation in the water level of the Paraná River (1964–2018), which RD Rosana Dam; SMD Sergio Motta Dam; Dashed lines = period of 2000–2016 in the Paraná River. c Daily variation in the water level of the Ivinhema River (2000–2016)
The hydrological variable used was the water level, from which we obtained two temporal series: 1964–2018 for the Paraná River and 2000–2016 for the Ivinhema River. The daily water level (m) of the Paraná River was obtained from Porto São José fluviometric station (code 64575000 – “Agência Nacional de Águas”; National Water Agency), located about 12 km upstream the study area. The data of Paraná River were grouped and analyzed in four time periods (Fig. 2b): (i) until the construction of Rosana Dam in the Paranapanema River (1964–1984); (ii) until the construction of Sérgio Motta Dam in the Paraná River (1964–1997); (iii) the complete temporal series (1964–2018), to verify the degree of alterations prompted by dams in the water level of the Paraná River; and (iv) the analogous temporal series for the Ivinhema River (2000–2016), to compare the two rivers in the same time scale. Daily water levels (m) of the Ivinhema River were obtained from Porto Sumeca fluviometric station (code 64617000 – “Agência Nacional de Águas”), located about 50 km upstream the study area (Fig. 2c).
Fish samplings were carried out quarterly (as the values of the ONI) in the surveys conducted in the Paraná and Ivinhema rivers under the Long Term Ecological Research, from 2000 and 2018. Fishes were collected using 11 gillnets (20 m long) with different mesh sizes (2.4 until 16.0 cm between opposite knots), which were operated near the margins and set for 24 h in each sampling location. Sampled individuals were anesthetized with eugenol (Resolution n° 1000/12, “Conselho Federal de Medicina Veterinaria”), euthanized, packed in thermal boxes, and transported to the field laboratory. After, fish were identified (Graça & Pavanelli, 2007; Ota et al., 2018). For each individual captured, we recorded the date and sampling station. As the fishing effort were the same for all samplings, the number of individuals caught was considered the abundance. Fish data were evaluated separately according to each river (Paraná and Ivinhema rivers). As we were not interested in the spatial variability inside each river, sampling locations (lakes and main channel) were not considered in the analysis.
Data analysis
To evaluate if ENSO events are directly associated with water level (first question), we applied cross-correlation function (CCF) analyses (Chatfeild & Xing, 2019) considering ONI as the time series of influence and the water level time series as the affected time-series (see Probst et al., 2012). For the water level data, CCFs were performed with each period of Paraná and Ivinhema rivers described in the section data collection, to evaluate the possible influence of dams on the floodplain responses to ENSO events.
To determine if ENSO events influence fish assemblage attributes (second question), we calculated the total abundance, abundance of migratory species, and species richness for each monthly sample. To assess the variation in the structure of fish assemblages for each river, a Principal Coordinate Analysis was applied (PCoA; Anderson et al., 2006) on the abundance data using the function “cmdscale” from the package “stats” in the R Environment (R Core Team, 2020). The PCoA was performed on the Bray-Curtis resemblance matrix and the first two axes of PCoA, which represented most of the variability in the original data matrix, were retained for interpretation. Thus, we obtained five dependent variables for each river (total abundance, abundance of migratory species, species richness, and structure of fish assemblage represented by PCoA1 and PCoA2), referred as fish assemblage attributes. After, cross-correlation functions were performed for each river separately, with ONI as the time series of influence and each assemblage attribute as the affected time series.
Cross-correlation function analysis allowed us to quantify the association and to identify the lags between the two-time series as a function of the displacement of one of the series relative to the other (third question). Thus, we were able to evaluate the nature of the association (positive or negative) of ONI with water level and assemblage attributes, the lag which presented the higher correlation value in these relationships (i.e., the time in which the association was stronger), and the extent of the response of the affected temporal series (i.e., how long the correlation was significant).
As the hydrological cycle lasts approximately a year in the upper Paraná River floodplain, CFFs were performed with a maximum lag of one year for water level. On the other hand, CFFs were performed with a maximum lag of two years for fish assemblage attributes since some fish species might present a delayed response to environmental changes (Moraes et al., 2012; Baumgartner et al., 2018). Juveniles of migratory species, for example, will be recruited to fishery stocks about one or two years after flood events (Agostinho et al., 1993; Gomes & Agostinho, 1997; Agostinho et al., 2004a; Oliveira et al., 2015). According to our sampling design, each lag in CCFs output for water level corresponded to one day, while for fish assemblage attributes time series corresponded to one month.
In some cases, it might be difficult to interpret which variable is leading and which variable is lagging in temporal analyses (Lingard et al., 2017). Here, we assume that ONI leads water level and fish assemblage attributes since the opposite was ecologically impossible. Therefore, only the left side of the CCFs graphical results was interpreted, which despite presenting negative values on the horizontal axis, it represents positive time lags of water level and assemblage attributes relative to the ONI time series. As it was difficult to visualize the stronger correlations in the graphical results of the CCFs regarding to water level data, we applied a LOESS Curve Fitting (Local Polynomial Regression) to find the higher lag-correlation between ONI and water level of each river. Therefore, water level results of CFFs were also summarized in a table. Significant values of CCFs were obtained by \(\left( {\alpha = 0.05} \right)\) at \(\pm 2/\surd n\), where n is the length of the time series (Berryman & Turchin, 2001; Chatfeild & Xing, 2019). The analyses were performed using the functions “ccf” from the package “forecast” (Hyndman et al., 2020), “cmdscale” and “loess” from the package “stats” in R Environment (R Core Team, 2020).
Results
The four analyzed periods of the water level in the Paraná River showed a declining pattern over time regarding the association with ONI, and an increasing pattern regarding time lag, after the construction of each dam (Rosana Dam and Sergio Mota Dam; Table 1). A higher correlation value between ONI and Paraná River time series was found before the construction of the Rosana Dam. The period before the construction of the Sérgio Mota Dam presented a similar correlation value with the entire time series of the Paraná River. The CCF between ONI and water level of the Ivinhema River presented the highest correlation value when compared with all Paraná River temporal series, mainly considering the same period (2000–2016; Table 1). In addition, the time lag of the Ivinhema River demonstrated a similar trend to the period of 1964–1984 in the Paraná River. In general, the extent of the water level time series regarding ONI lasts the entire period (maximum lag of one year) for both rivers (Fig. 3).

Cross-Correlation Functions (CCFs) between the Oceanic Niño Index (ONI) and the water level of Paraná and Ivinhema rivers. a period of 1964–1984 of Paraná River; b period of 1964–1997 of Paraná River; c the entire time-series of Paraná River (1964–2018); d period of 2000–2016 of Paraná River; e Ivinhema River time-series (2000–2016). The dashed lines indicate the thresholds of significance
The CCFs performed with the fish assemblage attributes showed different patterns between Paraná and Ivinhema rivers (Fig. 4). The Paraná River presented a significant and negative association between the total abundance and ONI (r = -0.193; Fig. 4a), while a significant and positive associations were found for the abundance of migratory species (r = 0.251; Fig. 4c) and PCoA 2 (r = 0.308; Fig. 4i) with ONI. Species richness and PCoA 1 did not present significant correlation values with ONI in the Paraná River (Fig.4e and 4g, respectively). Besides, for the Paraná River, the variables which presented the higher extent were total abundance and PCoA 2 (16 months) while the variable which presented the lower extent was abundance of migratory species (11 months). Considering the time with higher correlation values, Paraná River presented higher time lags correlation in 14 months for total abundance, 8 months for the abundance of migratory species, and 12 months for PCoA2.

Cross-Correlation Functions (CCFs) between the Oceanic Niño Index (ONI) and each fish assemblage attribute of Paraná (left side graphs) and Ivinhema (right side graphs) river. The dashed lines indicate the thresholds of significance. E = extent of the association (considering the last significant value of lag)
In the Ivinhema River, all fish assemblage attributes presented significant and positive associations with ONI (Correlation values: Total abundance = 0.279, Abundance of migratory species = 0.216, Species Richness = 0.256, PCoA1 = 0.219; Fig. 4b, d, f, h, respectively), except PCoA 2 (Fig. 4j). In addition, in the Ivinhema River, the variable which presented the higher extent was species richness (16 months) while the variables which presented the lower extent were abundance of migratory species and PCoA 1 (12 months). Considering the time with higher correlation values, Ivinhema River presented higher correlation lags in 8 months for total abundance and for abundance of migratory species, in addition to 6 months for species richness and for fish assemblage structure, represented in PCoA1.
Discussion
Answering our three questions, the results of this study showed that: (i) ENSO events are directly associated with the water level variation of the studied rivers; (ii) ENSO events influenced fish assemblage attributes of Paraná and Ivinhema rivers; and (iii) water level and fish assemblage attributes in the dammed and undammed river presented different association patterns regarding to ENSO events.
There was a positive and significant association between ONI (ENSO index) and water level of Paraná and Ivinhema rivers. However, this association decreased in the Paraná River as the river became more regulated, suggesting that upstream dams may interfere in the influence of ENSO on the water level of the Paraná River. This can be sustained mainly by the contrasting results in the period of 2000–2016 of Paraná and Ivinhema rivers, which represented the period that the water level time-series of the Paraná River was most intensely affected by upstream dams. Moreover, the increasing values of the time lag response of water level to ONI also corroborated our argument that dams may influence the ENSO effects in this river. It is also important to highlight that, in the Paraná River, the water level variation was greatly changed after the construction of dams, decreasing the frequency and intensity of floods, and also decreasing maximum and minimum water levels, as demonstrated by our historical series, and previously by Gubiani et al. (2007). Meanwhile, the Ivinhema River is free of dams and presented a higher correlation value than all periods in the Paraná River.
Higher values of ONI, which represent El Niño events, were associated with higher values of water level and most of the fish assemblage attributes in the Paraná and Ivinhema rivers. Thus, it is possible that El Niño events are associated to floods in the studied rivers as suggested by Fernandes et al. (2009), indicating that these events affect the structure of the fish assemblages and their attributes, mainly in the Ivinhema River due to the absence of dams regulating its water level. Previous studies showed that ENSO events may present a strong effect on fish assemblages that inhabit floodplains, regarding its effects on the hydrological regime (Swalles, 1999; Smolders et al., 2000; Camacho Guerreiro et al., 2020). Besides, Ponton (2001) also evidenced that dams may amplify droughts caused by these events on the Sinnamary River, in the Amazon region, directly affecting the fish assemblage downstream. Thus, our results corroborate that ENSO events may affect fish assemblages in floodplain ecosystems, but the presence of dams upstream can interfere in this association.
The differences found between fish assemblage attributes in the Paraná and Ivinhema rivers are probably related to the different environmental conditions of each river. In the Paraná River, dams cause the retention of sediments and flow regulation which leads to several impacts downstream, mainly considering the fish assemblage (Gubiani et al., 2007; Granzotti et al., 2018). Dams interrupt migration routes of migratory species and reduce the recruitment of fish species by trapping eggs and larvae in upstream sites, besides changing the food-web structure and favoring visual piscivorous fish (Suzuki et al., 2011; Turgeon et al., 2019; Rodrigues et al., 2020). In contrast, the Ivinhema River presents more pristine conditions, with higher values of total abundance and species richness than the Paraná River (Fernandes et al., 2009; Granzotti et al., 2018; Oliveira et al., 2018). Thus, it is possible that the differences in fish abundance and species richness between Paraná and Ivinhema rivers lead to different responses regarding ENSO events, as evidenced by our results.
Total abundance of migratory species was positively associated to ONI in the Paraná and Ivinhema rivers, despite the negative effect of dams over these species in the Paraná River (Agostinho et al., 2004a Makrakis et al., 2019). This result demonstrated that higher water levels triggered by El Niño events may favor the maintenance of migratory species populations, which take advantage of floods in the Paraná River and migrate to unregulated tributaries to spawn (Agostinho et al., 2003; Baumgartner et al., 2004). Besides, regarding the extent of the response to ONI, we found that the abundance of migratory species presented a significant association until 11 months and 12 months in the Paraná and Ivinhema rivers, respectively. Recruitment and abundance responses of migratory species may be slower when compared with non-migratory species due to their long life cycles (Agostinho et al., 1993; Gomes & Agostinho; 1997; Bailly et al., 2008; Suzuki et al., 2009; Oliveira et al., 2015), occurring a year after the flood (Agostinho et al., 2004a). Thus, the extent of the association in the two rivers may be explained by the delayed recruitment of migratory species.
The structure of the fish assemblages (indexed by the PCoA axes) was also positively associated to ONI (PCoA 2 for the Paraná River and PCoA 1 for the Ivinhema River). This was expected since life history traits of fishes of the upper Paraná River floodplain are closely related to the hydrological cycle (Agostinho et al., 2004c). Drought periods can facilitate the predatory activities of piscivorous fish, by increasing the confinement and density of prey (Rodríguez & Lewis, 1997; Turesson & Bronmark, 2007). On the other hand, flood periods can increase the input of allochthonous feeding resources for fish (e.g., detritus), provide nursery areas and shelter by flooding terrestrial vegetation, and generate bottom-up effects by affecting other organisms of the floodplain (e.g., phytoplankton, zooplankton and macrophytes) (Bailly et al., 2008; Aspin et al., 2018; Granzotti et al., 2018; Quirino et al., 2018).
Final considerations
The Oceanic Niño Index (ONI) proved to be a good proxy to assess the relationship of ENSO events with the water level variation and fish assemblages. The relationship between ENSO events and hydrological variables of the Paraná River has already been demonstrated in previous studies (Amarasekera et al., 1997; Camilloni & Barros, 2000; Berri et al. 2002; Antico et al., 2018), but for other regions of the basin. Here, beyond the association between ONI and the water level of the rivers that compose the upper Paraná floodplain, we demonstrated how dams may interfere in the effects of ENSO events in the freshwater environments of the region. Besides, previous studies suggested that ENSO effects were associated with fish assemblage attributes (e.g., Fernandes et al., 2009), and our findings corroborated this association, which might also be true for other tropical floodplains. El Niño events were associated with migratory species in both rivers and this response was evidenced until nearly one year after the events. It has also been shown that the construction of dams seems to have an even greater impact on the environments and organisms that live downstream, once dams can interfere in the effects of major climatic events over the ecosystems. Further studies should consider the temporal dynamics of changes caused by ENSO events in different hydrological attributes (e.g., seasonality, duration and intensity of floods), and consequently these effects over biological communities.
Data availability
The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Agostinho, A. A., L. C. Gomes, H. I. Suzuki & H. F. Júlio-Júnior, 2003. Migratory fish from the upper Paraná River basin, Brazil. In: Carolsfeld, J., B. Harvey, C. Ross, A. Baer & Ross, C. (eds). Migratory Fishes of South America: Biology, Social Importance and Conservation Status. World Fisheries Trust, the World Bank and the International Development Research Centre, Victoria, Canada.
Agostinho, A. A., A. E. A. Vazzoler, L. C. Gomes & E. K. Okada, 1993. Estratificacion espacial y comportamiento de Prochilodus scrofa en distintas fases del ciclo de vida, en la planici de inundacion del alto rio Paraná y embalse de Itaipu, Paraná. Brasil. Rev. Hydrobiol. Trop 26: 79–90.
Agostinho, A. A., L. C. Gomes, S. M. Thomaz & N. S. Hahn, 2004a. The upper Paraná River and its floodplain: main characteristics and perspectives for management and conservation. In Thomaz, S. M., A. A. Agostinho & N. S. Hahn (eds), The upper Paraná River and its floodplain. Backhuys Publishers, Leiden, The Netherlands.
Agostinho, A. A., L. C. Gomes, S. Veríssimo & E. K. Okada, 2004b. Flood regime, dam regulation and fish in the Upper Paraná River: effects on assemblage attributes, reproduction and recruitment. Reviews in Fish biology and Fisheries 14: 11–19.
Agostinho, A. A., S. M. Thomaz & L. C. Gomes, 2004c. Threats for biodiversity in the floodplain of the Upper Paraná River: effects of hydrological regulation by dams. Ecohydrology & Hydrobiology 4: 255–268.
Agostinho, A. A., L. C. Gomes & F. M. Pelicice, 2007. Ecologia e manejo de recursos pesqueiros em reservatórios do Brasil. EDUEM - Maringá State University Press, Maringá, Brazil.
Agostinho, A. A., F. M. Pelicice & L. C. Gomes, 2008. Dams and the fish fauna of the Neotropical region: impacts and management related to diversity and fisheries. Brazilian Journal of Biology 68: 1119–1132.
Agostinho, A. A., L. C. Gomes, N. C. L. Santos, J. C. Ortega & F. M. Pelicice, 2016. Fish assemblages in Neotropical reservoirs: colonization patterns, impacts and management. Fisheries Research 173: 26–36.
Amarasekera, K. N., R. F. Lee, E. R. Williams & E. A. B. Eltahir, 1997. ENSO and the natural variability in the flow of tropical rivers. Journal of Hydrology 200: 24–39.
Anderson, M. J., K. E. Ellingsen & B. H. McArdle, 2006. Multivariate dispersion as a measure of beta diversity. Ecology Letters 9: 683–693.
Antico, A., R. O. Aguiar & M. L. Amsler, 2018. Hydrometric data rescue in the Paraná River Basin. Water Resources Research 54: 1368–1381.
Aspin, T. W. H., K. Hart, K. Khamis, A. M. Milner, M. J. O’Callaghan, M. Trimmer, Z. Wang, G. M. D. Williams, G. Woodward & M. E. Leder, 2019. Drought intensification alters the composition, body size, and trophic structure of invertebrate assemblage in a stream mesocosm experiment. Freshwater Biology 64: 750–760.
Bailly, D., A. A. Agostinho & H. I. Suzuki, 2008. Influence of the flood regime on the reproduction of fish species with different reproductive strategies in the Cuiabá River, Upper Pantanal, Brazil. River Research and Applications 24: 1218–1229.
Baumgartner, G., K. Nakatani, L. C. Gomes, A. Bialetzki & P. V. Sanches, 2004. Identification of spawning sites and natural nurseries of fishes in the upper Paraná River, Brazil. Environmental Biology of Fishes 71: 115–125.
Baumgartner, M. T., A. G. Oliveira, A. A. Agostinho & L. C. Gomes, 2018. Fish functional diversity responses following flood pulses in the upper Paraná River floodplain. Ecology of Freshwater Fish 27: 910–919.
Berri, G. J., M. A. Ghietto & N. O. Garcia, 2002. The influence of ENSO in the Flows of the Upper Paraná River of South America over the Past 100 Years. Journal of Hydrometeorology 3: 57–65.
Berryman, A. & P. Turchin, 2001. Identifying the density-dependent structure underlying ecological time series. Oikos 92: 265–270.
Booth, D. J., N. Bond & P. Macreadie, 2011. Detecting range shifts among Australian fishes in response to climate change. Marine and Freshwater Research 62: 1027–1042.
Brander, K. M., 2007. Global fish production and climate change. Proceedings of the National Academy of Sciences 104: 19709–19714.
Camacho Guerreiro, A. I., S. A. Amadio, N. N. Fabré & V. Silva Batista, 2020. Exploring the effect of strong hydrological droughts and floods on populational parameters of Semaprochilodus insignis (Actinopterygii: Prochilodontidae) from the Central Amazonia. Environment, Development and Sustainability 1-11.
Camilloni, I. & V. Barros, 2000. The Parana River response to El Nino 1982–83 and 1997–98 events. Journal of Hydrometeorology 1: 412–430.
Caviedes, C. N., 2001. EI Nino in History: Storming Through the Ages. University Press of Florida, Florida, USA.
Chatfield, C. & H. Xing, 2019. The Analysis of Time Series: An Introduction with R. CRC Press, New York, USA.
Cheng, F., W. Li, L. Castello, B. R. Murphy & S. Xie, 2015. Potential effects of dam cascade on fish: lessons from the Yangtze River. Reviews in Fish Biology and Fisheries 25: 569–585.
Chiew, F. H. S., T. C. Piechota, J. A. Dracup & T. A. McMahon, 1998. El Nino/Southern Oscillation and Australian rainfall, streamflow and drought: links and potential for forecasting. Journal of Hydrology 204: 138–149.
Collins, M., S. I. An, W. Cai, A. Ganachaud, E. Guilyardi, F. F. Jin, M. Jochum, M. Lengaigne, S. Power, A. Timmermann, G. Vecchi & A. Wittenberg, 2010. The impact of global warming on the tropical Pacific Ocean and El Niño. Nature Geoscience 3: 391–397.
Dai, A., T. Qian, K. E. Trenberth & J. D. Milliman, 2009. Changes in Continental Freshwater Discharge from 1948 to 2004. Journal of Climate 22: 2773–2792.
Dudgeon, D., A. H. Arthington, M. O. Gessner, Z. I. Kawabata, D. J. Knowler, C. Lévêque, R. J. Naiman, A. H. Prieur-Richard, D. Soto, M. L. J. Stiassny & C. A. Sullivan, 2006. Freshwater biodiversity: importance, threats, status and conservation challenges. Biological reviews 81: 163–182.
Fernandes, R., A. A. Agostinho, E. A. Ferreira, C. S. Pavanelli, H. I. Suzuki, D. P. Lima & L. C. Gomes, 2009. Effects of the hydrological regime on the ichthyofauna of the riverine environments of the Upper Paraná River floodplain. Brazilian Journal of Biology 69: 669–680.
Garcia, A. M., J. P. Vieira, K. O. Winemiller & A. M. Grimm, 2004. Comparison of 1982-1983 and 1997-1998 El Niño effects on the shallow-water fish assemblage of the Patos Lagoon estuary (Brazil) Estuaries 27: 905-914.
Garcia, A. M., J. P. Vieira & K. O. Winemiller, 2001. Dynamics of the shallow-water fish assemblage of the Patos Lagoon estuary (Brazil) during cold and warm ENSO episodes. Journal of Fish Biology 59: 1218–1238.
Garcia, A. M., J. P. Vieira & K. O. Winemiller, 2003. Effects of 1997–1998 El Niño on the dynamics of the shallow-water fish assemblage of the Patos Lagoon Estuary (Brazil). Estuarine, Coastal and Shelf Science 57: 489–500.
Glynn, P. W., A. B. Mones, G. P. Podestá, A. Colbert & M. W. Colgan, 2017. El Niño-Southern Oscillation: effects on Eastern Pacific coral reefs and associated biota. In Glynn, P. W., D. Manzello & I. C. Enochs (eds), Coral reefs of the Eastern Tropical Pacific. Springer, Dordrecht.
Godínez-Dominguez, E., J. Rojo-Vázquez, V. Galván-Piña & B. Aguilar-Palomino, 2000. Changes in the structure of a coastal fish assemblage exploited by a small scale gillnet fishery during an El Nino–La Nina event. Estuarine, Coastal and Shelf Science 51: 773–787.
Gomes, L. C. & A. A. Agostinho, 1997. Influence of the flooding regime on the nutritional state and juvenile recruitment of the curimba, Prochilodus scrofa, Steindachner, in upper Paraná River, Brazil. Fisheries Management and Ecology 4: 263–274.
Graça, W. J. & C. S. Pavanelli, 2007. Peixes da planície de inundação do alto rio Paraná e áreas adjacentes. EDUEM - Maringá State University Press, Maringá, Brazil.
Granzotti, R. V., L. E. Miranda, A. A. Agostinho & L. C. Gomes, 2018. Downstream impacts of dams: shifts in benthic invertivorous fish assemblages. Aquatic Sciences 80: 1–28.
Grimm, A. M. & R. G. Tedeschi, 2008. ENSO and extreme rainfall events in South America. Journal of Climate 22: 1589–1609.
Gubiani, E. A., L. C. Gomes, A. A. Agostinho & E. K. Okada, 2007. Persistence of fish populations in the upper Paraná River: effects of water regulation by dams. Ecology of Freshwater Fish 16: 191–197.
Henderson, D. S., C. D. Kummerow & W. Berg, 2018. ENSO influence on TRMM Tropical oceanic precipitation characteristics and rain rates. Journal of Climate 31: 3979–3998.
Hollowed, A. B., S. R. Hare & W. S. Wooster, 2001. Pacific Basin climate variability and patterns of Northeast Pacific marine fish production. Progress in Oceanography 49: 257–282.
Holmgren, M., M. Scheffer, E. Ezcurra, J. R. Gutiérrez & G. M. Mohren, 2001. El Niño effects on the dynamics of terrestrial ecosystems. Trends in Ecology & Evolution 16: 89–94.
Huang, B. Y., P. W. Thorne, V. F. Banzon, T. Boyer, G. Cherupin, J. H. Lawrimore, M. J. Menne, T. M. Smith, R. S. Vose & H. M. Zhang, 2017. Extended reconstructed sea surface temperature, version 5 (ERSSTv5): upgrades, validations, and intercomparisons. Journal of Climate 30: 8179–8205.
Keener, V. W., G. W. Feyereisen, U. Lall, J. W. Jones, D. D. Bosch & R. Lawrence, 2010. El-Niño/Southern Oscillation (ENSO) influences on monthly NO3 load and concentration, stream flow and precipitation in the Little River Watershed, Tifton, Georgia (GA). Journal of hydrology 381: 352–363.
Lehodey, P., J. Alheit, M. Barange, T. Baumgartner, G. Beaugrand, K. Drinkwater, J. M. Fromentin, S. R. Hare, G. Ottersen, R. I. Perry, C. Roy, C. D. Van-der-Linger & F. Werner, 2005. Climate variability, fish, and fisheries. Journal of Climate 19: 5009–5030.
Lingard, H., M. Hallowell, R. Salas & P. Pirzadeh, 2017. Leading or lagging? Temporal analysis of safety indicators on a large infrastructure construction project. Safety science 91: 206–220.
Makrakis, S., A. P. Bertão, J. F. Silva, M. C. Makrakis, F. Sanz-Ronda & L. F. Celestino, 2019. Hydropower Development and Fishways: A Need for Connectivity in Rivers of the Upper Paraná Basin. Sustainability 11: 1–24.
Mol, J. H., D. Resida, J. S. Ramlal & C. R. Becker, 2000. Effects of El Niño-related drought on freshwater and brackish-water fishes in Suriname, South America. Environmental Biology of Fishes 59: 429–440.
Moraes, L. E., E. Paes, A. Garcia, O. Möller Jr. & J. Vieira, 2012. Delayed response of fish abundance to environmental changes: a novel multivariate time-lag approach. Marine Ecology Progress Series 456: 159–168.
Moran, E. F., M. C. Lopez, N. Moore, N. Müller & D. W. Hyndman, 2018. Sustainable hydropower in the 21st century. Proceedings of the National Academy of Sciences 115: 11891–11898.
Oliveira, A. G., H. I. Suzuki, L. C. Gomes & A. A. Agostinho, 2015. Interspecific variation in migratory fish recruitment in the Upper Paraná River: effects of the duration and timing of floods. Environmental Biology of Fishes 98: 1327–1337.
Oliveira, A. G., M. T. Baumgartner, L. C. Gomes, R. M. Dias & A. A. Agostinho, 2018. Long-term effects of flow regulation by dams simplify fish functional diversity. Freshwater Biology 63: 293–305.
Ota, R. R., G. C. Deprá, W. J. Graça & C. S. Pavanelli, 2018. Peixes da planície de inundação do alto rio Paraná e áreas adjacentes: revised, annotated and updated. Neotropical Ichthyology 16: 1–111.
Pereira, M. P. S., K. R. Mendes, F. Justino, F. Couto, A. S. Silva, D. F. Silva & A. C. M. Malhado, 2020. Brazilian dry forest (Caatinga) response to multiple ENSO: the role of Atlantic and Pacific Ocean. Science of the Total Environment 705: 1–7.
Philander, S. G. H., 1983. El Nino southern oscillation phenomena. Nature 302: 295.
Philander, S. G. H., 1990. El Niño, La Niña, and the South Oscillation. Academic Press, Massachusetts, USA.
Pineda, A., Ó. Peláez, J. D. Dias, B. T. Segovia, C. C. Bonecker, L. F. M. Velho & L. C. Rodrigues, 2019. The El Niño Southern Oscillation (ENSO) is the main source of variation for the gamma diversity of plankton communities in subtropical shallow lakes. Aquatic Sciences 81: 49.
Ponton, D., 2001. ENSO and the hydrology of the Sinnamary River (French Guiana) during the rainy season: will future El Nino events increase the impact of the Petit Saut dam on downstream fish communities? Archiv fur Hydrobiologie 152: 451–468.
Possamai, B., J. P. Vieira, A. M. Grimm & A. M. Garcia, 2018. Temporal variability (1997-2015) of trophic fish guilds and its relationships with El Niño events in a subtropical estuary. Estuarine, Coastal and Shelf Science 202: 145–154.
Probst, W. N., V. Stelzenmüller & H. Fock, 2012. Using crosscorrelations to assess the relationship between time-lagged pressure and state indicators: An exemplary analysis of North Sea fish population indicators. ICES Journal of Marine Science 69: 670–681.
Quirino, B. A., N. Carniatto, S. M. Thomaz & R. Fugi, 2018. Small fish diet in connected and isolated lakes in a Neotropical floodplain. Ecology of Freshwater Fish 28: 97–109.
R Core Team, 2020. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria.
Rodrigues, A. C., B. M. Dal Vesco, C. M. Muniz, C. P. Nascimento, G. F. Andreotti, J. C. Alves, M. H. F. Cardoso, M. H. Schmitz, M. J. M. Ganassin, M. P. Camargo, M. G. Buzo, M. T. Baumgartner, N. C. L. Santos, A. A. Agostinho & L. C. Gomes, 2020. How limnological variables influence the occurrence and abundance of a visual predator in an invaded river-floodplain system. Oecologia Australis 24: 461–473.
Rodríguez, M. A. & W. M. Lewis, 1997. Structure of fish assemblages along environmental gradients in floodplain lakes of the Orinoco River. Ecological Monographs 67: 109–128.
Ropelewski, C. F. & M. S. Halpert, 1987. Global and regional scale precipitation patterns associated with the El Niño/Southern Oscillation. Monthly Weather Review 115: 1606–1626.
Smolders, A. J. P., G. Van der Velde, J. G. M. Roelofs & M. G. Hiza, 2000. El Niño caused collapse of the Sabalo fishery (Prochilodus lineatus, Pisces: Prochilodontidae) in a South American river. Naturwissenschaften 87: 30–32.
Stevaux, J. C., D. P. Martins & M. Meurer, 2009. Changes in a large regulated tropical river: the Paraná River downstream from the Porto Primavera dam, Brazil. Geomorphology 113: 230–238.
Su, L., C. Miao, D. Kong, Q. Duan, X. Lei, Q. Hou & H. Li, 2018. Long-term trends in global river flow and the causal relationships between river flow and ocean signals. Journal of Hydrology 563: 818–833.
Suzuki, H. I., A. A. Agostinho, D. Bailly, M. F. Gimenes, H. F. Júlio-Junior & L. C. Gomes, 2009. Inter-annual variations in the abundance of young-of-the-year of migratory fishes in the Upper Paraná River floodplain: relations with hydrographic attributes. Brazilian Journal of Biology 69: 649–660.
Suzuki, F. M., L. V. Pires & P. S. Pompeu, 2011. Passage of fish larvae and eggs through the Funil, Itutinga and Camargos Reservoirs on the upper Rio Grande (Minas Gerais, Brazil). Neotropical Ichthyology 9: 617–622.
Swales, S., A. W. Storey, I. D. Roderick & B. S. Figa, 1999. Fishes of floodplain habitats of the Fly River system, Papua New Guinea, and changes associated with El Niño droughts and algal blooms. Environmental Biology of Fishes 54: 389–404.
Terra, B. D. F., A. B. I. D. Santos & F. G. Araújo, 2010. Fish assemblage in a dammed tropical river: an analysis along the longitudinal and temporal gradients from river to reservoir. Neotropical Ichthyology 8: 599–606.
Trenberth, K. E., 1997. The definition of El Niño. Bulletin of the American Meteorological Society 78: 2771–2778.
Tundisi, J. G. & M. Straskraba, 1999. Theoretical Reservoir Ecology and Its Applications. Backhuys Publishers, Leiden, The Netherlands.
Turesson, H. & C. Bronmark, 2007. Predator-prey encounter rates in freshwater piscivores: effects of prey density and water transparency. Oecologia 153: 281–290.
Turgeon, K., C. Turpin & I. Gregory-Eaves, 2019. Dams have varying impacts on fish communities across latitudes: a quantitative synthesis. Ecology Letters 22: 1501–1526.
Ward, J. V. & J. A. Stanford, 1995. Ecological connectivity in alluvial river ecosystems and its disruption by flow regulation. Regulated Rivers: Research & Management 11: 105–119.
Watters, G. M., R. J. Olson, R. C. Francis, P. C. Fiedler, J. J. Polovina, S. B. Reilly, K. Y. Aydin, C. H. Boggs, T. T. Essington, C. J. Walters & J. F. Kitchell, 2003. Physical forcing and the dynamics of the pelagic ecosystem in the eastern tropical Pacific: simulations with ENSO-scale and global-warming climate drivers. Canadian Journal of Fisheries and Aquatic Sciences 60: 1161–1175.
Acknowledgements
The authors thank the Long Term Ecological Research, Site 6, supported by the National Council for Scientific and Technological Development (“Conselho Nacional de Desenvolvimento Científico e Tecnológico” – CNPq) of Brazil for the financial support. The “Programa de Excelência Acadêmica” (PROEX) of the “Coordenação de Aperfeiçoamento de Pessoal de Nível Superior” (CAPES) and the Núcleo de Pesquisas em Limnologia, Ictiologia e Aquicultura (Nupélia - UEM) also provided financial support and infrastructure for developing this study. JCA received fellowship from CAPES.
Funding
The authors thank Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for providing grants.
Author information
Authors and Affiliations
Contributions
Conceived and designed the investigation: JCA, GFA, AAA, LCG Performed field and/or laboratory work: JCA, GFA, LCG. Analyzed the data: JCA, GFA, LCG. Wrote and revised the paper: JCA, GFA, AAA, LCG.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Handling editor: Fernando M. Pelicice
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Alves, J.C., Andreotti, G.F., Agostinho, A.A. et al. Effects of the El Niño Southern Oscillation (ENSO) on fish assemblages in a Neotropical floodplain. Hydrobiologia 848, 1811–1823 (2021). https://doi.org/10.1007/s10750-021-04555-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-021-04555-4