
Abstract
We examined the recovery of a fish assemblage following the catastrophic spill of highly alkaline (pH ~13) red sludge into a lowland river (i) to characterize taxonomic- and trait-based colonization of fishes in the river in the first 3 years of the recovery and (ii) to determine which structural- and trait-based variables best predicted colonization. Species richness showed comparable values to pre-disturbance state <1 year after the spill. We found only moderate changes in the dominance of the most abundant species between pre- and post-disturbance periods, and consistent changes in the relative abundance of some rare species during the post-disturbance period. Frequency of occurrence (%) of the fishes in the watershed and their relative abundance (%) proved to be the most important predictor variables in colonization, whereas trait-based variables had a less important role. Our study about one of the largest scale and most serious documented fish kill shows that both taxonomic- and trait-based structure of fish assemblages can regenerate remarkably fast in a modified river and also shows that unfortunate chemical spills provide insights into the assembly of stream fish assemblages.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Most of our knowledge about the organization of ecological assemblages comes from the examination of colonization dynamics after natural- or human-mediated disturbances (Nyström et al., 2000; Turner, 2010). For example, large-scale infrequent disturbances LIDs (see Turner & Dale, 1998) such as the 1980 eruption of the volcano Mount St. Helens (del Moral & Bliss, 1993) or the 1998 fires in the Yellowstone National Park, USA (Romme et al., 2011) provided natural experiments which gave insight into successional dynamics and assembly rules. Disturbance-mediated colonization dynamics in a network of habitat patches form the basic principle of metacommunity ecology, which proved to be one of the most influential approaches in understanding regional scale diversity and dynamics of assemblages (Leibold et al., 2004).
In riverine ecosystems, unpredictable floods and droughts provide the most characteristic examples of LIDs (Resh et al., 1988; Lake, 2000). Several studies showed the remarkable resilience of stream organisms to natural, flow-generated disturbances (Grimm & Fisher, 1989; Niemi et al., 1990; Stanley et al., 2010). Flow-related disturbances, however, are only rarely serious enough to extirpate the whole fauna. In flow-related disturbances, recovery happens both from instream refugium areas and from other nearby stream reaches. Less is known about how human-mediated LIDs, such as chemical pollution events, influence the recovery and colonization dynamics of aquatic assemblages. Chemical spills can be so extreme that the whole-stream fauna is extirpated. In these cases, recovery can be purely a primary successional process, where within habitat population size contributes only little or negligibly to recovery rates and, therefore, colonization happens predominantly from other habitats. How stream assemblages form from the regional species pool during primary succession is less known, and most of our knowledge comes from glacier streams (Milner et al., 2011). Further, although there are some well-documented case studies, which report on the recolonization of the aquatic fauna following large-scale pollution events, most of these studies simply followed changes and recovery in assemblage structure using taxonomic datasets (Meade, 2004; Kubach et al., 2011).
Trait-based analyses characterize species based on their biological features. Complementing taxonomic approaches with trait-based ones may help to better understand the factors associated with colonization and recovery after LIDs, and would promote better comparability with studies from different biogeographic regions. In fact, there are indications that interspecific variations in a variety of traits can influence colonization and recovery rates (Detenbeck et al., 1992; Vaughn, 2012). However, a more detailed understanding is necessary of how the interplay of taxonomic- (e.g., species composition and abundance) and trait-based assemblage properties influences colonization dynamics following LIDs. In a recent experimental defaunation study with stream fishes, it was found that both abundance of fishes in nearby (source) habitats and the differences in their mobility significantly influence recovery rates (Albanese et al., 2009). However, it remains a question of how the lessons of relatively small-scale experimental studies (i.e., <500-m-long reaches) can be extrapolated for whole streams or rivers. Overall, understanding colonization processes following large-scale pollution can further our basic knowledge of assemblage organization in streams and can help environmental managers to define restoration programs to aid large-scale recovery of the biota to pre-disturbance state.
As a result of a tragic industrial accident, highly alkaline red mud (pH ~13) of about 1 million m3 escaped from the depository of a Hungarian alumina factory on 4 October 2010 (Schiermeier & Balling, 2010; Gelencsér et al., 2011). The sludge flooded three settlements and ~40 km2 agricultural area along the Torna stream and the Marcal River (Figs. 1, 2), causing the death of 10 persons and seriously injuring more than 150 people. Water management was effective in neutralizing pH to levels below 10.5 soon after the spill by adding acid and gypsum to the water (Renforth et al., 2012). Yet, the large-scale pulse disturbance completely extirpated the fauna of the Torna stream and caused an almost complete extirpation in the affected 70.7-km-long section of the Marcal River (Vass et al., 2013). Fish kills and injured fish were observed even in the Rába River (the recipient river of the Marcal) and in the Danube River, respectively.

Color of the Marcal River at the S2 site 2 weeks after the red mud disaster

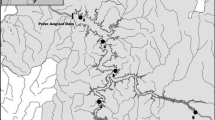
Map of the Marcal River system and the distribution of sampling sites on the river (S1–S6), and on its tributaries (open circles). Note that the symbols of some sites which are situated at the mouth of the tributary streams are covered by the S1–S6 symbols. Sites which were affected by the accident are indicated with red symbols. Note also that although some sampling sites were selected on very small creeks and contained fish, these waterflows are not indicated on the map. Inflow of the Torna stream which drained the highly alkaline red mud (pH ~13) into the river is indicated with a red arrow. The smaller map of Hungary indicates the route of the Danube River and the position of Budapest, the capital of Hungary
The purpose of this study was (i) to characterize the colonization of fishes of the Marcal River in the first three years from the start of the disaster and (ii) to determine which structural- and trait-based variables best predicted the recolonization of fishes. Since the Marcal is a recipient river of several smaller (i.e., 2–3 order) streams, we predicted fast recolonization congruent with the findings in the literature (Detenbeck et al., 1992; Meade, 2004; Kubach et al., 2011). However, given this was a large-scale disturbance, we were especially interested (i) to examine how recolonization is related to the spatial position of the sampling sites along the river and (ii) how structural- and trait-based variables may influence the proliferation of colonization in time.
Materials and methods
Study area
The Marcal River is a typical lowland river situated in West Hungary (Fig. 2). It has a length of 100.4 km, an average slope of 0.24 m km−1, and a total drainage area of 3084 km2. The river is slightly alkaline (pH 8.1–8.3) and has a conductivity of 850–1140 μS cm−1. The riverbed is covered by a variety of substrate types including clay, sandy-clay, silty-sand, and a smaller portion of gravel. In the vegetation period, the river is densely covered by both emergent (e.g., Phragmites australis, Typha spp.) and submerged (e.g., Nuphar lutea, Polygonum amphibium, Potamogeton spp.) macrophytes. The river is channelized, which allows only little meandering. Therefore, reach-level habitat heterogeneity is restricted mainly to variations in the macrovegetation coverage. Although the river is modified by channelization, it had a relatively species-rich fauna, due to the permeability of the river and its tributary streams (i.e., no major dams or culverts influence movement), and instream vegetation cover, which provides habitat heterogeneity to some degree.
Sampling design and fish surveys
For the purpose of this study, six river sections (S1–S6) were selected along the course of the river. Site 1 was located 7.5 km upstream of the spill origin (i.e., mouth of the Torna stream). Sites S2, S3, S4, S5, and S6 were located 6.6, 19.0, 43.8, 52.5, and 67.0 km downstream of the spill, respectively, and all were affected by the disturbance event (Fig. 2). In the examined study sites, mean wetted channel width, mean current velocity, and mean water depth varied downstream between 3.2 and 18.0 m, 3.0 and 31.8 cm s−1, and 0.3 and 1.4 m, respectively.
Samples were collected during spring (end of April or early May), summer (July or early August), and autumn (October), starting 2 weeks after the spill on 19 October 2010 and ending on 7 October 2013, and this has yielded a total of 10 surveys. We used either wading or boat electrofishing for collecting fish. Since S1 was wadeable and was covered with heavy reed vegetation, we used upstream wading, single-pass electrofishing for collecting fish. Here, a 200-m-long reach was sampled using a backpack electrofishing gear (Hans-Grassl IG200/2B, PDC, 75–100 Hz, 350–650 V, max. 10 kW; Hans Grassl GmbH, Germany), wherein sampling effort was found to yield representative samples for characterizing the reach-level composition of fish assemblages in wadeable streams (for further details see Erős, 2007; Sály et al., 2009). The other sites (S2–S6) were sampled from a boat using the same electrofishing device and its accessories as was used for S1 site. To allow effective maneuvering in this shallow and vegetated river, a small rubber boat (Yamaha 300S) was used. The crew comprised two persons: one for catching the fish with the hand-held anode (2.5-m-long pole with a net of 40-cm diameter, mesh size 6 mm) and one for driving the boat. Continuous electrofishing was carried out for a 400-m-long reach per site, by dipping the anode into the water at approximately 2-m-long intervals and pulling the anode toward the boat, while moving slowly ahead with the flow and controlled by an oar if it was necessary. Caught fish were placed into a 100-L water-filled tank in the boat and kept alive until the end of the fishing. Fish were then identified, counted, and measured for standard length (SL) to the nearest mm and released back to the river.
Although the Marcal was an unmonitored river before the pollution, data of a former fish assemblage survey (Harka et al., 2009) could be used as a pre-disturbance dataset for evaluating “before-after changes” in fish assemblages. In the frame of this survey, seven sites in the Marcal River and two sites in its tributaries were sampled using both electrofishing and seine nets in the summer of 2008. Of these, four sites were close (i.e., within 3 km) to our sampling sites and we used these data as reference to get some insight into pre-disturbance state for S1 (no disturbance), S2, S3, S6 sites. Note that S1 cannot be used as a reference site for comparison with the polluted sites because of its different environmental and topographical characteristics (i.e., shallow, wadeable upstream reach). However, data from this site can be used to get insight into the potential source of colonizing species.
Finally, to quantify relative abundance and frequency of occurrence of each species at the catchment level, we used our field surveys and literature sources (Harka & Szepesi, 2011), which accumulated about the fish assemblages of the tributary streams of the Marcal and its recipient watercourse (mouth of the Rába River; Fig. 2). All of the tributary streams of the Marcal are wadeable, relatively small streams (i.e., <5 m wide) with similar environmental characteristics to S1 site. The surveys were carried out in almost all tributaries of the river using both electrofishing and seine netting in a variety of seasons (spring, summer, autumn) between 2008 and 2013. The accomplished dataset contains 149 records (survey of a site in a single occasion) of 93 sites, representing 42 species and 31,259 individuals. We believe it provides a representative sample of fish at the water basin scale (Table 1).
Species traits
We considered altogether three main trait groups that can be highly influential in colonization dynamics and post-disturbance trait-based organization in fishes and other organism groups (Detenbeck et al., 1992; Hitt & Roberts, 2012; Vaughn, 2012). These trait groups were (1) life-history strategy, (2) coarse-scale vertical habitat use (i.e., position in the water column), and (3) feeding (Table 1). Life-history strategy was characterized using the system of Winemiller & Rose (1992) and Winemiller (1992). In this system, periodic strategist species are large bodied with late maturation, high fecundity, low juvenile survivorship, and with no developed parental care. They typically occupy seasonal, periodically suitable environments and since they are large bodied they are capable of medium- or long-distance migration (101–102 km) within short time periods (weeks or months). Opportunistic strategist species are small bodied with early maturation, low fecundity, and low juvenile survivorship, and that typically inhabit unpredictable environments. Equilibrium strategist species are small- to medium-bodied fishes with intermediate maturation age, low fecundity, high juvenile survivorship with developed parental care, and that typically inhabit constant environments. We used our former works to classify species to these main life-history groups and to intermediate classes (Erős, 2005; Erős et al., 2012b). We were also interested to test how position in the water column (i.e., benthic vs. non-benthic) influences colonization processes since this trait can strongly interact with both body size and trophic position (i.e., feeding) in shaping the organization of the assemblage following disturbance. Finally, feeding is an especially important trait in both colonization and post-disturbance niche-based interactions (Mackay, 1992; Czeglédi & Erős, 2013). We therefore also categorized species based on their feeding traits (e.g., herbivore, omnivore, invertivore, and piscivore see Table 1 for details) similar to other trait-based studies (Frimpong & Angermeier, 2010; Heino et al., 2013). For specific hypotheses and a summary about the role of individual traits in the colonization process, see the “Discussion” section.
Data analysis
To get basic inferences on the process of recolonization, we examined changes in species richness and rank abundance of species comparing pre- and post-disturbance periods. For species richness measures, we used the actual number of species per site, the total number of species collected per sampling occasion, and the total cumulated number of species in time (i.e., pooled over sampling occasions see Fig. 3).

Changes in the values of species richness in the Marcal river before and after the red mud accident. Dashed line indicates the date of the spill (October 2010). Srtotal shows the total number of species collected on a given sampling occasion (e.g., on spring 2001), and Srcum shows the total cumulated number of species in time (i.e., pooled over sampling occasions). Note that S1 was unaffected by the spill
The examination of changes in species rank abundances is an often used, easy to interpret analysis in succession research (Harpole & Tilman, 2006; Collins et al., 2008). To examine robust changes, we depicted the relative rank abundance of species by pooling seasonal data for each post-disturbance year separately (i.e., 2011, 2012, 2013), and compared changes in the rank order of species with the results of the pre-disturbance 2008 survey.
Beside these basic metrics, we used principal component analyses (PCA) to assess changes in both taxonomic- and trait-based assemblage structures following the disaster. We used Hellinger-transformed abundance data (Legendre & Gallagher, 2001) to characterize taxonomic structure in the ordination. Extremely rare species (i.e., species with an overall relative abundance <0.1%) were deleted from the analysis. For the examination of trait-based changes, a trait abundance matrix was used. This was obtained by multiplying the species abundance matrix with the dummy (i.e., 0/1) coded species trait matrix. The trait abundance matrix was also standardized so that the summed proportional abundance values of the traits within each trait group were equal to 100%. This standardization gave equal weight for each trait group (life history, position in water column, feeding) and yielded proportional weight for each trait within the trait group (%). Similar to the taxonomic data table, the trait abundance data were Hellinger transformed prior to the analysis.
Predicting colonization with structural- and trait-based attributes
We used a predictive modeling framework to select those variables, both structural and trait based that may provide the most plausible explanation for the colonization dynamics and recovery of this riverine fish assemblage. For structural predictor variables, we used the relative abundance and frequency of occurrence of each fish species at the water basin level (Table 1). In this way, we considered the whole species pool as potential source of colonist species in the successional process of recovery. We hypothesized that both variables can be influential, since colonization may depend both on the abundance of species and the number of source patches from which colonization can happen (Meffe & Sheldon, 1990; Adams & Warren, 2005; Albanese et al., 2009). The trait-based attributes (i.e., life history, vertical habitat use, feeding; see above) of the species were used as another set of predictor variables. For response variable (i.e., as an indicator of colonization and recovery), we used the relative abundance of colonist species in the Marcal River averaged over seasons in each of the three years separately.
We used univariate regression TREEs to model the importance of structural- and trait-based variables in the process of colonization. Regression TREEs are highly suitable for analysis of complex ecological data with nonlinear relationships (De’ath & Fabricius, 2000). TREE models recursively split the observations (mean relative abundance of fish species in our case) into two separate groups by values of the descriptor variables in a way that the heterogeneity of the response variable (measured e.g., by sum of squares) should be maximized between the groups. After model fitting, the full-grown (i.e., overfitted) TREE models were pruned back according to the so-called “1-SE rule” (for details see Zuur et al., 2007). We characterized the overall model performance on the basis of the R 2 value and a sevenfold cross validation error value. Importance of the descriptor variables was evaluated by comparing the sum of the gain in the R 2, attributed to each variable at each split in the model. Again, we ran three models for each year separately to examine possible changes in the importance of the variables during different phases of colonization. All statistical analyses were performed in the freely available software package R (R Core Team, 2013), using the packages vegan (Oksanen et al., 2013), part (Therneau et al., 2013), and Caret (Kuhn, 2014).
Results
We collected altogether 35 species and 13,177 individuals of fish in the Marcal River during the ten surveys from October 2010 to October 2013 (Fig. 3). Twenty-two species were found in the river during the 2008 survey, before the pollution event. However, 2 weeks after the spill only four species, the bleak (Alburnus alburnus), the roach (Rutilus rutilus), the bitterling (Rhodeus sericeus), and the gibel (Carassius gibelio), were observed in the disturbed section (S2–S6 sites), and seven species from the most upstream unaffected site (S1). The appearance of new species in the river was fast. One year after the disturbance, in autumn 2011, the total cumulated number of species detected in the river reached 31, and it increased only very slightly during the two further years to 35 in autumn 2013. This increase in species richness was clearly due to the colonization of new species at the disturbed sites, since species richness remained relatively stable at the unaffected S1 site. In sum, although there was some spatial (i.e., between site) and temporal (i.e., between occasion) variability in the detected number of species, species richness stabilized at a higher level 9 months post-disturbance than it was found by the reference 2008 survey. Overall, most species (35 of the 42 species, 83.3%) which were found in the watershed’s streams appeared in the affected S2–S6 sites of the Marcal during the three year survey. Only three rare tributary species (i.e., crucian carp Carassius carassius, three-spined stickleback Gasterosteus aculeatus, and European minnow Phoxinus phoxinus) and four species from the recipient Rába River (spirlin Alburnoides bipunctatus, white-eye bream Abramis sapa, racer goby Babka gymnotrachelus, and bighead goby Ponticola kessleri) remained undetected compared with the reference dataset (Table 1).
Similar to species richness, a sharp increase in abundance (CPUE data) values could be observed at the disturbed sites (Fig. 4). Only 59 specimens were collected after the spill, but in spring 2011, 6 months after the disaster, more than sixfold increase in abundance was found. Overall, total abundance of fish generally increased during the first year of the study then it started to fluctuate. Changes in abundance were mainly driven by the three most abundant species (bleak, roach, and bitterling), while the contribution of other species to total abundance remained relatively low. Although the relative abundance of the dominant species did not change much at the affected section compared with the reference 2008 survey, when these three species comprised 66% of the assemblage, moderate changes in the rank abundance of species could be observed (Fig. 5). For example, roach (27.5%) and bitterling (34.6%) were the most dominant species in 2008, similar to the post-disturbance period, whereas bleak, which proved to be the most dominant fish after the spill, was only the fifth most dominant species (3.9%) during the 2008 survey. The monkey goby (Neogobius fluviatilis) and the dace (Leuciscus leuciscus) were the third (8.8%) and the fourth (6.4%) most dominant species in 2008, respectively, but their contribution was relatively low in the post-disturbance phase (e.g., 0.7% for both species in 2013). Consistent temporal changes in the relative abundance of rare species included the increasing dominance of chub (Squalius cephalus), perch (Perca fluviatilis), round goby (Neogobius melanostomus), and pike (Esox lucius) in the catches during the post-disturbance years (Fig. 5). The abundance of the most invasive species (black bullhead Ameiurus melas, gibel, stone morocco Pseudorasbora parva, Neogobius spp.) proved to be low during the 3 years.

Changes in the values of catch per unit effort abundance data for the polluted river section (i.e., catches for S2–S6 sites are combined). Different shadings show the contribution of the three most abundant species (bleak, albalab; bitterling, rhoser; roach, rutrut) and those of other less abundant species (Other spp.) to overall abundance. Numbers show the number of species for each survey

Rank abundance curves of pooled seasonal data for each year. Species are ordered according to their rank order in the pre-disturbance 2008 survey. Note that only the most abundant 20 species are shown for brevity. For species name abbreviations, see Table 1
The first two axes of the species abundance-based PCA indicated high temporal and spatial variability among the sampling sites (Fig. 6a, b). Pre-disturbance samples (i.e., 2008 survey) showed some separation from post-disturbance ones along the combination of the first two PCA axes, reflecting some differences in the relative abundance of species between the two periods (Fig. 6a), which corresponded with the results of the rank abundance curves (i.e., relatively high abundance of roach, bitterling, monkey goby, and some rare species in the pre-disturbance period). However, samples from post-disturbance years did not clearly separate and showed high temporal variability and largely overlapping assemblage structure at the disturbed sites (S2–S6) (Fig. 6b). Roach and belica (Leucaspius delineatus), two characteristic species of lowland streams, were more abundant at the most upstream and unaffected S1 site (for roach negative correlation with axis 1 and positive correlation with axis 2; for belica negative correlation with axis 1), while the relative abundance of some riverine cyprinids (bleak, gibel, ide Leuciscus idus, common bream Abramis brama) increased at the more downstream S2–S6 sites (positive correlation with axis 1). The temporal trajectories did not indicate strong directional changes in assemblage composition at either site (Fig. 6b).

The principal component ordination (PCA) of fish assemblage samples based on relative abundance data, where symbols differentiate samples collected in different years (a), and subfigures show the temporal trajectories of the sampling sites (S1–S6) in the ordination plane (b). Variables which showed significant (P < 0.5) Spearman rank correlation values with PC1 and PC2 axes are also indicated. For species name abbreviations, see Table 1
The first two axes of the trait abundance-based PCA also showed high variability among samples both between years and sites (Fig. 7a, b), similar to the result obtained for taxonomic structure. Site 1 separated the most from the other sites, especially along PC1, and could be characterized by the relative dominance of the life-history trait “periodic” and the feeding trait “omnivore” (positive correlation with axis 1). Both traits are the characteristic ones of roach, which was the most dominant fish at this site in all occasions.

The principal component ordination (PCA) of fish assemblage samples based on relative trait abundance data, where symbols differentiate samples collected in different years (a), and subfigures show the temporal trajectories of the sampling sites (S1–S6) in the ordination plane (b). Variables which showed significant (P < 0.5) Spearman rank correlation values with PC1 and PC2 axes are also indicated. For species name abbreviations, see Table 1
The regression TREE models indicated that a moderate amount of variance (47.9–52.5%) in colonization dynamics (i.e., mean relative abundance of fishes in the affected section in a given year) could be explained by some simple structural- and trait-based variables (Table 2). The models indicated subtle increase in the predictability of the assemblage from year to year, although the order of importance of the variables remained the same. Frequency of occurrence (%) of the fishes in the watershed and their relative abundance (%) proved to be the most important variables, which individually could explain virtually the total variability in the dataset. Trait-based variables had a less important role in the colonization process (Table 2).
Discussion
In this study, we reported on one of the largest scale and most serious documented fish kill following a pulse disturbance event. Species richness showed comparable values to pre-disturbance state <1 year after the spill. However, spatial and temporal variations in assemblage structure were high and did not depend on longitudinal position of the sampling sites in the polluted section. Percent of sites occupied in the water basin and relative abundance of fishes proved to be the most important predictors of species colonization (~50%), whereas trait-based variables had a less important role (<15%). This study thus shows that both taxonomic- and trait-based structure of fish assemblages can regenerate remarkably fast after chemical spills even at the whole river scale (here 70.7-km-long affected section), if there is possibility for colonization from the tributaries and from upstream and downstream source areas.
In riverine systems, the structure of the dendritic stream network largely determines dispersal possibilities, contrary to terrestrial landscapes, where colonization can happen from many directions (Labonne et al., 2008; Erős et al., 2012a). The topology of the watershed, specifically the unaffected upstream headwater section and the many right-side tributaries, with relatively similar size and even distribution, could ensure equal colonization probabilities along the longitudinal section of the Marcal River from source to mouth (see Fig. 2). Comparing species composition in the watershed and in the Marcal, it is likely that colonization of the river happened from all possible sources of direction. Since the tributaries have very similar species composition to that of the upstream (unaffected) section of the river, we cannot separate their relative roles in colonization. The most dominant immigrants were the most abundant tributary species (e.g., roach, bitterling, chub). Nevertheless, the appearance of some riverine species in the Marcal (e.g., barbel Barbus barbus, ide, round goby, vimba Vimba vimba) also suggests some role of upstream migration from the recipient Rába River. Although bleak, a common shoal forming water column cyprinid, occupies the tributary mouth sections, it could also colonize the river from downstream source areas. Consequently, colonization from downstream source areas proved to be also important, and given the permeability of the river, fish could move freely even to the most upstream parts of the affected section. We believe the extreme fast increment in species richness at this large spatial scale (i.e., from seven species to 31 species within one year, and a total of 35 species during three years for this 100.5-km-long river) adds important findings to the literature on the recovery of fishes following disturbance events. In fact, there are no other studies we could effectively compare our results with, because former events happened at smaller spatial scales and the studies did not address trait-based changes (22.3 km in Meade, 2004 and 37 km in Kubach et al., 2011, which are the longest sections found in the literature).
In their review on the recovery of temperate stream fishes from disturbance, Detenbeck et al. (1992) concluded that species composition, richness, and total density of fish recovered within one year over 70% of the systems were studied, although they did not address natural variability in assemblages in reference samples. In the most comparable situation to the LID we examined, Kubach et al. (2011) found the strongest similarity between disturbed and reference group samples 52 months post-disturbance after an oil spill in the Reedy River, USA, but the assemblage did not show clear stability on forthcoming sampling event. We found clear successional processes of species recolonization and fast recovery in the first year. However, the ordination analyses indicated variability in taxonomic- and trait-based structure in the next 2 years of the study without clear stabilization. These changes can be mainly related to changes in the relative abundance of species among sites and sampling occasions since species richness relatively stabilized one year post-disturbance. Our large-scale study thus fully supports the predictions of small-scale, experimental defaunation studies, which suggested fast recolonization, but temporal variability in assemblage structure after pulse disturbances in low-gradient streams with low habitat complexity and permeability to movement (Peterson & Bayley, 1993; Albanese et al., 2009).
Disentangling the relative importance of structural- and trait-based variables in recovery after LIDs is a largely unknown but one of the most interesting tasks in ecology with strong applied aspects (Turner & Dale, 1998; Albanese et al., 2009). Frequency of occurrence (%) and relative abundance of the fishes (%) in the water basin proved to be equally important variables in determining colonization, indicating the critical importance of the distribution of fishes in the surrounding riverscape. The number of occupied patches is critical in colonizing new areas for metapopulations (Gotelli & Taylor, 1999), while abundant species can supply more colonists for recovery (Albanese et al., 2009). A colonization where the most frequent and/or abundant species are the most important for recovery can be considered as a neutral (random) process. Alternatively, abundance and frequency may also indicate the importance of niche-based processes if the rarity of species correlates strongly with their environmental requirements. Unfortunately, we cannot really disentangle the role of neutral versus niche-based processes in colonization, because the most dominant species of the Marcal River and the tributaries were the same independent on pre- or post-disturbance state, and such a pattern can support both hypotheses. In sum largely the same assemblage recovered in the Marcal River as was found formerly in the river itself or as can be found overall in the watershed’s streams.
Biological traits have been used successfully in predicting human impacts (Olden et al., 2006; Dolédec et al., 2011), but their role in large-scale colonization dynamics is much less known (but see e.g., Mackay, 1992). We considered key trait variables in this study. Life-history strategy, which incorporates information on size, generation time, juvenile survivorship, and fecundity, may be very important for understanding the role of extinction–colonization dynamics and environmental filtering from the species pool following LIDs. For example, it can be predicted that periodic strategist species are potentially the first colonizers following LIDs due to their high migration potential. Opportunistic species are probably worse colonizers, due to their small size, but since they live in unpredictable environment they may colonize disturbed areas successfully. If body size is an important predictor of colonization potential equilibrium species can be either bad or relatively good colonizers. However, they generally prefer constant environment, and therefore disturbance events may halt their successful colonization on short time scales. Similar predictions can be also addressed for feeding traits in their role in the process of colonization. For example, it can be predicted that omnivore species are more successful colonizers than strictly insectivore species after LIDs, since they rely on a variety of food resource, while the establishment of insectivores depends more on the speed of the colonization of the macroinvertebrate fauna. Following this argument, piscivore species can be the last colonizers, since their permanent establishment requires stable populations of small-bodied fishes.
Our TREE models suggested some role of traits in predicting fish colonization, but their role was rather minor. We anticipate this finding to the extreme fast colonization process. Even seemingly bad disperser species repopulated the polluted river section fast. For example, bitterling is a small, rather sedentary shoaling fish (especially juveniles), but the second most abundant in the watershed’s streams. Its 30–40-mm-long individuals were found en masse even in the first spring along the affected rivers section (i.e., S2–S6 sites), probably due to either passive or active drifting from the tributaries by the autumn 2010 and spring 2011 floods (see e.g., Jurajda, 1998). Therefore, a fish assemblage with relatively diverse trait structure could colonize the river successfully from the tributaries even in the first year, and relatively minor changes happened in the rank abundance distribution of the dominant species in the following two years. Example of trait-based changes for the less abundant species includes the increasing dominance of piscivore fish in the catches (e.g., pike). Naturally, a stable prey population is a prerequisite for the proliferation of piscivores, and the increasing abundance of small cyprinids (e.g., roach, bitterling) provided an ideal food resource for stabilizing their populations. Although we did not experience a completely extirpated community, the number of species and the number of individuals were so extremely low in our first sampling occasion that we can consider the recovery of the polluted section of the river as a primary successional process. Note that it is hardly improbable that any fish could survive the extremely alkaline water, but autumn floods could continuously wash down fish (especially the young-of-the-year fish) into the river from the tributaries, which could survive there soon after the pollution effect mitigated (within some days). In this regard, an important finding of this study is that species entered the system in their proportional ratio of the water basin’s streams. Invasive species could not proliferate, although there are some highly invasive species in the watershed which could be fast and successful colonizers due to their early maturation, relatively short life cycle and reproduction potential (black bullhead, gibel, stone morocco, goby species). However, the most abundant native species show similar trait characteristics. Therefore, the often reported rule in invasion biology that disturbances can promote invasion (Hobbs & Huenekke, 1992) could not really fit to the successional dynamics of fishes in the Marcal River.
In conclusion, our results about the fast colonization and recovery of fish assemblages in the Marcal River after the catastrophic red mud accident are similar to the values found for smaller scale disturbances, when shorter stream sections were affected by chemical pollution (for review see Detenbeck et al., 1992; Kubach et al., 2011). Our study highlights the importance of both the topological structure and the permeability of the stream network, which can enhance recovery from pollution events, and shows that LIDs by unfortunate chemical spills provide insights into the taxonomic- and trait-based assembly of stream fish assemblages.
References
Adams, S. B. & M. L. Warren, 2005. Recolonization by warmwater fishes and crayfishes after severe drought in upper Coastal Plain hill streams. Transactions of the American fisheries Society 134: 1173–1192.
Albanese, B., P. Angermeier & J. T. Peterson, 2009. Does mobility explain variation in colonization and population recovery among stream fishes? Freshwater Biology 54: 1444–1460.
Collins, S. L., K. N. Suding, E. E. Cleland, M. Batty, S. C. Pennings, K. L. Gross, J. B. Grace, L. Gough, J. E. Fargione & C. M. Clark, 2008. Rank clocks and plant community dynamics. Ecology 89: 3534–3541.
Czeglédi, I. & T. Erős, 2013. Characterizing congruency in the long-term taxonomic and functional variability of a stream fish assemblage. Fundamental and Applied Limnology 183: 153–162.
De’ath, G. & K. E. Fabricius, 2000. Classification and regression trees: a powerful yet simple technique for the analysis of complex ecological data. Ecology 81: 3178–3192.
del Moral, R. & L. C. Bliss, 1993. Mechanism of primary succession: insights resulting from the eruption of the Mount St Helens. Advances in Ecological Research 24: 1–66.
Detenbeck, N. E., P. W. DeVore, G. J. Niemi & A. Lima, 1992. Recovery of temperate-stream fish communities from disturbance: a review of case studies and synthesis of theory. Environmental Management 16: 33–53.
Dolédec, S., N. Phillips & C. Townsend, 2011. Invertebrate community responses to land use at a broad spatial scale: trait and taxonomic measures compared in New Zealand rivers. Freshwater Biology 56: 1670–1688.
Erős, T., 2005. Life history diversification in the Middle Danubian fish fauna – a conservation perspective. Archiv für Hydrobiologie, Supplementband 158, Large Rivers 16: 289–304.
Erős, T., 2007. Partitioning the diversity of riverine fish: the roles of habitat types and non-native species. Freshwater Biology 52: 1400–1415.
Erős, T., J. D. Olden, R. S. Schick, D. Schmera & M. J. Fortin, 2012a. Characterizing connectivity relationships in freshwaters using patch-based graphs. Landscape Ecology 27: 303–317.
Erős, T., P. Sály, P. Takács, A. Specziár & P. Bíró, 2012b. Temporal variability in the spatial and environmental determinants of functional metacommunity organization – stream fish in a human-modified landscape. Freshwater Biology 57: 1914–1928.
Frimpong, E. A. & P. L. Angermeier, 2010. Trait-based approaches in the analysis of stream fish communities. American Fisheries Society Symposium 73: 109–136.
Gelencsér, A., N. Kováts, B. Túróczi, Á. Rostási, A. Hoffer, K. Imre, I. Nyírő-Kósa, D. Csákberényi-Malasics, Á. Tóth, A. Czitrovszky, A. Nagy, S. Z. Nagy, A. Ács, A. Kovács, Á. Ferincz, Z. Hartyáni & M. Pósfai, 2011. The red mud accident in Ajka (Hungary): characterization and potential health effects of fugitive dust. Environmental Science and Technology 45: 1608–1615.
Gotelli, N. J. & C. M. Taylor, 1999. Testing metapopulation models with stream-fish assemblages. Evolutionary Ecology Research 1: 835–845.
Grimm, N. B. & S. G. Fisher, 1989. Stability of periphyton and macroinvertebrates to disturbance by flash floods in a desert stream. Journal of the North American Benthological Society 8: 293–307.
Harka, Á., Z. Szepesi & L. Nagy, 2009. Faunal survey on the fish community of Marcal River. Pisces Hungarici 3: 27–32. (in Hungarian with a summary in English).
Harka, Á. & Z. Szepesi, 2011. Investigation on the fish fauna on the side streams of the Marcal River. Pisces Hungarici 5: 99–110. (in Hungarian with a summary in English).
Harpole, W. S. & D. Tilman, 2006. Non-neutral patterns of species abundance in grassland communities. Ecology Letters 9: 15–23.
Heino, J., D. Schmera & T. Erős, 2013. A macroecological perspective of trait patterns in stream communities. Freshwater Biology 58: 1539–1555.
Hitt, N. P. & J. H. Roberts, 2012. Hierarchical spatial structure of stream fish colonization and extinction. Oikos 121: 127–137.
Hobbs, R. J. & L. F. Huenekke, 1992. Disturbance, diversity and invasion: implications for conservation. Conservation Biology 6: 324–337.
Jurajda, P., 1998. Drift of larval and juvenile fishes, especially Rhodeus sericeus and Rutilus rutilus, in the River Morava (Danube basin). Archiv für Hydrobiologie 141: 231–241.
Kubach, K. M., M. C. Scott & J. S. Bulak, 2011. Recovery of a temperate riverine fish assemblage from a major diesel oil spill. Freshwater Biology 56: 503–518.
Kuhn, M., 2014. Caret: Classification and Regression Training. R package version 6.0-21. http://CRAN.R-project.org/package=caret. Available on 21 Jan 2014.
Labonne, J., V. Ravigné, B. Parisi & C. Gaucherel, 2008. Linking dendritic network structures to population demogenetics: the downside of connectivity. Oikos 117: 1479–1490.
Lake, P. S., 2000. Disturbance, patchiness and diversity in streams. Journal of the North American Benthological Society 19: 573–592.
Legendre, P. & D. Gallagher, 2001. Ecologically meaningful transformations for ordination of species data. Oecologia 129: 271–280.
Leibold, M. A., M. Holyoak, N. Mouquet, P. Amarasekare, J. M. Chase, M. F. Hoopes, R. D. Holt, J. B. Shurin, R. Law, D. Tilman, M. Loreau & A. Gonzalez, 2004. The metacommunity concept: a framework for multi-scale community ecology. Ecology Letters 7: 601–613.
Mackay, R. J., 1992. Colonization by lotic macroinvertebrates: a review of processes and patterns. Canadian Journal of Fisheries and Aquatic Sciences 49: 617–628.
Meade, R., 2004. Fish and invertebrate recolonization in a Missouri prairie stream after and acute pollution event. North American Journal of Fisheries Management 24: 7–19.
Meffe, G. K. & A. L. Sheldon, 1990. Post-defaunation recovery of fish assemblages in southeastern blackwater streams. Ecology 71: 657–667.
Milner, A. M., A. L. Robertson, L. E. Brown, S. H. Sønderland, M. McDermott & A. J. Veal, 2011. Evolution of a stream ecosystem in recently deglaciated terrain. Ecology 92: 1924–1935.
Niemi, G. J., P. Devore, N. Detenbeck, D. Taylor, A. Lima, J. Pastor, J. D. Yount & R. J. Naiman, 1990. Overview of case studies on recovery of aquatic systems from disturbance. Environmental Management 14: 571–587.
Nyström, M., C. Folke & F. Moberg, 2000. Coral reef disturbance and resilience in a human-dominated environment. Trends in Ecology and Evolution 15: 413–417.
Oksanen, J., F. B. Guillaume, R. Kindt, P. Legendre, P. R. Minchin, R. B. O’Hara, G. L. Simpson, P. Solymos, M. H. H. Stevens & H. Wagner, 2013. vegan: Community Ecology Package. R package version 2.0-10. http://CRAN.R-project.org/package=vegan. Available 23 Jan 2014.
Olden, J. D., N. L. Poff & K. R. Bestgen, 2006. Life-history strategies predict fish invasions and extirpations in the Colorado River basin. Ecological Monographs 76: 25–40.
Peterson, J. T. & P. B. Bayley, 1993. Colonization rates of fishes in experimentally defaunated warmwater streams. Transactions of the American Fisheries Society 122: 199–207.
R Core Team, 2013. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna. http://www.R-project.org/.
Renforth, P., W. M. Mayes, A. P. Jarvis, I. T. Burke, D. A. C. Manning & K. Gruiz, 2012. Contaminant mobility and carbon sequestration downstream of the Ajka (Hungary) red mud spill: the effects of gypsum dosing. Science of the Total Environment 421–422: 253–259.
Resh, V. H., A. V. Brown, A. P. Covich, M. E. Gurtz, H. W. Li, G. W. Minshall, S. R. Reice, A. L. Sheldon, J. B. Wallace & R. C. Wissmar, 1988. The role of disturbance in stream ecology. Journal of the North American Benthological Society 7: 433–455.
Romme, W. H., M. S. Boyce, R. Gresswell, E. H. Merrill, G. W. Minshall, C. Whitlock & M. G. Turner, 2011. Twenty years after the 1988 Yellowstone fires: lessons about disturbance and ecosystems. Ecosystems 14: 1196–1215.
Sály, P., T. Erős, P. Takács, A. Specziár, I. Kiss & P. Bíró, 2009. Assemblage level monitoring of stream fishes: the relative efficiency of single vs. double pass electrofishing. Fisheries Research 99: 226–233.
Schiermeier, Q. & Y. Balling, 2010. Analysis lags on Hungarian sludge leak. Nature News 2010: 531.
Stanley, E. H., S. M. Powers & N. R. Lottig, 2010. The evolving legacy of disturbance in stream ecology: concepts, contributions and coming challenges. Journal of the North American Benthological Society 29: 67–83.
Therneau, T., E. J. Atkinson & B. Ripley, 2013. rpart: Recursive Partitioning. R package version 4.1-4. http://CRAN.R-project.org/package=rpart. Available on 21 Jan 2014.
Turner, M. G. & V. H. Dale, 1998. Comparing large, infrequent disturbances: what we have learned? Ecosystems 1: 493–496.
Turner, M. G., 2010. Disturbance and landscape dynamics in a changing world. Ecology 91: 2833–2849.
Vass, M., Á. Révay, T. Kucserka, K. Hubai, V. Üveges, K. Kovács & J. Padisák, 2013. Aquatic hyphomycetes as survivors and/or first colonizers after a red sludge disaster in the Torna stream, Hungary. International Review of Hydrobiology 98: 217–224.
Vaughn, C. C., 2012. Life history traits and abundance can predict local colonization and extinction rates of freshwater mussels. Freshwater Biology 57: 982–992.
Winemiller, K. O., 1992. Life-history strategies and the effectiveness of sexual selection. Oikos 63: 318–327.
Winemiller, K. O. & K. A. Rose, 1992. Patterns of life-history diversification in North American fishes: implications for population regulation. Canadian Journal of Fisheries and Aquatic Sciences 49: 2196–2218.
Zuur, A. F., E. N. Ieno & G. M. Smith, 2007. Analysing Ecological Data. Springer, New York.
Acknowledgments
This work was supported by the OTKA K104279 Grant and the Bolyai János Research Scholarship of the Hungarian Academy of Sciences (Tibor Erős). We thank Anna Dolezsai, Árpád Ferincz, Mónika Tóth, Zoltán Vitál, and numerous other people for their help in the field work.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: Judit Padisák
Rights and permissions
About this article

Cite this article
Erős, T., Takács, P., Czeglédi, I. et al. Taxonomic- and trait-based recolonization dynamics of a riverine fish assemblage following a large-scale human-mediated disturbance: the red mud disaster in Hungary. Hydrobiologia 758, 31–45 (2015). https://doi.org/10.1007/s10750-015-2262-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-015-2262-9