
Abstract
Soft corals show a wide variety of reproductive strategies, including both asexual and sexual reproduction which can influence macro-evolutionary processes. The octocoral Carijoa riisei has an ample geographical distribution in Pacific and southern Atlantic and also in Caribbean region. This species was considered invasive in Hawaii, and its capacity for rapid proliferation is recognized, but recent studies, however, have indicated that it appears to be native to the Indo-Pacific region. The present study examined the reproductive biology of C. riisei in the Atlantic basin. The results were then compared to previous studies of the same species from the Pacific (Hawaii) and Caribbean (Puerto Rico) regions to examine the hypothesis that the reproductive patterns of C. riisei populations are the same throughout its geographical distribution, independent of its native or non-native status. Samples were collected on a monthly basis from May/2007 to April/2008 at Porto de Galinhas (Pernambuco State, Brazil). This species was found to have similar reproduction patterns in Brazil, the Caribbean, and in Hawaii (a gonochoric reproductive pattern and continuous and asynchronous gamete release). The similarities of their traits contribute to its rapid proliferation and occupation of spaces left by other species, independent of its native or non-native status.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Sexual reproduction processes are usually fundamental to maintaining populations and influence macro-evolutionary processes (Kahng et al., 2011). Soft corals belong to the Order Alcyonacea and show a wide variety of reproductive strategies, including both sexual and asexual reproduction (e.g., Benayahu & Loya, 1984; Alino & Coll, 1989; Sammarco & Coll, 1992). Three gamete release strategies have been observed in these corals: gamete release with external fertilization (spawners); internal brooding; and external surface brooding (Benayahu et al., 1990; Benayahu, 1991; Dahan & Benayahu, 1997; Ben-David-Zaslow et al., 1999; Cordes et al., 2001; see a review by Kahng et al., 2011). Gametogenesis can be affected by environmental factors such as water temperature, lunar cycles and energetic resources (Ben-David-Zaslow et al., 1999; Zeevi Ben-Yosef & Benayahu, 1999), and different populations of the same species can show different reproductive strategies. Significant intraspecific variability of sexual attributes may, however, indicate that apparently conspecific organisms are actually morphologically similar cryptic sibling species (Babcock, 1995; Kahng et al., 2011).
The octocoral Carijoa riisei (Duchassaing & Michelotti, 1860) (Order Alcyonacea, Family Clavulariidae) is common from Florida (USA) to Santa Catarina (Brazil) and throughout the Caribbean region (Bayer, 1961; Pérez, 2002); it has also been recorded in Hawaii and other Pacific sites, as well as in the eastern Atlantic (Bayer, 1961; Concepcion et al., 2008). This species is very abundant along the Brazilian coast, especially in the northeastern region of that country, and previous studies have examined their associated fauna (Neves et al., 2007; Souza et al., 2007) and trophic ecology (Lira et al., 2009; Gomes et al., 2012).
Carijoa riisei was initially considered a non-indigenous and invasive species in Hawaii and other Pacific localities dispersed by marine vectors (Grigg, 2003), and was observed to successfully compete with, and even grow over, black coral and other native invertebrate forms (Kahng & Grigg, 2005; Kahng & Kelley, 2007). Recent studies using molecular techniques, however, indicated that the oldest populations of this species can be found in the Indo-Pacific basin, followed by Hawaii, while the youngest population analyzed was from the Atlantic-Caribbean region—so that it appears to be native to the Indo-Pacific region (Concepción et al., 2010). The ample geographical distribution of this species may reflect multiple introductions or ancient natural dispersals, although cryptic variations and the poorly resolved taxonomy of this species may have contributed to reports of its occurrence in many parts of the world (Concepcion et al., 2008, 2010). Concepcion et al. (2008), in fact, reported the existence of a cryptic species (transparent individuals of C. riisei) in Hawaii.
While studies on the reproductive biology of C. riisei have been conducted in Hawaii (Kahng et al., 2008) and Puerto Rico (Bardales 1981 in Kahng et al., 2011), there have been no previous examinations of the sexual reproduction of established populations in the southern Atlantic Ocean region. Considering the ample geographical distribution of this species, the unresolved species-level taxonomy of this genus in the Indo-Pacific region (Fabricius & Alderslade, 2001), and its capacity for rapid proliferation (as was seen in mesophotic reef habitats in Hawaii), the present study should provide important insights into the ecology of this puzzling species.
We, therefore, sought to characterize the sexual reproduction of C. riisei in the tropical South Atlantic and compare the results to previous Hawaiian and Caribbean investigations to examine the hypothesis that the reproductive patterns of C. riisei are the same in different populations, and that its rapid proliferation is characteristic of the species, independent of its native or non-native status.
Materials and methods
Study site and sampling
The present study was undertaken on coastal reefs at Porto de Galinhas beach, Pernambuco State, Brazil (8°30′20″S × 35°00′34″W) (Fig. 1) in a natural reef pool up to 8 m deep that is locally known as “Piscina dos 8” or “Boca da Barra”. Sampling was carried out along a 6-m vertical wall of an established population of C. riisei extending from the water surface to the ocean floor.

Geographical localization of the Porto de Galinhas beach on the coast of Pernambuco State, Brazil
The local climate is tropical humid with a mean annual temperature of 25°C (with fluctuations smaller than 5°C), a dry season (less than 100 mm/year) from September to February, and a rainy season (1,850–2,364 mm/year) from March to August (Medeiros et al., 1999). Water salinity (using a refractometer) and temperature (using a manual instant-read thermometer) were measured on each sampling occasion and remained stable throughout the study period, with means of 37.7 ± 0.56‰ and 27.6 ± 0.41°C, respectively, values within the normal range for the area (Medeiros et al., 1999).
In order to study the cycle of gonad development of this species and determine its reproductive season, samples were collected on a monthly basis from May/2007 to April/2008 (except in March/2008, due to persistent bad weather conditions). Fifty axial polyps were haphazardly collected each month at two depths (25 samples at two meters, and 25 samples at six meters). Specimens were collected from discontinuous aggregations at least 2 m apart to reduce the probability of sampling the same genet multiple times. The two depths were sampled only to increase the variability of samples and the power of inference of the results, since the colonies at 6 m were higher, despite being the same species. The study did not aim to compare the profundities as they were very close. The samples were placed in tagged plastic bags in situ, fixed in 4% formaldehyde in seawater for 24 h, washed with running fresh water in the laboratory, and subsequently preserved in 70% alcohol. The number of colonies, polyps, and oocytes used in each studied parameter is presented in Table 1.
Sexuality and sex ratios
Ten secondary polyps (emerging from axial polyps) and ten tertiary polyps (emerging from secondary axial polyps) were randomly removed from each sample and dissected under a stereomicroscope to determine their sexual categories: male, female, sterile (no gonads), or undefined (the gender could not be determined through gonad analysis). The data subsequently analyzed by log-linear testing using Systat 8.0 software (Wilkinson, 1996), considering the collection months.
The sex ratios were determined for the whole study period; significant deviations from 1:1 ratios were determined using Chi square testing (χ 2) (Sokal & Rohlf, 1996). The height of the primary axial polyp of each colony was measured (in millimeters) using a digital caliper to assess colony size distributions. The measure was taken from the basis, the point of fixation of the colony in the substrate (where the colony was cut during the sampling), until the tip of the axial primary polyp. The Chi square test was also used to compare the distributions of colony size categories; and the sizes of fertile and sterile colonies were compared (t test) using Systat 8.0 software (Wilkinson, 1996). Colony size at first sexual maturation was also assessed.
Gametogenesis and the reproductive cycle
Three polyps were randomly selected among the sampled colonies on a monthly basis for gametogenesis characterization. The polyps were immersed in 2% nitric acid for 24 h for decalcification (modified from Mahoney, 1966), dehydrated in pure ethanol, diaphanized in xylene, and embedded in paraffin; 6–8 µm histological sections were subsequently stained with hematoxylin and eosin (HE) for microscopic analyses, according to Belém (1987). Slides of gonads in different stages of development were photographed under a microscope.
The polyps of the female colonies were dissected under a stereomicroscope to characterize the reproductive statuses of the corals. The diameters of 1,100 oocytes were measured and divided into three size classes corresponding to their developmental stages (primordial: <100 μm diameter; intermediary: 100–320 μm; mature: >320 μm) modified from Zeevi Ben-Yosef & Benayahu (1999). The frequencies of the classes were assessed each month using log-linear testing. A histogram was constructed of total and monthly oocyte size distributions (divided into smaller intervals) to evaluate the reproductive cycle of this species.
Ten colonies were analyzed, five in January (dry season) and five in June (rainy season), to assess possible oogenesis synchronization. Five polyps from each of these ten colonies were randomly selected and dissected, and the diameters of their oocytes measured to calculate the percentages of polyps at each stage. Synchronization levels were determined by the size frequency distributions of oocytes among polyps within the same colony (intra-colony synchronization) and among polyps of different colonies (inter-colony synchronization).
The fertilities of male and female colonies were calculated in two ways: (1) as the percentage of fertile polyps containing gonads; and (2) as the percentages of female and male polyps having mature gonads. Both fertility values were analyzed using log-linear testing, considering polyp sex and the degree of ramification of each polyp (secondary or tertiary). Fecundity was calculated as the average number of mature oocytes per polyp. The significance level used in all statistical tests was α = 0.05.
Results
Sexuality and sex ratios
All of the C. riisei colonies examined were gonochoric, as determined by microscopic and histological analyses. Of the 550 colonies examined, 202 (36.7%) were female, 187 (34%) were male, 22 (4%) were indeterminate, and 139 (25.3%) were sterile. The sexual category percentages varied considerably during the year (χ 2 = 7.94; d.f. = 3; p = 0.0472), but no seasonal pattern could be determined (Fig. 2a). Fertile colonies were more frequent than sterile colonies (χ 2 = 9.056, d.f. = 1, p = 0.0026), with frequency differences between female and sterile colonies (χ 2 = 9.056, d.f. = 1, p = 0.0026) and between male and sterile colonies (χ 2 = 7.574, d.f. = 1, p = 0.0059). The sex ratio was 1:1, without any significant differences between male and female colony frequencies during the year.

a Monthly frequency distributions of the sexual categories of Carijoa riisei colonies (n = 100 each month); b Frequency distributions of the sizes (µm) of Carijoa riisei oocytes (n = 50 each month) during the study period at Porto de Galinhas beach, Pernambuco State, Brazil
Carijoa riisei colonies in the study sites were between 37 and 355 mm in height and the predominant size class (accounting for up to 72% of the samples) was 80–160 mm. The mean fertile colony size was 132 ± 2.11 mm (mean ± standard deviation; n = 411), while the mean sterile colony was 118.6 ± 3.68 mm (n = 139); being significantly different (t = 3.192; d.f. = 525; p = 0.0015). Large and small colonies of both genders were found. The smallest colony with spermaries was 52.8 mm in height, while the smallest with oocytes was 53.9 mm.
Gametogenesis descriptions and the reproductive cycle
The gonads of both genders developed along the mesenteries. Immature gonads were attached to the mesentery by a pedicel, gradually becoming detached during maturation. All of the developmental stages of C. riisei gonads were observed in the present study.
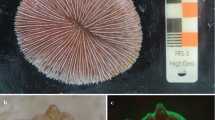
The oocytes were covered with a thick follicular cell layer that varied in color from transparent to peach, according to their diameters. The oocytes had well-defined internal regions with a readily visible nucleus (Fig. 3a, b). Oogenesis can be classified into three developmental stages. During stage 1, the oocytes were transparent due to an almost complete absence of cytoplasm, being closely grouped together and firmly attached to the mesentery. The diameters of these primordial oocytes ranged up to 100 μm, with a mean of 80.6 ± 15.7 μm (n = 282). In stage 2, intermediary oocytes were attached to the mesenteries by pedicels, with diameters ranging from 100 to 320 μm (mean 175.7 ± 52.5 μm; n = 547). These oocytes showed little pigmentation and could be identified by their light peach color and a distinct follicular layer. In stage 3, the oocytes had reached maturity with diameters greater than 320 μm (600 μm maximum, mean 466.7 ± 54.2 μm; n = 221), being weakly attached or totally detached from the mesenteries. Oocytes >300 μm in diameter were consistently pigmented and became darker as their diameters increased.

Gonads of Carijoa riisei a Histological section and b view under stereomicroscope of female polyp with oocytes at different stages of development (arrows): 1 Primordial; 2 Intermediary; 3 Intermediary oocyte attached to the mesentery by pedicel; c male polyp with spermaries in different stages of development
There were significant monthly differences among the three oocyte size class percentages during the entire study period (χ 2 = 74.94, d.f. = 20, p = < 0.0001) (Fig. 2b), with oocytes of all size classes (different developmental stages) being consistently found every month with bimodal frequency distributions, with a peak in classes <300 μm and a less prominent peak >400 μm (Fig. 4).

Bimodal frequency distributions of the sizes (µm) of Carijoa riisei oocytes during the study months at Porto de Galinhas beach, Pernambuco State, Brazil
Male colonies showed development patterns similar to those of female colonies, and all spermary developmental stages were recorded throughout the year. Their pigmentation development, color, and general appearances were similar to the oocytes (Fig. 3c), although mature spermaries were smaller than oocytes, ranging from 60 to 440 μm (average 247.87 ± 74.22 μm; n = 250).
The levels of intra-colony and inter-colony synchronization were often low, in either the dry (January) or rainy (June) season (Fig. 5). No significant differences were observed between the percentages of fertile polyps (with gonads) of males and females, or between secondary and tertiary polyps. No evidence for internal fertilization or for internal or external surface brooding was observed in the polyp cavities in histological cuts or in dissections of the 7191 polyps examined. Mature gonads were found in 37% of the polyps of female colonies and 49.20% of the polyps of male colonies (χ 2 = 7.975, d.f. = 1, p = 0.0047). Fertility was greater on tertiary polyps (χ 2 = 11.9, d.f. = 1, p = 0.0006). Average female fecundity, considering the numbers of mature oocytes per polyp, was 3.54 ± 2.44. This average was not determined for males, but it was found a large numbers of mature spermaries per polyp (often >100).

Intra-colony (among polyps of the same colony, numbered 1, 2, 3) and inter-colony (among colonies labeled A, B, C, D, E) synchronicity among female Carijoa riisei colonies sampled at Porto de Galinhas beach, Pernambuco State, Brazil in January (a) and June (b). (magnification 200×). Oocyte sizes (diameters) in µm
Discussion
Carijoa riisei has a gonochoric reproductive pattern in northeastern Brazil with a 1:1 sex ratio and continuous and asynchronous gamete release, similar to reports of populations in Hawaii and Puerto Rico (Table 2). Although soft coral sexuality has been reported to vary geographically (Benayahu et al., 1990; Ben-David-Zaslow et al., 1999), this has not been seen with C. riisei. The uniformity of its reproductive pattern indicates that this is an intrinsic characteristic of the species that allows its rapid proliferation.
Kahng et al. (2011) proposed that certain reproductive characteristics (such as oocyte sizes) are associated with subclades, while other characteristics are common to most octocorals (such as the prevalence of gonochorism and broadcast spawning). As certain reproductive characteristics may be conserved in the genus Carijoa, similarities between Caribbean, Pacific, and southern Atlantic basin populations do not necessarily prove their cospecificity. Comparative studies of morphological and molecular data from large numbers of populations in these localities will be necessary to confirm their co-specificity, as was noted by Concepcion et al. (2010).
The observed gonochorism of the C. riisei population in Brazil followed the usual octocoral pattern (see a review by Kahng et al., 2011). Both genders were reported on the same polyp mesentery in Hawaii, but these authors considered it to be a rare occurrence (Kahng et al., 2008). The regular occurrence of hermaphroditic and gonochoristic colonies has been reported before (McFadden 1999, 2001; Schleyer et al., 2004) but this may have been due to the presence of cryptic species (McFadden et al., 2006; Kahng et al., 2011).
The minimum size at which octocorals are capable of reproducing is variable (Benayahu & Loya, 1984; Coma et al., 1995). There are significant advantages to initiating sexual reproduction while the colonies are still small in unstable environments as mortality rates can be quite high (Excoffon et al., 2004), and accelerated initial growth can help colonies attain the minimum size necessary for reproduction—thus diminishing the risk of death before reproducing (Gutierrez-Rodriguez & Lasker, 2004). In all of the populations of C. riisei studied to date, the minimum reproductive size was ~≤5 cm (considering the height of the primary axial polyp) (Table 2). This species demonstrates rapid initial growth (Kahng 2006 in Kahng et al., 2008 for Hawaii; Perez, unpublished data, for Brazil) and can initiate reproduction just a few months after larval settlement (Kahng et al., 2008).
The sex ratio for C. riisei in the present study was the same as that reported in Hawaii and Puerto Rico (Kahng et al., 2008, 2011) and in other octocoral populations (Schleyer et al., 2004; Fan et al., 2005; Tsounis et al., 2006).Gonad production and development in C. riisei are similar to those processes seen in other octocorals, as the gonads are generated on the mesenteries and later migrate to the polyp cavities while still held on pedicels (Farrant, 1986; Benayahu, 1991; Dahan & Benayahu, 1997; Kapela & Lasker, 1999; McFadden & Hochberg, 2003; Gutierrez-Rodriguez & Lasker, 2004; Hwang & Song, 2007; Seo et al., 2008). Both male and female colonies in the C. riisei population studied here consistently showed spermaries and oocytes, respectively, at different development stages during every month throughout the year, indicating that they were continually reproducing and releasing gametes (as had been observed in both Hawaii and Puerto Rico) (Kahng et al., 2008, 2011). This behavior, associated with its capacity for vegetative reproduction, gives this species a significant advantage in colonizing new environments and increasing its geographical distribution (Kahng et al., 2008).
The maximum oocyte size of C. riisei (600 μm) was similar to that seen in Hawaii for the same species (Kahng et al., 2008). The mature oocytes of most alcyonaceans are ≥500 μm diameter (Choi & Song, 2007), even in species with different reproductive strategies such as internal brooding (Benayahu & Loya, 1983; Benayahu, 1991; Kruger et al., 1998), spawning (Benayahu & Loya, 1984, 1986; Farrant, 1986; Dahan & Benayahu, 1997; Kapela & Lasker, 1999; Schleyer et al., 2004; Fan et al., 2005), or external brooding (Brazeau & Lasker, 1990; Gutierrez-Rodriguez & Lasker, 2004). Most octocoral species with mature oocytes approximately 500 μm in diameter are found in tropical regions, although some inhabit temperate zones (Farrant, 1986; Excoffon et al., 2004; Hwang & Song, 2007; Seo et al., 2008). In their review of octocoral reproduction, Kahng et al. (2011) noted that zooxanthellate species have significantly larger oocytes than azooxanthellate species, suggesting that symbiotic relationship with photosynthetic microalgae aids in the provisioning of oocytes. In spite of the fact that C. riisei is azooxanthelate, its oocytes are large and well-provisioned, which would be expected to enhance their initial survival (Kahng et al., 2011). None of the studies undertaken to date with C. riisei has examined their larva to determine their pelagic duration or settlement patterns, factors that could also help explain its wide geographical distribution (the Indo-Pacific, Caribbean and Atlantic basins). Importantly, no indication of internal fertilization was noted among the 2000 polyps analyzed in Hawaii or the more than 7000 polyps analyzed in the present study—indicating that this species liberates its gametes directly into the water. Continuous reproduction without prolonged internal brooding probably facilitates the high and continuous production of planulae and guarantees free space in the polyp cavity for the development of additional oocytes (Vermeij et al., 2004). Bardales (1981 in Kahng et al., 2008) and Calcinai et al. (2004), however, assumed that C. riisei provided internal brooding, as they observed a single polyp with a planula larva inside. Loose oocytes have also been observed in the gastrovascular cavities of polyps in Indonesia (Calcinai et al., 2004), and these same authors encountered only female colonies, and smaller oocytes, than were observed in Hawaii or Brazil (maximum of 451 µm). These differences will have to be examined in more detail, as they may either reflect cryptic species or real changes in reproductive modes in different geographical regions.
Carijoa riisei was observed with oocytes in every developmental stage throughout the year, peaking in the <300 μm class. Mature oocyte frequencies were probably low because they are soon released in the environment for fertilization, while the primordial and intermediary stages remain connected to the mesenteries until maturation. This same pattern has been recorded in many gorgonian octocorals (Brazeau & Lasker, 1989; Brito et al., 1997; Orejas et al., 2002; Excoffon et al., 2004). The bimodal distribution of oocyte sizes demonstrated consistently low numbers of oocytes in the 300–400 μm range, suggesting that growth through this stage is very rapid and monthly collections could not be sufficient to detect this developmental stage, although it could also reflect the reabsorption of small oocytes. A similar situation was observed in Acabaria biserialis Kukenthal, 1913, in which size class distributions indicated fast initial development followed by slow and continuous maturation, and possibly the reabsorption of mature oocytes (Zeevi Ben-Yosef & Benayahu, 1999). Asynchronous spawning with continuous gamete release also allows C. riisei to more efficiently exploit substratum availability and favorable recruitment conditions (Kahng et al., 2008, 2011). Substrate availability to larval settlement seems to be related with reproductive strategy, even during an epibiosis relation. This is the case of the epibiotic soft coral Alcyonium coralloides that presents short gametogenesis and timing as an adaptation to optimize the colonization of its host, the gorgonian octocoral Paramuricea clavata (Quintanilla et al., 2013).
Coral reproduction in some species (including gamete maturation and release) has been directly related to high seawater temperatures (Harii et al., 2001; Neves & Pires, 2002; Vermeij et al., 2004). The resident population of C. riisei in Pernambuco is exposed to essentially constant high water temperatures throughout the year (with essentially no seasonal temperature variations), which could contribute to the liberation of gametes at any time. Food resource availability is also a critical factor for gonad production in azooxanthellate octocorals, as they depend on planktonic resources for the metabolic energy required for growth and reproduction (Fabricius et al., 1995a, b). Lira et al. (2009) and Gomes et al. (2012) undertook a study focusing on C. riisei in the same area as the present project, and encountered an ample spectrum of prey in the gastric cavities of its polyps throughout the entire year [predominately phytoplankton (diatoms and cyanophytes) and fragments of crustaceans]. Thus, high seawater temperatures and consistent food availability in the study area appears to facilitate the production of large numbers of gonads throughout the year in this octocoral.
It was not possible to determine the duration of the spermatogenic and oogenic cycles in C. riisei due to the continuous presence of gonads with oocytes and spermaries in all developmental stages throughout the entire study period. In alcyonaceans, the simultaneous development of multiple but distinct oocyte age/size classes has been associated with overlapping oogenic cycles lasting more than a year (Benayahu, 1997). Kahng et al. (2008) observed that the multiple overlapping of C. riisei oogenic cycles in Hawaii appeared to result from continuous breeding associated with high fecundity and rapid oogenesis. These same authors could not determine the duration of gametogenesis of C. riisei for the same reasons cited in the present study, but suggested that oogenesis may be shorter than a year, as inferred by the small sizes of these corals when they initiate their first reproductive cycle (male: 2.5 cm; female: 5 cm; Kahng et al., 2008) and their fast growth rates. Early first reproduction was also observed in the present study.
Variability of individual fertility and fecundity has frequently been reported within octocoral colonies (e.g., Brazeau & Lasker, 1990; Coma et al., 1995). Significantly different mature oocyte distributions were observed among secondary and tertiary polyps of C. riisei, although no significant differences were recorded among mature spermaries. Younger tertiary polyps emerging from secondary polyps showed higher fertility levels. These differences may be related to currents and to food resource availability or even to differentiation of the functions of polyps in relation to their positions within the colony, as was pointed out by Orejas et al. (2002). This same situation was not observed in male colonies, perhaps due to the fact that males generally require less energy for gametogenesis than females (Benayahu, 1997) and male invertebrates commonly released their gametes prior to the females (possibly stimulating oocyte release) (Levitan, 2005).
Concepcion et al. (2010) analyzed populations of C. riisei from Hawaii, from various other localities in the Indo-Pacific and Caribbean regions (Florida, Panama, and Puerto Rico/US Virgin Islands), and from a single locality in the southern Atlantic (the southeastern coast of Brazil) and demonstrated that the Brazilian samples shared alleles with all other Atlantic locations and also refuted a Caribbean-Atlantic introduction of Carijoa to Hawaii, however, a modern introduction from another source is still possible. This scenario goes against the view of the introduction of C. riisei from the Caribbean to Hawaii, as has been suggested by a number of authors (Thomas, 1979; Coles & Eldredge, 2002; Calcinai et al., 2004; Kahng & Grigg, 2005). These interesting results helped to define the biogeographical history of C. riisei, although a matching source population has not been found. Although potential cryptic species (Concepcion et al., 2008) were excluded from the molecular study, there is still the possibility of the existence of cryptic species in some of the numerous localities in which this taxon has been recorded.
As such, it is not yet possible to definitively determine if the presence of C. riisei in Hawaii is the result of a recent introduction followed by differential asexual proliferation of clones among the islands; the result of multiple introductions; or if the species was historically present but only recently detected, or combinations of any of these possibilities (Concepcion et al., 2010). Even if C. riisei had resided for a long period of time in Hawaii, its recent massive proliferation at depth throughout the many islands there was only recently noted. A number of biological characteristics apparently facilitated this process, including rapid growth, small size at the time of initial sexual maturation, gamete liberation throughout the year, and vegetative growth that allows rapid occupation of substrates and the exclusion of local species. This octocoral occupies many different natural and artificial substrates in Hawaii and directly competes with other species (Grigg, 2003; Kahng & Grigg, 2005; Kahng & Kelley, 2007). C. riisei is widely encountered on coastal reefs off the coast of northeastern Brazil, as well as on piers and wooden structures near estuaries, and is especially abundant on shipwrecks, thus constituting part of the fouling community (Amaral et al., 2009/2010). This distribution pattern has been observed along the coastal areas of other Brazilian states from Maranhão in the north to Santa Catarina in the south. Independent of the origin of C. riisei and of the processes that led to its introduction or dispersal over such an extensive geographical area, the species demonstrates rapid populational dynamics with elevated vegetative growth rates and continuous sexual reproduction. These characteristics favor its rapid proliferation in appropriate environments and could explain its populational explosion along mesophotic reefs in Hawaii. This appears to be an example of opportunist behavior, in which a given species can take advantage of reductions in the abundance of superior competitors, as has been reported with other cnidarians considered weedy species (Hawkins et al., 1999; Shaish et al., 2010).
The present study demonstrated that C. riisei populations have similar reproduction patterns in Brazil, the Caribbean, and in Hawaii. The similarities of their traits contribute to its rapid proliferation and occupation of spaces left by other species, independent of its native or non-native status. More detailed studies will be needed to firmly establish their historical–geographical relationships, including additional morphological and molecular investigations using larger numbers of survey populations.
References
Alino, P. M. & J. C. Coll, 1989. Observations of the synchronized mass spawning and postsettlement activity of octocorals on the Great Barrier Reef, Australia: biological Aspects. Bulletin of Marine Science 45: 697–707.
Amaral, F. D., C. M. R. Farrapeira, S. M. A. Lira & C. A. C. Ramos, 2009/2010. Benthic macrofauna inventory of two shipwrecks from Pernambuco coast, northeastern of Brazil. Revista Nordestina de Zoologia 4: 24–41.
Babcock, R., 1995. Synchronous multispecific spawning on coral reefs: potential for hybridization and roles of gamete recognition. Reproduction, Fertility and Development 7: 943–950.
Bayer, F., 1961. The shallow-water Octocorallia of the West Indian Region. A manual for marine biologists. Martinus Nijhoff, The Hague.
Belém, M. J. C., 1987. Anatomy and biology of Bunodosoma caissarum Corrêa, 1964 (Cnidaria, Anthozoa, Actiniidae) 1. Systematic position and morphological and microanatomical revision. Anais da Academia Brasileira de Ciências 59: 275–275.
Ben-David-Zaslow, R., G. Henning, D. K. Hoffmann & Y. Benayau, 1999. Reproduction in the Red Sea soft coral Heteroxenia fuscescens: seasonality and long-term record (1991 to 1997). Marine Biology 133: 553–559.
Benayahu, Y., 1991. Reproduction and developmental pathways of Red Sea Xeniidae (Octocorallia, Alcyonacea). Hydrobiologia 216(217): 125–130.
Benayahu, Y., 1997. Developmental episodes in reef soft corals: ecological and cellular determinants. In Lessios, H. A. & I. G. Macintyre (eds), Proceedings 8th International Coral Reef Symposium, Vol. 2., Smithsonian Tropical Research Institute Balboa, Panama 1997: 1213–1218.
Benayahu, Y. & Y. Loya, 1983. Surface brooding in the Red Sea soft coral Parerythropodium fulvum fulvum (Forskål, 1775). Biological Bulletin 165: 353–369.
Benayahu, Y. & Y. Loya, 1984. Life history studies on the Red Sea soft coral Xenia macrospiculata Gohar, 1940. I. Annual dynamics of gonadal development. Biological Bulletin 166: 32–43.
Benayahu, Y. & Y. Loya, 1986. Sexual reproduction of a soft coral: synchronous and brief annual spawning of Sarcophyton glaucum (Quoy and Gaimard, 1833). Biological Bulletin 170: 32–42.
Benayahu, Y., D. Weil & M. Kleinman, 1990. Radiation of broadcasting and brooding patterns in coral reef alcyonaceans. Advances in Invertebrate Reproduction 5: 323–328.
Brazeau, D. A. & H. R. Lasker, 1989. The reproductive cycle and spawning in a Caribbean gorgonian. Biological Bulletin 176: 1–7.
Brazeau, D. A. & H. R. Lasker, 1990. Sexual reproduction and external brooding of the Caribbean gorgonian Briareum asbestinum. Marine Biology 104: 465–474.
Brito, T. A. S., P. A. Tyler & A. Clarke, 1997. Reproductive biology of the Antarctic octocoral Thouarella variabilis Wright & Studer, 1889. In den Hartog, J. C. (ed.), Proceedings of the 6th International Conference on Coelenterate Biology, Nationaal Natuurhistorisch Museum, Leiden: 63–69.
Calcinai, B., G. Bavestrello & C. Cerrano, 2004. Dispersal and association of two alien species in the Indonesian coral reefs: the octocoral Carijoa riisei and the demosponge Desmapsamma anchorata. Journal of the Marine Biological Association of the United Kingdom 84: 937–941.
Choi, E. J. & J. I. Song, 2007. Reproductive biology of the temperate soft coral Dendronephthya suensoni (Alcyonacea: Nephtheidae). Integrative Biosciences 11: 215–225.
Coles, S. L. & L. G. Eldredge, 2002. Nonindigenous species introductions on coral reefs: a need for information. Pacific Science 56: 191–209.
Coma, R., M. Ribes, M. Zabala & J. M. Gili, 1995. Reproduction and cycle of gonadal development in the Mediterranean gorgonian Paramuricea clavata. Marine Ecology Progress Series 117: 173–183.
Concepcion, G. T., M. W. Crepeau, D. Wagner, S. E. Kahng & R. J. Toonen, 2008. An alternative to ITS, a hypervariable, single-copy nuclear intron in corals, and its use in detecting cryptic species within the octocoral genus Carijoa. Coral Reefs 27: 323–336.
Concepcion, G. T., S. E. Kahng, M. W. Crepeau, E. C. Franklin, S. L. Coles & R. J. Toonen, 2010. Resolving natural ranges and marine invasions in a globally distributed octocoral (genus Carijoa). Marine Ecology and Progress Series 401: 113–127.
Cordes, E. E., J. W. Nybakken & G. VanDykhuizen, 2001. Reproduction and growth of Anthomastis ritteri (Octocorallia: Alcyonacea) from Monterey Bay California, USA. Marine Biology 138: 491–501.
Dahan, M. & Y. Benayahu, 1997. Reproduction of Dendronephthya hemprichi (Cnidaria: Octocorallia): year-round spawning in an azooxanthellate soft coral. Marine Biology 129: 573–579.
Duchassaing, P. & J. Michelotti, 1860. Mémoire sur les Coralliaires des Antilles, Vol Imprimerie Royale, Turin.
Excoffon, A. C., F. H. Acuña, M. O. Zamponi, & G. N. Genzano, 2004. Reproduction of the temperate octocoral Tripalea clavaria (Octocorallia: Anthothelidae) from sublittoral outcrops off Mar del Plata, Argentina. Journal of the Marine Biological Association of the United Kingdom 84: 695–699.
Fabricius, K. E. & P. Alderslade, 2001. Soft Coral and Sea Fans: A Comprehensive Guide to the Tropical Shallow Water Genera of the Central-West Pacific, the Indian Ocean and the Red Sea. Australian Institute of Marine Science, Townsville: 264.
Fabricius, K. E., Y. Benayahu & A. Genin, 1995a. Herbivory in asymbiotic soft corals. Science 268: 90–92.
Fabricius, K. E., A. Genin & Y. Benayahu, 1995b. Flow-dependent herbivory and growth in zooxanthellae-free softcorals. Limnology and Oceanography 40: 1290–1301.
Fan, T. Y., Y. H. Chou & C. F. Daí, 2005. Sexual reproduction of the alcyonacean coral Lobophytum pauciflorum in southern Taiwan. Bulletin of Marine Science 76: 143–154.
Farrant, P. A., 1986. Gonad development and the planulae of the temperate Australian soft coral Capnella gaboensis. Marine Biology 92: 381–392.
Gomes, P. B., A. K. F. Lira, J.-P. Naud, A. M. Santos & C. D. Perez, 2012. Prey selectivity of the octocoral Carijoa riisei at Pernambuco, Brazil. Anais da Academia Brasileira de Ciências 84: 157–164.
Grigg, R. W., 2003. Invasion of a deep water coral bed by an alien species, Carijoa riisei. Coral Reefs 22: 121–122.
Gutierrez-Rodriguez, C. & H. R. Lasker, 2004. Reproductive biology, development, and planula behavior in the Caribbean gorgonian Pseudopterogorgia elisabethae. Invertebrate Biology 123: 54–67.
Harii, S., M. Omori, H. Yamakawa & Y. Koike, 2001. Sexual reproduction and larval settlement of the zooxanthellate coral Alveopora japonica Eguchi at high latitudes. Coral Reefs 20: 19–23.
Hawkins, J. P., C. M. Roberts, T. Van’T Hof, K. De Meyer, J. Tratalos & C. Aldam, 1999. Effects of recreational scuba diving on Caribbean coral and fish communities. Conservation Biology 13: 888–897.
Hwang, S. J. & J. I. Song, 2007. Reproductive biology and larval development of the temperate soft Dendronephthya gigantea (Alcyonacea: Nephtheidae). Marine Biology 152: 273–284.
Kahng, S. E. & R. W. Grigg, 2005. Impact of an alien octocoral, Carijoa riisei, on black corals in Hawaii. Coral Reefs 24: 556–562.
Kahng, S. E. & C. Kelley, 2007. Vertical zonation of habitat forming benthic species on the deep reef (60–150 m) in the Au’au Channel, Hawaii. Coral Reefs 26: 679–687.
Kahng, S. E., Y. Benayahu, D. Wagner & N. Rothe, 2008. Sexual reproduction in the invasive octocoral Carijoa riisei in Hawaii. Bulletin of Marine Science 82: 1–17.
Kahng, S. E., Y. Benayahu & H. R. Lasker, 2011. Sexual reproduction in octocorals. Marine Ecology and Progress Series 443: 265–283.
Kapela, W. & H. R. Lasker, 1999. Size-dependent reproduction in the Caribbean gorgonian Pseudoplexaura porosa. Marine Biology 135: 107–114.
Kruger, A., M. H. Schleyer & Y. Benayahu, 1998. Reproduction in Anthelia glauca (Octocorallia: Xeniidae). I. Gametogenesis and larval brooding. Marine Biology 13: 423–432.
Kukenthal, W., 1913. Ueber die Alcyonarienfauna Californiens und ihre tiergeographischen Beziehungen. Zoologische Jahrbucher, Abteilung für Systematik 35: 219–270.
Levitan, D. R., 2005. Sex-specific spawning behavior and its consequences in an external fertilizer. American Naturalist 165: 682–694.
Lira, A. K. F., J. P. Naud, P. B. Gomes, A. M. Santos & C. D. Perez, 2009. Trophic ecology of the octocoral Carijoa riisei from littoral of Pernambuco, Brazil. I. Composition and spatio-temporal variation of the diet. Journal of the Marine Biological Association of the United Kingdom 89: 89–99.
Mahoney, R., 1966. Laboratory Techniques in Zoology. Butterworth and Co, London.
McFadden, C. S., 1999. Genetic and taxonomic relationships among northeastern Atlantic and Mediterranean populations of the soft coral Alcyonium coralloides. Marine Biology 133: 171–184.
McFadden, C. S., 2001. A molecular phylogenetic analysis of reproductive trait evolution in the soft coral genus Alcyonium. Evolution 55: 54–67.
McFadden, C. S. & F. G. Hochberg, 2003. Biology and taxonomy of encrusting alcyoniid soft corals in the northeastern Pacific Ocean with descriptions of two new genera (Cnidaria, Anthozoa, Octocorallia). Invertebrate Biology 122: 93–113.
McFadden, C. S., P. Alderslade, L. P. van Ofwegen, H. Johnsen & A. Rusmevichientong, 2006. Phylogenetic relationships within the tropical soft coral genera Sarcophyton and Lobophytum (Anthozoa, Octocorallia). Invertebrate Biology 125: 288–305.
Medeiros, C., S. J. Macedo, F. A. N. Feitosa & M. L. Koening, 1999. Hydrography and phytoplankton biomass and abundance of North-East Brazilian waters. Archive of Fishery and Marine Research 47: 133–151.
Neves, E. G. & D. O. Pires, 2002. Sexual reproduction of Brazilian coral Mussismilia hispida (Verrill, 1902). Coral Reefs 21: 161–168.
Neves, B. M., E. J. B. Lima & C. D. Perez, 2007. Brittle stars (Echinodermata: Ophiuroidea) associated with the octocoral Carijoa riisei (Cnidaria: Anthozoa) from the littoral of Pernambuco, Brazil. Journal of the Marine Biological Association of the United Kingdom 87: 1263–1267.
Orejas, C., P. J. Lopez-Gonzalez, J. M. Gili, N. Teixido, J. Gutt & W. E. Arntz, 2002. Distribution and reproductive ecology of the Antarctic octocoral Ainigmaptilon antarcticum in the Weddell Sea. Marine Ecology Progress Series 231: 101–114.
Pérez, C. D., 2002. Octocorais (Cnidaria, Anthozoa) do litoral Pernambucano (Brasil). In Tabarelli, M. & J. M. Silva (eds), Diagnóstico da Biodiversidade de Pernambuco. Editora Massangana, Recife: 365–368.
Quintanilla, E., J. M. Gili, P. J. López-González, G. Tsounis, T. Madurell, I. Fiorillo & S. Rossi, 2013. Sexual reproductive cycle of the epibiotic soft coral Alcyonium coralloides (Octocorallia, Alcyonacea). Aquatic Biology 18: 113–124.
Sammarco, P. W. & J. C. Coll, 1992. Chemical adaptations in the Octocorallia: evolutionary considerations. Marine Ecology Progress Series 88: 93–104.
Schleyer, M. H., A. Kruger & Y. Benayahu, 2004. Reproduction and the unusual condition of hermaphroditism in Sarcophyton glaucum (Octocorallia, Alyoniidae) in KwaZulu-Natal, South Africa. Hydrobiologia 530(531): 399–409.
Shaish, L., G. Levya, G. Katzirc & B. Rinkevicha, 2010. Employing a highly fragmented, weedy coral species in reef restoration. Ecological Engineering 36: 1424–1432.
Seo, S. Y., J. H. Hwang & J. I. Song, 2008. Sexual Reproduction of Anthoplexaura dimorpha (Gorgonacea: Octocorallia) from Munseom, Jejudo Islands, Korea. Animal Cells and Systems 12: 231–240.
Sokal, R. R. & F. J. Rohlf, 1996. Biometry. Freeman & Company, New York.
Souza, J. R. B., H. A. Rodrigues, B. M. Neves & C. D. Pérez, 2007. First report of bristleworm predator of the reef octocoral Carijoa riisei. Coral Reefs 26: 1033.
Thomas, W. J., 1979. Aspects of the Micro-community Associated with Telesto riisei an Introduced Alcyonarian Species. Department of Zoology, University of Hawaii, Honolulu.
Tsounis, G., S. Rossi, M. Aranguren, M. Gili & W. Arntz, 2006. Effects of spatial variability and colony size on the reproductive output and gonadal development cycle of the Mediterranean red coral (Corallium rubrum L.). Marine Biology 148: 513–527.
Vermeij, M. J. A., E. Sampayo, K. Bröker & R. P. M. Bak, 2004. The reproductive biology of closely related coral species: gametogenesis in Madracis from the southern Caribbean. Coral Reefs 23: 206–214.
Wilkinson, L., 1996. Systat, Version 6.0. SPSS, Chicago, IL.
Zeevi Ben-Yosef, D. & Y. Benayahu, 1999. The gorgonian coral Acabaria biserialis: life history of a successful colonizer of artificial substrata. Marine Biology 135: 473–481.
Acknowledgments
The authors would like to thank the boatman Júnior (from Porto de Galinhas), the Porto Point Dive Shop, Aquaticus, David Belanger, and Diego Leonel Costa for their invaluable assistance with the field work. This work was supported by the Conselho Nacional de Pesquisa- CNPq (MCT/CNPq 14/2008-Universal, Grant number 480478/2008-2) and the Fundação de Aperfeiçoamento de Pessoal de Nível Superior-CAPES (T.M.B. Masters research fellowship).
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: I. A. Nagelkerken
Rights and permissions
About this article

Cite this article
Barbosa, T.M., Gomes, P.B., Bergeron, AS. et al. Comparisons of sexual reproduction in Carijoa riisei (Cnidaria, Alcyonacea) in South Atlantic, Caribbean, and Pacific areas. Hydrobiologia 734, 201–212 (2014). https://doi.org/10.1007/s10750-014-1893-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-014-1893-6