
Abstract
Rapid development has fragmented Singapore’s coastal habitats and converted them into artificial shorelines, with implications for the ecosystem services they provide. This study investigated the impact of urban development on two ecosystem services related to fisheries (direct provisioning and indirect nursery functions) by comparing adjacent mangrove and artificial rocky shore sites at two locations. In terms of fisheries provisioning ecosystem services, fish video data indicated that fish assemblage structure was significantly different between the two habitat types, with Ellochelon vaigiensis, Ambassis kopsii, Ambassis interrupta, and Zenarchopterus buffonis contributing most significantly. Trap data indicated that there were significantly more fish on the artificial rocky shore than mangrove at one of our locations in Pasir Ris. There were also significant differences in the size distribution of fish between habitat types at both locations, with high proportions of smaller fish in the mangroves. Even with higher juvenile presence, the role of urban mangroves in providing nursery ecosystem services is less clear, since we cannot determine whether this habitat type is essential for any individual fish species. This study indicates that mangroves provide a level of fisheries ecosystem services as part of a heavily developed and fragmented landscape in conjunction with other substrate types.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Intertidal mangrove forests that fringe tropical and subtropical coastlines are now commonly examined from an ecosystem services perspective. The aim of such a framework is to quantify the roles and benefits that ecosystems provide to human populations (e.g., Millennium Ecosystem Assessment, 2005). For mangroves, these are diverse and commonly include direct provisioning services such as timber, fuel and lime render (Bandaranayake, 1998; Dahdouh-Guebas et al., 2000; Scales et al., 2017), and indirect regulating services related to wave attenuation (Barbier, 2016) and carbon sequestration and storage (Nam et al., 2016), amongst others. Additionally, mangroves provide a range of tangible and abstract cultural ecosystem services (James et al., 2013) to hundreds of millions of people throughout the tropics.
Within mangrove ecosystem services research, the role of mangroves in supporting tropical fisheries is a foundational paradigm (Alongi, 2009; Lee et al., 2014; Whitfield, 2017). Mangrove–fisheries linkages can be viewed through the lens of direct provisioning ecosystem services, where the mangrove ecosystem provides fish and aids food security. Mangrove–fisheries linkages can also be considered as an indirect regulating ecosystem service; mangroves can act as a nursery habitat for juvenile fish that provides food and acts as a refuge from predators due to the complex habitat structure (Manson et al., 2005; Nagelkerken et al., 2010; Igulu et al., 2014). This ultimately leads to an increase in provisioning services to humans. The mangroves–fisheries paradigm has important implications for fisheries, as some studies have suggested positive relationships between mangrove area and fish yield for both coastal and offshore fisheries (e.g., Loneragan et al., 2005; Aburto-Oropeza et al., 2008). However, the way in which fish spatially utilize mangroves is likely to be more complex than previously thought (e.g., Lee et al., 2014; Sheaves et al., 2016).
Although we still lack key data on how patterns vary between regions, results from meta-analyses of mangrove–fishery linkages at a global level indicate that mangroves do have a considerable effect on fisheries. For example, Carrasquilla-Henao & Juanes, (2017) demonstrated that mangrove area is a highly significant predictor of fishery catches in a variety of mangrove settings across the world. These findings potentially provide incentives for mangrove habitat conservation to assist in the stabilization of declining fish populations. However, evaluating the nursery role of mangroves is complex given that the mechanisms behind fishery yields are difficult to predict. Given the tendency of fish to migrate between multiple interacting habitat types and locations (Sheaves, 2005), it is difficult to quantify the extent to which fish rely on any specific habitat type, such as a mangrove or rocky shore. This is further complicated by the fact that species tend to utilize mangroves for only part of their life cycle, and even then only when they are accessible during favorable tidal conditions, as well as using other habitats such as adjacent seagrass beds and coral reefs (Alongi, 2009). As such, the extent to which mangroves provide nursery habitat compared to other natural and artificial shorelines in the coastal landscape, how this changes from species to species, and the ecological aspects of fishery linkages are still largely unclear, despite positive correlations in different geographical regions (Manson et al., 2005; Carrasquilla-Henao & Juanes, 2017).
Urban mangrove systems provide a unique experimental setting with which to test mangrove–fishery linkages that usually occur over larger scales, as coastlines with fragmented mangrove patches within a matrix of artificial structures will alter habitat availability for fish populations. The aim of this study was to provisionally examine the role of mangroves in supporting fisheries ecosystem services along a stretch of urban coastline in Singapore, by quantifying differences in fish community composition and the size of individuals between mangrove and adjacent artificial rocky shore sites. This aim was undertaken from an ecosystem services perspective, considering the differences between the direct provisioning and indirect nursery regulating services contributed by each habitat type.
Methods
Study locations
Singapore is a highly urbanized City-State located in Southeast Asia, experiencing a tropical rainforest climate characterized by hot, humid weather year-round with no distinct seasons and little variation in climatic conditions. As much as 75 km2 of Singapore’s land area was originally covered with coastal mangrove forest, though this has now been reduced to an estimated 6.6 km2 today (Yee et al., 2010). This loss was primarily due to reservoir construction, land reclamation, and coastal urban development, particularly after national Independence in 1965 (Friess et al., 2012). 63.3% of Singapore’s coastline is now completely artificial, lined by seawalls, revetments, and other artificial rocky shores (Lai et al., 2015). As much as 30% of Singapore’s remaining mangrove is expected to be lost by the year 2030 according to publicly available government planning documents, so the proportion of artificial habitat is expected to increase further (Lai et al., 2015).
While knowledge of indirect nursery ecosystem services is lacking in Singapore, the history of direct provisioning ecosystem services related to fisheries is summarized by Friess (2017), with fisheries having been important in the Singapore context for centuries (Gibson-Hill, 1950). As recently as the 1970s, fish caught by Singaporean trawlers accounted for 50% of local consumption (Hill, 1980). Locally caught fish accounts for 6% of consumption today, with plans to increase this further in the context of local food security, including growth in aquaculture production (AVA, 2015). Fisheries are also important for economic security, with the sector estimated to directly employ 14,000 people (Teh & Sumaila, 2013).
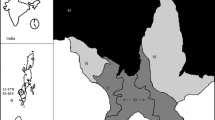
Field data collection was conducted at two coastal locations in northeast Singapore within the eastern Johor Strait, Pulau Ubin and Pasir Ris Park (Fig. 1). Two different sites were sampled at each location, with a total of four sites, corresponding to either a mangrove with muddy substrate, or an artificial rocky shore consisting of a cobble/boulder matrix. Both mangrove sites were situated within a mangrove creek and both artificial rocky sites were situated on the open coastline adjacent to these mangrove creeks. All study locations experience the same mixed semidiurnal tides and a maximum tidal range of ~3 m. The presence of a manmade causeway across the Johor Strait limits water movement through the whole Strait, resulting in the tidal influence being predominantly from the east shore (see Table 1 for additional physical site characteristics).

Locations of the four study sites: Ubin mangrove, Ubin artificial rocky shore, Pasir Ris artificial rocky shore, and Pasir Ris mangrove. Inset images: stars indicate individual sites; white area indicates mangrove
Both mangrove locations are currently managed by the National Parks Board, Government of Singapore. Mangrove sites were chosen to be as structurally similar as possible, with the sampled fringes composed primarily of Rhizophora apiculata Blume at similar vegetation densities. Mangrove widths were also similar at the two sites, ~122 m at Pulau Ubin and ~98 m at Pasir Ris. Similarly, the artificial rocky shore sites were chosen to be as comparable to each other as possible within the constraints of habitat available for sampling. Both artificial sites are structurally similar, made of the same material and using the same construction techniques. The artificial sites were situated at distances of ~1,200 m and ~650 m, respectively, from the mangroves, separated by stretches of mud and sand foreshore. The exact locations of the artificial sites were constrained by habitat availability along the coast. The locations were separated from each other by Serangoon Harbour, a natural channel with a maximum depth <25 m C.D. (below Chart Datum). Subtidal areas offshore of all sites were predominantly made up of soft sediment, gradually sloping down to ~10 m C.D. at least 300 m away from the shoreline, with no permanent subtidal habitats (such as seagrass and coral) present.
Fish video sampling
The fish community at each site was surveyed using a baited underwater video system (BRUV) (sensu Cappo et al., 2007), a commonly used method of surveying fish associated with a variety of habitat types (Murphy & Jenkins, 2010). Each BRUV consisted of a GoPro Hero camera (model# HWBL1) with a wide angle setting (1080p resolution, 30 frames per second, 170° field of view), on a frame composed of an 85 cm long PVC pipe crossbar situated 20 cm above the seabed on legs (Fig. 2). Prior to deployment a plastic mesh bait basket (5 mm mesh diameter) was attached 52 cm away from the camera on top of the crossbar.

Diagrammatic representation of BRUV frame used in present study
All sampling was conducted between December 2015 and June 2016, during daytime hours, on a rising or high tide. Each site was surveyed on five regular but randomly selected dates throughout the sampling period, to cover a range of variable tidal conditions. A range of tidal heights were sampled to gain a representative understanding of fisheries under different conditions. While multiple sites were not sampled on the same date, care was taken to ensure that they were sampled as close as possible to each other to reduce possible confounding factors. On each date, five BRUVs were concurrently deployed ~1.5 h before high tide, perpendicular to the shore. BRUVS were fully submerged on deployment and on retrieval 1 h later, water depth was <2 m. We omitted video data from further analysis for two sample days where high turbidity and poor visibility impacted the ability to clearly view the bait basket and nearby fish in the field of view.
BRUVs were baited with 80 g of canned wet cat food, consisting of mackerel/sardine in jelly, with a moisture content of 82%, protein 12%, fiber 1%, and fat 1%. The canned food was found in preliminary trials during this study to be more effective than fresh bait, and previous studies have found bait comprising of oily fish to be highly effective (Whitelaw et al., 1991; Wraith et al., 2013). Baiting techniques can potentially produce biased estimates of small-scale habitat use by drawing fish in from adjacent habitats (Bassett & Montgomery, 2011). Nevertheless, we deployed baited cameras because the water turbidity rendered it difficult to observe naturally passing fish using an un-baited technique (Pers. Obs.). The caveat of this method is that samples from baited video are often presumed to be biased towards predatory or scavenging species and exclude herbivorous or omnivorous species (Harvey et al., 2007). However, others have demonstrated the use of bait in increasing the ability to discriminate mixed-diet fish assemblages due to the increased numbers of individuals and species sampled at the baited stations (Watson et al., 2005; Cappo et al., 2007; Harvey et al., 2007).
Video data analysis
Fish were identified to species level where possible from each 1 h video. Fish abundance was assessed with the commonly used MaxN method (Cappo et al., 2007; Murphy & Jenkins, 2010), whereby fish abundance was recorded for each species as the greatest number of individuals in a single frame using the software package Event Measure (SeaGIS. Pty. Ltd, Australia). Variation in assemblage structure was analyzed with a variety of non-parametric techniques. Bray–Curtis dissimilarities were calculated using square-root transformed MaxN data and non-metric multidimensional scaling ordinations (nMDS) were constructed to visualize variation between assemblages across both locations and habitat types. nMDS stress levels <0.2 are thought to give an adequate representation of relationships between each sample (Clarke, 1993). Two-factor, crossed ANOSIM was used to further investigate differences identified from the nMDS. This analysis was performed on the dissimilarity data from all sampling dates to examine differences between locations (across both habitat types) and between habitat types (across both locations). This was followed by SIMPER analysis to examine the contribution of individual species to observed differences (PRIMER v6, Clarke, 1993).
Spatial variation in total species richness and fish abundance (recorded as MaxN) were analyzed from the BRUV videos. Two-factor ANOVAs were used to examine differences between the locations at Pasir Ris and Pulau Ubin (two levels: random) and habitat type, i.e., mangrove and artificial rocky shores (two levels: orthogonal and fixed). The null hypotheses being that there were no significant differences in total species richness or fish abundance between either the locations or habitat type. Eighteen replicates were randomly selected from the total cameras and traps deployed in each habitat type at each location due to trap damage during occasional deployments. Only those species that were observed on more than 5 occasions were analyzed.
Data for all ANOVAs were checked for homogeneity of variances using Cochran’s C-test, and Ln(x + 1) transformations were performed to homogenize variances. Significant differences were further examined, when appropriate, using the Student–Newman–Keuls (SNK) multiple comparison tests (GMAV5, EICC, The University of Sydney). The fragmented nature of Singapore’s coastal ecosystems and issues with access to many areas constrained the sampling design. Limited spatial replication of habitat types reduced the power of these tests, potentially lessening the ability to identify differences between habitat types.
Trap surveys
Trap surveys were carried out at each sampling site on four dates during the same periods as BRUVs were deployed. Each survey consisted of 5 concurrent 1-h deployments of baited mesh fish traps (75 cm × 75 cm base, 27 cm height, 4 mm mesh size, with 4 × 15 cm diameter entrances). Traps were baited similarly to the BRUVs and deployed haphazardly throughout the range of microhabitats within a ~100 m area at each site, with a minimum distance of 3 m between traps in <2 m of water. The aim of the varied trap deployments was to survey a more diverse and representative catch in order to gain a better perspective on the variety of fish species located within the four sites. Once traps were retrieved, fish were identified to species level and total length was recorded prior to immediate release.
Trap data analysis
Spatial variation in fish abundance recorded in traps was analyzed using a two-factor ANOVA to determine differences between the two locations at Pasir Ris and Pulau Ubin (two levels: random) and habitat type, i.e., mangrove and artificial rocky shores (two levels: orthogonal and fixed). The null hypotheses being that there were no significant differences in fish abundance between either the locations or habitat type. Eighteen replicates were randomly selected from the total traps laid in each location.
Data for all ANOVAs were checked for homogeneity of variances (Cochran’s C-test) and Ln(x + 1) transformations were performed to homogenize variances. Significant differences were further examined when appropriate using SNK multiple comparison tests. The fragmented nature of Singapore’s coastal ecosystems and issues with access to many areas constrained the sampling design. Limited spatial replication of habitat types reduced the power of these tests, potentially lessening the ability to identify differences between habitat types. Kolmogorov–Smirnov two sample tests of length frequency were used to determine if there were significant differences between the size distributions of fish at each site.
Results
Fish video data: differences in fish assemblage structure
There were significant differences between mangrove and artificial rocky shore fish assemblages both within and between locations (Fig. 3). The ANOSIM analysis indicated that assemblage structure varied significantly between locations across all habitat types (Global R = 0.391, P = 0.001) and between habitats across all locations (Global R = 0.544, P = 0.001).

nMDS ordination comparing the similarity of fish assemblages in mangrove and artificial rocky shore habitats at Pasir Ris and Pulau Ubin, calculated from MaxN data. Increasing distance between points indicates decreasing similarity between fish assemblages
The species that made the most significant contributions to differences in assemblage structure between the two habitat types, were Ellochelon vaigiensis Quoy & Gaimard, 1825 (20.77%), Ambassis kopsii Bleeker, 1858 (17.9%), Ambassis interrupta Bleeker, 1853 (14.21%), and Zenarchopterus buffonis Valenciennes, 1847 (11.65%). These same species were also responsible for the greatest differences in assemblage structure between the locations, although to differing degrees, e.g., A. interrupta (19.31%), A. kopsii (14.3%), E. vaigiensis (12.85%), and Z. buffonis (8.45%) (Fig. 4).

Summary of similarity percentage (SIMPER) analyses for species contributions to observed differences between habitat types (A) and between sites (B)
Fish video data: spatial variation in species richness and fish abundance
There were no significant differences in species richness or abundance of Scatophagus argus Linnaeus, 1766, and Abudefduf bengalensis Bloch, 1787, between locations or habitat types (Table 2A). For Z. bufonis, there was a significant interaction between location and habitat type (P = 0.0480), with significantly more individuals in mangroves than artificial rocky shores at Pasir Ris, but no significant difference in abundances between habitat types at Pulau Ubin (Table 2B). For A. interrupta, there was a significant difference in abundances between locations (P = 0.0041), but not between habitat types (Table 2B). For A. kopsii, there were significant differences in abundances between locations (P = 0.0005) and habitats (P = 0.0133) and a consistent pattern with higher abundances in mangroves than at artificial rocky shores at both locations (Table 2C). At each location, the mean abundances were higher in mangroves than rocky shores for all five species evaluated (Table 3).
Fish trap data: spatial variation in species richness and fish abundance from traps
The ANOVA found significant differences in fish abundances between sites, with an interaction between habitat type and location for both total fish (P = 0.0282) and A. kopsii (P = 0.0371) (Table 2D). In both cases there were significantly more fish on the artificial rocky shores than mangroves at Pasir Ris, while there was no significant difference in numbers between the habitats at Pulau Ubin. The mean abundances were higher in rocky shores for A. kopsii at Pasir Ris, but higher in mangroves at Pulau Ubin. E. vaigiensis was only recorded on rocky shores at Pasir Ris. Abundances of A. interrupta, Stigmatogobius sadanundio Hamilton, 1822, and Butis butis Hamilton, 1822, were higher in mangroves (Table 4).
Fish trap data: length frequency differences
Kolmogorov–Smirnov tests of length frequency between fish at mangrove and artificial rocky shore sites indicated that there was a significant difference in the size distribution of fish between habitat types, with high proportions of smaller fish in the mangroves (Fig. 5).

Kolmogorov–Smirnov two sample tests of length frequency between fish, for all species combined at all sites (D is: 0.1040, P of: 0.047) (A), A. kopsii combined at all sites (D is: 0.1782, P of: 0.003) (B), all species at Pasir Ris sites (D is: 0.1828, P of: 0.003) (C), A. kopsii at Pasir Ris sites (D, is: 0.2713, P of: <0.001) (D), all species at Pulau Ubin sites (D is: 0.8168, P of: <0.001) (E), and A. kopsii at Pulau Ubin sites (D is: 0.7600, P of: <0.001) (F)
Discussion
Habitat differences in fish assemblage structure and contributions to direct provisioning ecosystem services
While practical difficulties of habitat availability constrained replication in the present study, we identified significant differences between fish assemblages at adjacent mangrove and artificial rocky shore sites. This indicates that the two habitat types may provide different, yet complimentary, direct provisioning services to local fish populations in Singapore. Indeed, studies elsewhere indicate that such artificial hard substrates can themselves support diverse, abundant assemblages of fish (e.g., Weis & Weis, 2005; Waltham & Connolly, 2007). At the mangrove sites, the genera that contributed the most to differences in fish assemblages between locations and habitat types were the Ambassidae, particularly A. kopsii and A. interrupta species that are of minor commercial value elsewhere in Southeast Asia (Froese & Pauly, 2017). Of the other main species that contributed to differences between habitats, S. argus is of limited large-scale commercial food value due to its small size, but it is harvested on local scales in small-scale artisanal fisheries, and is also used in traditional Chinese medicine and the aquarium trade (Collen et al., 2010), along with B. butis (Monks, 2006). The mangroves supporting these three species therefore provide a level of direct provisioning ecosystem services. The artificial rocky shore sites, particularly at Pasir Ris, also contributed provisioning services. For example, the mullet E. vaigiensis, an important commercial species of both capture fisheries and aquaculture, was commonly recorded around this habitat type. Mullet products such as roe, testes, and stomach are popular items in restaurants and markets (Crosetti & Blaber, 2015), while regionally, E. vaigiensis is also taken as a food fish (Harrison & Senou, 1999).
Habitat differences in fish assemblage structure and contributions to indirect nursery ecosystem services
Both the mangrove and the artificial rocky shore habitats could also be considered as providing more indirect fish nursery regulation services, as they contained fish species that served as food for larger, carnivorous fish higher up in the food chain (Sheaves & Molony, 2000). For the most part, the juvenile fish we observed at all sites held commercial importance insofar as they were a primary food source for many commercially important fish (Harrison & Senou, 1999; Monks, 2006; Collen et al., 2010; Froese & Pauly, 2017). There were also distinctly different fish assemblage structures between our sites, with higher proportions of fish in smaller size classes in the mangroves. A study by Shinnaka et al. (2007) in Thailand also compared fish populations across habitat types, finding distinct differences in fish assemblages between mangrove sites and adjacent sites cleared of mangroves. There were also significant differences in length frequency distributions for the most abundant species, with smaller individuals found at the mangrove site. Specifically, for three of the four most abundant species collected, there were higher proportions of small individuals at the mangrove site than the cleared site (Shinnaka et al., 2007).
While the video results for the abundance estimates of the dominant species, A. kopsii, demonstrated a consistent pattern of higher abundances in mangroves than at the artificial rocky shores, the traps contained significantly higher abundances of individuals at the artificial rocky shore than mangroves at Pasir Ris. Further analysis of size classes from traps indicates that this was only apparent in larger individuals, with a higher proportion of small fish occurring in the mangroves at both sites.
Debates on the role of mangroves as fisheries provisioning or regulating services
The size class found in the mangrove was predominantly juveniles (see Froese & Pauly, 2017 for overviews of species life history), suggesting a potential nursery value of this habitat compared to the artificial rocky shore sites. This seems to be a broader pattern, as other regions have also found that fish species use shallow water biotopes such as mangroves and seagrass beds as nurseries during their juvenile stage, but migrate permanently to deeper habitats when reaching a specific size class (Nagelkerken et al., 2000). The difference of smaller size classes within the mangroves suggests that the mangrove sites may be playing a role as a nursery during this life stage, where mangrove habitats are expected to protect, shelter, and provide food for juveniles (Manson et al., 2005; Meynecke et al., 2008; Froese & Pauly, 2017). However, a number of authors have emphasized that the presence of juvenile fish, proof of food utilization, and protection from predators in a particular habitat is not solely sufficient to define that habitat as a nursery area (Whitfield, 2017). Instead, additional proof must be provided that a particular type of juvenile habitat contributed a greater than average number of individuals per-unit-area to the adult population of a species than other habitat types. It is difficult to demonstrate the relative importance of the nursery function between habitat types, since there is no universal acceptance that individual habitats are essential to the juveniles of any particular fish species (Beck et al., 2001; Whitfield, 2017).
Thus, while we found that mangrove sites used in this study contained a higher proportion of smaller fish than the artificial habitat type, it is unclear from the current study whether they satisfy a nursery regulating function. To elucidate this, we need to further investigate the ecological aspects of fisheries linkages within and between nearby mangrove habitats to determine the relative nursery service contributions of various habitat types. As is the case in other estuarine ecosystems (e.g., Weinstein et al., 2005; Wasserman & Strydom, 2011), many of the advantages provided by mangroves are probably derived at an ecosystem scale, with mangroves part of a complex, life supporting habitat mosaic rather than a site-by-site effect (Nagelkerken et al., 2015).
The extent to which a nursery regulating function is a distinct ecosystem service or a biodiversity component is still under contention, since nursery services are necessary for the production of most other ecosystem services, and thus have only indirect impacts on people (Millennium Ecosystem Assessment, 2005; TEEB, 2010). The UK National Ecosystem Assessment states that the nursery function is already valued through fish that are caught and contribute to fisheries, meaning that fisheries ecosystem services are not recognized as anything more than provisioning (Haines-Young & Potschin, 2013). Liquete et al. (2016) suggest including the nursery function as an ecosystem service, but supported by an integrated ecosystem assessment that comprises at least ‘biodiversity and ecosystem condition’, so there is a clear differentiation between components and specific references to species of direct use for humans. By making a clear distinction between biodiversity and ecological integrity, it may be possible to quantify the link between biodiversity and ecosystem services, allowing ecosystem services to potentially act as a policy tool for protecting biodiversity (Liquete et al., 2016). Categorizing fisheries ecosystem services as more than just a provisioning service would encompass the wide-ranging importance of fisheries, and also represent the main themes of the mangrove–nursery literature.
Conclusions
Continuing coastal development and land cover change in Singapore (Lai et al., 2015) and throughout Southeast Asia (Richards & Friess, 2016) are leading to continued mangrove loss and conversion to artificial habitats. Singapore’s mangroves are now known to provide numerous ecosystem services to the Singaporean population (Friess, 2017), and this study suggests that this may also include a level of direct provisioning and indirect regulating ecosystem services related to fisheries.
We found significant differences between mangrove and artificial rocky shore sites within the fish assemblages and in terms of the ecosystem services provided by each habitat type. The mangrove sites provided key fishery habitat for certain mangrove-dependent species. Although fish abundances were higher at one of the artificial rocky shore sites, we found that fish species of smaller size classes were located within mangroves compared to artificial rocky shore sites, indicating a potential indirect nursery service for juvenile fish. Further investigations are necessary along urban coasts such as in Singapore with greater spatial and temporal replication to determine how mangrove-dependent species influence the broader ecosystem, how different zones within mangroves influence the types of fish caught, and how scale influences nursery services in a highly interconnected system. Even though the mechanisms behind mangrove–fisheries dynamics are still unclear, recent studies show that mangrove degradation can substantially contribute to catch declines (Carrasquilla-Henao & Juanes, 2017). Thus, it is crucial to enforce local conservation strategies to protect mangroves as critical habitats for mangrove-dependent species and species that may rely on these populations, to protect global fisheries resources. Singapore, with its rapid development and mangrove fragmentation, is an important tropical case study that allows us to understand how increasingly urbanized coastal areas may influence fish population dynamics.
References
Alongi, D. M., 2009. Paradigm shifts in mangrove biology. In Perillo, G. M. E., E. Wolanski, D. R. Cahoon & M. M. Brinson (eds), Coastal wetlands: an integrated ecosystem approach. Elsevier, Amsterdam: 615–640.
Aburto-Oropeza, O., E. Ezcurra, G. Danemann, V. Valdez, J. Murray & E. Sala, 2008. Mangroves in the Gulf of California increase fishery yields. Proceedings of the National Academy of Sciences 105: 10456–10459.
AVA, 2015. Working together as one: annual report 2014/15. Agri-Food and Veterinary Authority of Singapore, Government of Singapore. Singapore.
Bandaranayake, W. M., 1998. Traditional and medicinal uses of mangroves. Mangroves and Salt Marshes 2: 133–148.
Barbier, E. B., 2016. The protective service of mangrove ecosystems: a review of valuation methods: marine Pollution Bulletin special issue: “Turning the tide on mangrove loss”. Marine Pollution Bulletin 109: 676–681.
Bassett, D. K. & J. C. Montgomery, 2011. Investigating nocturnal fish populations in situ using baited underwater video: with special reference to their olfactory capabilities. Journal of Experimental Marine Biology and Ecology 409: 194–199.
Beck, M. W., K. L. Heck Jr., K. W. Able, D. L. Childers, D. B. Eggleston, B. M. Gillanders & R. J. Orth, 2001. The identification, conservation, and management of estuarine and marine nurseries for fish and invertebrates: a better understanding of the habitats that serve as nurseries for marine species and the factors that create site-specific variability in nursery quality will improve conservation and management of these areas. Bioscience 51: 633–641.
Cappo, M., E. Harvey & M. Shortis, 2007. Counting and measuring fish with baited video techniques – an overview. In Lyle, J. M., Furlani, D. M., Buxton, C. D. (eds), Proceedings of the 2006 Australian Society of Fish Biology Conference and Workshop Cutting-edge Technologies in Fish and Fisheries Science, Hobart, August 2006. ASFB: Hobart: 101–114.
Carrasquilla-Henao, M. & F. Juanes, 2017. Mangroves enhance local fisheries catches: a global meta-analysis. Fish and Fisheries 18: 79–93.
Clarke, K. R., 1993. Non-parametric multivariate analyses of changes in community structure. Australian Journal of Ecology 18: 117–143.
Collen, B., N. Richman, A. Beresford, A. Chenery & M. Ram, Sampled Red List Index Coordinating Team, 2010. Scatophagus argus. The IUCN Red List of Threatened Species 2010. doi:10.2305/IUCN.UK.2010-4.RLTS.T155268A4761779.en.
Crosetti, D. & S. J. Blaber, (eds), 2015. Biology, Ecology and Culture of Grey Mullets (Mugilidae). CRC Press, Boca Raton.
Dahdouh-Guebas, F., C. Mathenge, J. G. Kairo & N. Koedam, 2000. Utilization of mangrove wood products around mida creek (Kenya) amongst subsistence and commercial users. Economic Botany 54: 513–527.
Friess, D. A., 2017. Singapore as a long-term case study for tropical urban ecosystem services. Urban Ecosystems 20: 277–291.
Friess, D. A., J. Phelps, R. C. Leong, W. K. Lee, A. K. S. Wee, N. Sivasothi & E. L. Webb, 2012. Mandai mangrove, Singapore: lessons for the conservation of Southeast Asia’s mangroves. Raffles Bulletin of Zoology 25: 55–65.
Froese, R. & D. Pauly. Editors. 2017. FishBase. World Wide Web electronic publication. www.fishbase.org, version 06/2016.
Gibson-Hill, C. A., 1950. The fishing boats operated from Singapore Island. Journal of the Malayan Branch of the Royal Asiatic Society 23: 148–170.
Haines-Young, R. & M. Potschin, 2013. Common International Classification of Ecosystem Services (CICES): consultation on version 4. In: EEA Framework Contract No EEA/IEA/09/003. August–December 2012. http://cices.eu/.
Harrison, I. J. & H. Senou, 1999. Mugilidae. In: Carpenter, K. E., Niem, V. H. (eds), FAO Species Identification Guide for Fishery Purposes. The Living Marine Resources of the Western Central Pacific, vol. 4. B.
Harvey, E. S., M. Cappo, J. J. Butler, N. Hall & G. A. Kendrick, 2007. Bait attraction affects the performance of remote underwater video stations in assessment of demersal fish community structure. Marine Ecology Progress Series 350: 245–254.
Hill, R. D., 1980. Singapore – an Asian City State. Geojournal 4: 5–12.
Igulu, M. M., I. Nagelkerken, M. Dorenbosch, M. G. Grol, A. R. Harborne, I. A. Kimirei & Y. D. Mgaya, 2014. Mangrove habitat use by juvenile reef fish: meta-analysis reveals that tidal regime matters more than biogeographic region. PLoS ONE 9: e114715.
James, G. K., J. O. Adegoke, S. Osagie, S. Ekechukwu, P. Nwilo & J. Akinyede, 2013. Social valuation of mangroves in the Niger delta region of Nigeria. International Journal of Biodiversity Science, Ecosystem Services and Management 9: 311–323.
Lai, S., L. H. Loke, M. J. Hilton, T. J. Bouma & P. A. Todd, 2015. The effects of urbanisation on coastal habitats and the potential for ecological engineering: a Singapore case study. Ocean & Coastal Management 103: 78–85.
Lee, S. Y., J. H. Primavera, F. Dahdouh-Guebas, K. McKee, J. O. Bosire, S. Cannicci & I. Mendelssohn, 2014. Ecological role and services of tropical mangrove ecosystems: a reassessment. Global Ecology and Biogeography 23: 726–743.
Liquete, C., N. Cid, D. Lanzanova, B. Grizzetti & A. Reynaud, 2016. Perspectives on the link between ecosystem services and biodiversity: the assessment of the nursery function. Ecological Indicators 63: 249–257.
Loneragan, N. R., N. A. Adnan, R. M. Connolly & F. J. Manson, 2005. Prawn landings and their relationship with the extent of mangroves and shallow waters in western peninsular Malaysia. Estuarine, Coastal and Shelf Science 63: 187–200.
Manson, F. J., N. R. Loneragan, B. D. Harch, G. A. Skilleter & L. Williams, 2005. A broad-scale analysis of links between coastal fisheries production and mangrove extent: a case-study for northeastern Australia. Fisheries Research 74: 69–85.
Meynecke, J. O., S. Y. Lee & N. C. Duke, 2008. Linking spatial metrics and fish catch reveals the importance of coastal wetland connectivity to inshore fisheries in Queensland, Australia. Biological Conservation 141: 981–996.
Millennium Ecosystem Assessment, 2005. Ecosystems and Human Well-Being: Synthesis. Island Press, Washington, DC.
Monks, N. (ed.), 2006. Brackish-Water Fishes. T.F.H. Publications, Inc, Neptune City.
Murphy, H. M. & G. P. Jenkins, 2010. Observational methods used in marine spatial monitoring of fishes and associated habitats: a review. Marine and Freshwater Research 61: 236–252.
Nagelkerken, I., G. Van der Velde, M. W. Gorissen, G. J. Meijer, T. Van’t Hof & C. Den Hartog, 2000. Importance of mangroves, seagrass beds and the shallow coral reef as a nursery for important coral reef fishes, using a visual census technique. Estuarine, Coastal and Shelf Science 51: 31–44.
Nagelkerken, I., A. M. de Schryver, M. C. Verweij, F. Dahdouh-Guebas, G. Van der Velde & N. Koedam, 2010. Differences in root architecture influence attraction of fishes to mangroves: a field experiment mimicking roots of different length, orientation, and complexity. Journal of Experimental Marine Biology and Ecology 396: 27–34.
Nagelkerken, I., M. Sheaves, R. Baker & R. M. Connolly, 2015. The seascape nursery: a novel spatial approach to identify and manage nurseries for coastal marine fauna. Fish and Fisheries 16: 362–371.
Nam, V. N., S. D. Sasmito, D. Murdiyarso, J. Purbopuspito & R. A. MacKenzie, 2016. Carbon stocks in artificially and naturally regenerated mangrove ecosystems in the Mekong delta. Wetlands Ecology and Management 24: 231–244.
Richards, D. R. & D. A. Friess, 2016. Rates and drivers of mangrove deforestation in Southeast Asia, 2000-2012. Proceedings of the National Academy of Sciences USA 113: 344–349.
Scales, I., D. A. Friess, L. Glass & L. Ravaoarinorotsihoarana, 2017. Rural livelihoods and mangrove degradation in south-west Madagascar: lime production as an emerging threat. Oryx: 1-5.
SeaGIS, 2016. CAL—Stereo Camera Calibration. SeaGIS Pty. www.seagis.com.au.
Sheaves, M., 2005. Nature and consequences of biological connectivity in mangroves systems. Marine Ecology Progress Series 302: 293–305.
Sheaves, M. & B. Molony, 2000. Short-circuit in the mangrove food chain. Marine Ecology Progress Series 199: 97–109.
Sheaves, M., R. Johnston & R. Baker, 2016. Use of mangroves by fish: new insights from in-forest videos. Marine Ecology Progress Series 549: 167–182.
Shinnaka, T., M. Sano, K. Ikejima, P. Tongnunui, M. Horinouchi & H. Kurokura, 2007. Effects of mangrove deforestation on fish assemblage at Pak Phanang Bay, southern Thailand. Fisheries Science 73: 862–870.
TEEB, 2010. Biodiversity, ecosystems and ecosystem services. In Kumar, P. (ed.), The Economics of Ecosystems and Biodiversity: Ecological and Economic Foundation. UNEP/Earthscan, London: 41–112.
Teh, L. C. & U. R. Sumaila, 2013. Contribution of marine fisheries to worldwide employment. Fish and Fisheries 14: 77–88.
Waltham, N. J. & R. M. Connolly, 2007. Artificial waterway design affects fish assemblages in urban estuaries. Journal of Fish Biology 71: 1613–1629.
Wasserman, R. J. & N. A. Strydom, 2011. The importance of estuary head waters as nursery areas for young estuary-and marine-spawned fishes in temperate South Africa. Estuarine, Coastal and Shelf Science 94: 56–67.
Watson, D. L., E. S. Harvey, M. J. Anderson & G. A. Kendrick, 2005. A comparison of temperate reef fish assemblages recorded by three underwater stereo-video techniques. Marine Biology 148: 415–425.
Weinstein, M. P., S. Y. Litvin & V. G. Guida, 2005. Considerations of habitat linkages, estuarine landscapes, and the trophic spectrum in wetland restoration design. Journal of Coastal Research SI(40): 51–63.
Weis, J. W. & P. Weis, 2005. Use of intertidal mangrove and sea wall habitats by coral reef fishes in the Wakatobi Marine Park, Indonesia. The Raffles Bulletin of Zoology 53: 119–124.
Whitfield, A. K., 2017. The role of seagrass meadows, mangrove forests, salt marshes and reed beds as nursery areas and food sources for fishes in estuaries. Reviews in Fish Biology and Fisheries: 1–36.
Whitelaw, A. W., K. J. Sainsbury, G. J. Dews & R. A. Campbell, 1991. Catching characteristics of four fish-trap types on the North West Shelf of Australia. Marine and Freshwater Research 42: 369–382.
Wraith, J., T. Lynch, T. E. Minchinton, A. Broad & A. R. Davis, 2013. Bait type affects fish assemblages and feeding guilds observed at baited remote underwater video stations. Marine Ecology Progress Series 477: 189–199.
Yee, A. T. K., W. F. Ang, S. Teo, S. C. Liew & H. T. W. Tan, 2010. The present extent of mangrove forests in Singapore. Nature in Singapore 3: 139–145.
Acknowledgements
RB was supported by the Fulbright Program, United States Government. DAF acknowledges support from the Singapore Ministry of Education Academic Research Fund Tier 1, Government of Singapore (R-109-000-166-112) and NH acknowledges internal funding from JCU (CRT/2016/04). All research was conducted with approval from IACUC (B14-0207). Research was conducted with permission of the National Parks Board, Government of Singapore, under the research permit NP/RP936. We acknowledge advice and support from Leon Gaw, Lee Wei Kit, Ong Wei Bin, Zheng Zhangxin (National University of Singapore) and Phillip Lim.
Author information
Authors and Affiliations
Corresponding author
Additional information
Guest editors: K. W. Krauss, I. C. Feller, D. A. Friess, R. R. Lewis III / Causes and Consequences of Mangrove Ecosystem Responses to an Ever-Changing Climate
Rights and permissions
About this article

Cite this article
Benzeev, R., Hutchinson, N. & Friess, D.A. Quantifying fisheries ecosystem services of mangroves and tropical artificial urban shorelines. Hydrobiologia 803, 225–237 (2017). https://doi.org/10.1007/s10750-017-3299-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-017-3299-8