
Abstract
We investigated the distributional patterns of shallow-water fish and their environmental correlates along a broad spatial scale encompassing estuarine and freshwater ecosystems. Marine-vagrant species were restricted to the sites under the influence of salinity intrusion, whereas estuarine-related and freshwater guilds were found along the entire estuarine–freshwater gradient. Primary- and secondary-freshwater fish guilds had the most widespread spatial distribution and comprised a major fraction of the total abundance and species richness. Abiotic factors correlated with fish abundance and composition along two main environmental axes, one related with variation in salinity, water transparency, and sediment granulometry and the other with the slope gradient. Species richness was significantly higher at the natural channel linking the estuarine- and freshwater-ecosystem, which probably was due to: (a) a steeper slope that favored the confluence of fish from the littoral (<2 m) and limnetic (>2 m) zones and (b) the sporadic inflow of saltwater that carried into this region several marine-related species. Although estuarine–freshwater ecotones are known to support few species, mainly salinity tolerant, our results suggest that habitat features and seasonal fish movement associated with salinity intrusion could lead to more diverse fish assemblages in this transitional zone.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Estuaries are characterized by transitional communities containing freshwater, estuarine, and marine taxa, usually dominated by species with extreme physiological tolerances (Rundle et al., 1998). These estuarine-related species can have high biomass and sustain commercially important artisanal and coastal fisheries (Day et al., 1989; Blaber, 2000). It is usually assumed that abiotic factors (e.g., salinity, temperature, turbidity, dissolved oxygen, and depth) may define community structure in these variable systems, whereas biotic interactions may refine species distributional patterns within that structure (Sanders, 1968; Menge & Olson, 1990; Martino & Able, 2003). Among abiotic factors, variation in salinity is frequently recognized as the primary factor influencing species distributional patterns in estuaries (Day et al., 1989; Martino & Able, 2003; Vega-Cendejas & Santillana, 2004). The distribution of fish species along estuarine–freshwater gradients and the factors that govern its dynamics have been mainly investigated at warm-temperate and tropical estuaries (Winemiller & Leslie, 1992; Jung & Houde, 2003; Martino & Able, 2003; Akin et al., 2005; Sosa-López et al., 2007).
Prior studies on fish distribution along marine–estuarine and freshwater ecosystems in warm-temperate estuaries of North America and Europe show that marine and estuarine-dependent fish guilds have a greater influence on species richness and abundance patterns than freshwater species (Jung & Houde, 2003; Martino & Able, 2003; Akin et al., 2005). Akin et al. (2005), for instance, showed that 88.1% of the 42 species distributed along a salinity gradient (5–40 ppt) on the northwestern Turkish coast of the Mediterranean were marine or estuarine-dependent fish and only 7.1% were freshwater fish. A different pattern would be expected in estuaries located in the Neotropical zoogeographic region, whose rivers, streams, and ponds are home to the largest freshwater fish fauna in the world (Blaber, 2000; Barletta et al., 2003, 2005; Buckup et al., 2007; Helfman et al., 2009). A cross-system comparison carried out by Vieira & Musick (1994) suggest that estuaries of western Atlantic are more diverse than warm-temperate ones probably because of the greater number of estuarine-opportunist fishes, such as freshwater species, found in tropical estuaries. Other studies showed that primary- and secondary-division freshwater fish can be flushed in great number into a subtropical estuary of the Neotropics during heavy rainfall episodes (Garcia et al., 2003a, 2004). In this context, we hypothesized that primary- and secondary-division freshwater fish guilds would be the most diverse fish guilds along an estuarine–freshwater ecosystem in the Neotropics. Currently, little published information on large-scale fish distribution along estuarine–freshwater ecosystems in the Neotropics are available to evaluate this hypothesis (Winemiller & Leslie, 1992; Jaureguizar et al., 2003; García et al., 2010). As pointed out by Martino & Able (2003) most studies are restricted to oligohaline–polyhaline salinity gradients and do not encompass a complete environmental gradient from estuarine to freshwater conditions.
In order to evaluate our hypothesis, we investigated the abundance and diversity patterns of shallow-water fish assemblages along a broad (~300 km) environmental gradient from estuarine to freshwater in a subtropical coastal lagoon complex in the Neotropics. This complex is formed by the confluence of two important bodies of water: Patos Lagoon, which is considered the largest “choked” coastal lagoon (sensu Kjerfve, 1986) in the world, and Mirim Lake, the second largest lake in Brazil. We also analyzed the relationships between fish abundance and abiotic factors to investigate the relative importance of physicochemical variables (e.g., salinity, temperature, and water transparency) and structural characteristics of the habitat (substrate granulometry and organic matter content, slope, and depth) in structuring fish assemblages along the estuarine–freshwater gradient.
Materials and methods
Study area and data collection
Situated in extreme southern Brazil, the Patos–Mirim lagoon complex (PMLC) is formed by the confluence of two large aquatic systems: the Patos Lagoon (10,360 km2) and Mirim Lake (3,750 km2), with a drainage basin of 263,876 km2 (Seeliger et al., 1997). In the past, the brackish waters from Patos Lagoon estuary could reach Mirim Lake through a 75-km long natural channel (locally known as “São Gonçalo”). This salinity intrusion used to damage inundated rice crops in the region. In order to prevent such episodes, a subsurface dam of concrete (245 m long) was built across the channel in 1977 to block denser brackish waters. A lock was also built adjacent to the dam, in the east bank of the channel, to allow the passage of fishing boats and amateur sailboats along the São Gonçalo natural channel. This lock is 17 m wide and extends for 120 m along the channel and operates four times during the day, with each operation lasting for approximately 25 min. After the São Gonçalo dam was built, Mirim Lake waters remained fresh all the time, with an estimated mean overflow into Patos Lagoon of 700 m3/s (CLM, 1970; Machado, 2007; Hirata et al., 2010).
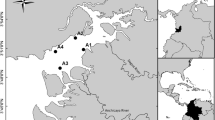
Our study area encompasses the shallow-water regions (<2 m) along the PMLC, with sampling sites in the estuarine zone of Patos Lagoon (E1–E2), São Gonçalo natural channel (C1–C2), and Mirim Lake (L1–L2–L3). These study regions are hereafter referred as estuary, natural channel, and freshwater lake, respectively. In total, there were seven sampling sites along a spatial axis of approximately 326 km (Fig. 1).

PMLC (A) showing the location of the sampling sites in estuarine zone of Patos Lagoon (E1–E2), São Gonçalo’s natural channel (C1–C2) and Mirim Lake (L1–L2–L3) (B)
Fish were collected monthly during March 2004 and February 2005 using a beach seine net (9 m wide and 2.4 m high; 13 mm mesh) with a pocket of 5 mm mesh in its rear portion. A total of five beach seine hauls were conducted in each of the seven sampling sites per month, with each haul considered as a sample. Following the sampling, fish were fixed in 10% formaldehyde and subsequently identified at the species level and each individual was measured for total length (TL) in millimeters (mm) and weighted in grams (g) at the Ichthyology Laboratory of the Rio Grande Federal University. If the number of individuals of a given species in a sample was greater than 50, we randomly picked 50 specimens for measuring. The other individuals of the same species in that sample were counted and added to the total. Species were identified according to Figueiredo & Menezes (1978, 1980), Menezes & Figueiredo (1980), Reis et al. (2003), and Buckup et al. (2007). At the same time as the fish were collected, salinity and temperature were measured using an YSI multiparameter probe (model 30–50 FT), the water transparency was measured (in cm) using a Secchi disk, and the maximum depth (cm) was recorded at each site.
To characterize the structural heterogeneity and complexity of the habitat, we measured the substrate granulometry and beach slope of each of the seven sampling sites. This characterization was conducted seasonally between 2009 and 2010. For the granulometry analysis, 60 g of sediment samples were collected using a 50-mm diameter PVC pipe at each site. The sediment samples were transported in coolers on ice and preserved in a freezer in the laboratory. The organic matter content of the sediment was determined by burning 5 g of the material in a muffle furnace at 550°C for 8 h. The amount of organic matter present in each sample was estimated as the difference between the initial and final mass.
For the granulometry analysis, the samples were dried in an oven at 60°C and then quartered and weighted. A total of 50 g of each dry sample was sifted on intervals of 1/2 phi according to the Wentworth classification to separate the coarse sediments (fine, medium, and coarse sand) from the finer sediment fractions (silt and clay, which were separated from each other) (Wentworth, 1922). The coarse and finer fraction of the sediment added up to 100% in each sample.
Although the substrate granulometry characterization was carried out approximately 4 to 5 years apart from the fish sampling, we assumed no major differences in granulometry composition among these years in this study site. Prior sedimentological surveys in Patos Lagoon corroborate this assumption (Calliari et al., 1977; Calliari, 1997). For instance, Calliari and collaborators analyzed the sediment composition along a depth profile in the upper portion of Patos Lagoon estuary in 1977 and found that sand (which correspond to coarse sediment in the current work) is the dominant fraction in the shallower waters, whereas the proportion of silt and clay (which correspond to fine sediment in the current work) increases with the depth. A map of the distribution of bottom sediment in this entire region published in 1997 also revealed a similar pattern (Calliari, 1997).
The slope at each site at each season was obtained from a topographic profile measured using a centimeter-scale ruler (Raseira, 2003). Depth measurements were taken at every 5 m from the swash zone to the maximum depth of collection at each site. We computed a slope index (Sind) based on the maximum depth/maximum perpendicular distance from the shoreline ratio. We used this ratio to classify sampling sites into three groups: beaches with low (<1.5), intermediate (5–10), and high (>10) slope.
Data analysis
Species composition, abundance and diversity patterns
The species composition and patterns of dominance were analyzed based on the frequency of occurrence (%F) and relative abundance (catch per unit effort; %CPUE) of each species (Garcia et al., 2004). Species with %F values equal to or greater than the average value of all species were considered frequent. Species with %F values smaller than the average value were rare. The same procedure was employed for the %CPUE values, resulting in higher abundant (%CPUE ≥ average %CPUE) and lower abundant species (%CPUE < average %CPUE). Finally, based on these criteria, the species were placed into four groups: (1) higher abundant and frequent, (2) higher abundant and rare, (3) lower abundant and frequent and (4) lower abundant and rare (Garcia et al., 2006). This classification was used to identify the dominant species. Thus, we considered species as dominant when the species was within first, second, and third categories in at least one of the sites (see Table 1). These criteria resulted in 31 dominant species, which were used in subsequent analysis (see below).
The MDS (non-metric multidimensional scaling) ordination technique was used to evaluate the similarity in species composition from a presence/absence matrix including all of the species and another matrix including the relative abundance of the dominant species. The relative-abundance data were square-root transformed, and the similarity matrix was calculated using the Bray–Curtis algorithm in the PRIMER statistical software package version 6.1.9 (Clarke & Gorley, 2006).
In some of the abundance analyses, the fish species were grouped into the following ecological guilds following Garcia et al. (2001, 2003b): (a) estuarine resident, fish that occur throughout the year and reproduce in the estuary; (b) estuarine-dependent, marine- or freshwater-spawning fish that occur in large numbers in the estuary at certain times of the year; (c) marine vagrant, fish coming from the marine environment that rarely occur in the estuary; (d) secondary freshwater, fish that are restricted to freshwater and occasionally occur in the estuary; and (e) primary freshwater, inhabitants of the limnic and fluvial environments that rarely occur in the estuary.
In the present study, however, the species Platanichthys platana and Jenynsia multidentata were considered as secondary-freshwater guild and not as estuarine residents as originally classified by Garcia et al. (2001, 2003b). This is because the current study was not restricted to the estuarine portion of the PMLC as prior studies (Garcia et al., 2001; 2003b), but instead, it also includes individuals of these species occurring in the freshwater reaches of this lagoon complex. In this context, we believe that “secondary-freshwater guild” is a more precise classification for these two species.
Two components of diversity were analyzed separately: observed species richness (S) and evenness. The evenness (i.e., the distribution of the total abundance among the species) was calculated by the modified (E5) Hill index (1973):
where D is the Simpson Index and H′ is the Shannon Index. This index assumes its maximum value when all the species in a sample are equally abundant and decreases toward zero as the difference in relative abundance between the species increases (Ludwig & Reynolds, 1988). We applied an arcsine transformation to the evenness data before the ANOVA test (Sokal & Rohlf, 1995).
The differences in the observed species richness (S) and evenness (E5) between seasons and sampling sites were examined using a two-way ANOVA. The assumptions of homogeneity of the variance and normality were evaluated using Cochran’s test and the Kolmogorov–Smirnov test, respectively (Sokal & Rohlf, 1995). The post hoc Tukey test was employed to determine which averages were significantly different from each other. The seasons of the year were defined as follows: summer (January, February, and March), autumn (April, May, and June), winter (July, August, and September) and spring (October, November, and December).
Relationships between fish abundance and abiotic factors
A two-way ANOVA was used to test for significant differences in salinity [log10 (x + 1)], water temperature and depth among months and sites. When a significant main effect was observed, pairwise comparisons of means were performed using Tukey post hoc procedure. The water transparency (Secchi disk) data did not meet the ANOVA assumptions, so the differences in water transparency among months and sites were tested using Kruskal–Wallis non-parametric test (Sokal & Rohlf, 1995; Conover, 1999).
Canonical correspondence analysis (CCA) was used to evaluate the relationships between abiotic factors including percentage of fine sediment, percentage of coarse sediment, organic matter, slope, depth of the haul, salinity, water temperature, and water transparency, and abundance of the dominant fish species (31 species). CCA analysis was performed using the software CANOCO for Windows 4.5.
In the CCA module, the unimodal method was used with biplot scaling, focusing on the inter-specific distance scale. We used the criterion proposed by Leps & Smilauer (2003) to choose between unimodal and linear ordination methods. According to this criterion when the gradient length, which measures the species turnover in the community composition, is larger than four (4), the unimodal method is more appropriate than the linear. The linear method would not be suitable because the data are too heterogeneous and too many species would deviate from the assumed model of linear response (Leps & Smilauer, 2003). The environmental factors were expressed as standardized scores (= (observed value − mean)/standard deviation), whereas fish abundance was expressed as log10 (CPUE + 1). The forward selection procedure was used to determine significant (α = 0.05) environmental variables explaining the observed variation in the species matrix. For those factors that were significantly correlated with each axis, a series of permutations were performed to determine which factors better explained the variation in the fish assemblage. The Monte Carlo permutation test (with 9999 permutations) was used to test the significance (P < 0.05) of the contribution of each factor to each of the CCA axes (ter Braak & Verdonschot, 1995).
Results
Spatiotemporal variation in physicochemical factors
The salinity varied from 0 to 29 ppt, but values above zero occurred only in the estuary (E1 and E2) and in the northern portion of the natural channel (C1), which is downstream of the São Gonçalo’s dam lock and has a free connection with the estuarine region of Patos Lagoon. With respect to seasonal variation, salinity was higher during the summer months (January and February) than during October (P < 0.05) (Fig. 2a). The water temperature varied from 10°C (July) to 31.2°C (January) and differed significantly among months (P < 0.05), with high values in the summer and low values in the winter. There were no significant differences among sites (P > 0.05) (Fig. 2b). The water transparency (Secchi depth) ranged from 8 to 100 cm. There were no significant differences among monthly Secchi disk depth (P > 0.05). However, water transparency of the estuary (E1 = 62.45 cm) and the natural channel (C1 = 27 cm and C2 = 27.5 cm) differed significantly (P < 0.05) (Fig. 2c). Water depth of sites varied from 10 to 130 cm, with no significant differences among months (P > 0.05). The average depth values were significantly higher (P < 0.05) at the sites of natural channel (~99 cm) than at the other collection sites (~50 cm) (Fig. 2d).

Spatiotemporal variation in average (+95% confidence intervals) values of salinity (a), water temperature (b), water transparency (c) and depth (d) in the estuarine zone of Patos Lagoon (E1–E2), São Gonçalo Channel (C1–C2) and Mirim Lake (L1–L2–L3)
Species abundance and diversity along the freshwater–estuarine gradient
A total of 45,929 individual fish, representing 11 orders and 25 families were collected. These were identified to 63 species, excluding those identified only to the level of the family Clupeidae, and some unidentified species of the genus Odontesthes (Table 1). Most of these 63 species were freshwater fish (61.9%), belonging to primary- (44.4%) and secondary-freshwater (17.4%) ecological guilds. The marine-vagrant, estuarine-dependent, and estuarine-resident guilds comprised 19.0, 11.1, and 7.9% of the collected fish species, respectively (Table 1).
The following ten species summed up nearly 90% of the total catch: the mullets Mugil liza and Mugil curema, the freshwater characids Cyanocharax alburnus, Cheirodon ibicuhiensis, Astyanax fasciatus, the sardines Brevoortia pectinata and P. platana, the silversides Odontesthes mirinensis and Odontesthes argentinensis, and the anchovy Lycengraulis grossidens.
The non-metric multidimensional analyses (MDS) based on the species composition (presence/absence) and abundance (%CPUE) of dominant species matrices resulted in similar patterns. Both MDS analyses supported two main groups, one dominated by estuarine samples (E1 and E2) and the other, the freshwater sites of natural channel region (C1 and C2) and freshwater lake (L1, L2, L3) (Fig. 3). The only exception was the occurrence of a single channel sample (C1) in the estuarine group. This sample came from the sampling site at the northern end of the natural channel, which is located downstream the São Gonçalo’s dam lock and has a free connection with the estuarine region of Patos Lagoon. It was collected in February and showed a typically estuarine ichthyofauna due to the inflow of saline water into the natural channel during January and February 2005. Excepting this, the samples were formed by two subgroups; channel and freshwater lake (Fig. 3).

MDS ordination of fish samples based on abundance of dominant species collected in the estuarine zone of Patos Lagoon (E1, E2), São Gonçalo Channel (C1, C2), and Mirim Lake (L1, L2, L3). Data values were square-root transformed and Bray–Curtis similarity were used. The solid line indicates the two main groups formed (estuary vs. freshwater samples), whereas the dotted line indicates the two sub-groups (Channel vs. Lake) within the freshwater group
According to the percentage similarity analysis (SIMPER), the estuarine group presented an average similarity of 28.45% in species composition, and the species that contributed the most to this group were M. liza, M. curema, B. pectinata, and O. argentinensis. The freshwater group formed by those species found in the natural channel and freshwater lake regions showed an average similarity of 19.57%. Cyanocharax alburnus, O. mirinensis, A. fasciatus, P. platana, C. ibicuhiensis, and Bryconamericus iheringii were the species that contributed most to this similarity.
A strong substitution of ecological guilds occurred between the estuarine zone (estuary and northern portion of the channel) and the limnic zone (southern portion of the channel and freshwater lake) (Fig. 4), coinciding with the decrease in salinity along the gradient (Fig. 2a). This substitution was gradual in terms of %F (Fig. 4a), but was more abrupt in terms of abundance (Fig. 4b). The marine-vagrant guild was restricted to the sites under influence of salinity intrusion (E1, E2, C1), whereas the other ecological guilds occurred along the entire estuarine–freshwater gradient. Both freshwater guilds, in particular, had the most widespread distribution, occurring across all sampling sites (from E1 to L3) (Fig. 4a). In terms of relative abundance, the estuarine-dependent guild was dominant in the estuarine region (with a mean of 85%), the primary-freshwater guild was dominant in the natural channel region (mean of 80%), and the primary- and secondary-freshwater guilds were dominant in the freshwater lake region (mean 51 and 45%, respectively).

Frequency of occurrence (%F) and relative abundance (%CPUE) of estuarine resident (ESTRES), estuarine dependent (ESTDEP), marine vagrant (MARVAG), and secondary freshwater (2ndFRESH), and primary freshwater (1stFRESH) in the estuarine zone of Patos Lagoon (E1–E2), São Gonçalo Channel (C1–C2) and Mirim Lake (L1–L2–L3)
Table 2 shows the ANOVA results concerning the variation in observed species richness and evenness among collection sites and seasons. For species richness, there was a significant interaction between sites and season (P < 0.01) (Table 2). In general, species richness was significantly higher on average at the natural channel sites (C1 and C2) than at the other sites (P < 0.00001) and significantly lower on average during the winter than during the summer and spring (P < 0.0318). More specifically, the highest average values in species richness were observed during autumn and spring at C1 and during summer, autumn, and spring at C2 (Fig. 5). Evenness differed significantly only among sites. The site C1 exhibited lower evenness compared to E2 and L3 but did not differ significantly from the other sites (Tukey test; P > 0.05).

Average values (+ standard error) of species richness (S) across sampling sites (E1, E2 estuarine zone of Patos Lagoon; C1, C2 São Gonçalo Channel; L1, L2, L3 Mirim lake) and seasons (filled triangle summer, empty circle autumn, filled square winter, filled square spring)
Relationships between fish abundance and abiotic factors
The results of the CCA are shown in Table 3. The total inertia was 2.995, of which 37% (1.115) was explained by the analysis. The first CCA axis had an eigenvalue of 0.661 and explained 22.1 and 59.3% of the species data and the species–environment relation variance. The second axis had an eigenvalue of 0.251, and the first and second axes combined accounted for 30.5 and 81.8% of the species data and the species–environment relation variance. The relationships between the dominant species and the environmental variables represented by these two axes were statistically significant (P < 0.05) according to the Monte Carlo permutation test. The depth and the percentage of coarse sediment did not contribute significantly to the model (FS test; P > 0.05) and were excluded from the analysis.
In general, the CCA analysis revealed that the relationship between fish abundance and abiotic factors occurred mainly across two contrasting environmental gradients (salinity-water transparency and the %fine sediment-organic matter) associated with axis 1, and also along a slope gradient represented by axis 2 (Fig. 6). Considering the axis 1, it can be observed that estuarine samples were strongly correlated with higher water transparency and salinity values, whereas samples from the natural channel were associated with higher percentages of fine sediment and organic matter. The slope gradient represented by axis 2, in contrast, depicted mainly differences between samples from the freshwater lake and the natural channel. The former were strongly correlated with lower slope values, whereas the latter with higher slope values (Fig. 6).

CCA ordination plot. a sample sites (black filled circle estuarine zone of Patos Lagoon, grey filled circle São Gonçalo Channel, empty circle Mirim Lake) and environmental variables (% fine granulometry, % organic matter, slope, salinity, water temperature, and water transparency). Each point represents a single sample site and date (five beach seine hauls at the location for a given month). b Association of fish species with each axis is represented by its score (plotted) and correlation between environmental variables and axes are represented by the length and angle of arrows. Refereed to Table 1 for species and guild codes
Species belonging to the estuarine-dependent and estuarine-resident guilds, which were abundant in the estuary, were associated with higher values along the salinity gradient, whereas primary- and secondary-freshwater species were found at the opposite far end of this gradient. Odonthestes argentinensis exhibited the strongest correlation with water transparency. The distributions of G. brasiliensis and C. punctata were closely related to slope, and those of Loricariichthys anus, Oligosarcus jenynsii, Charax stenopterus, C. ibicuhiensis, and Hyphessobrycon luetkenii were closely related to fine-grained sediment.
Discussion
Species abundance and diversity across the estuarine–freshwater gradient
Primary- and secondary-freshwater fish guilds comprised a major fraction of the total abundance and species richness along the studied gradient, not only in the limnic zone of the gradient (C2, L1–L3), but also in one of the sampling sites (C1) under the sporadic influence of salinity intrusion. Moreover, nine species of freshwater fish were also recorded, although in very low numbers, in those estuarine sites (E1, E2) located in the lower reaches of the estuary, where brackish water conditions are prevalent throughout the year. Previous studies on the inter-annual abundance of freshwater fishes in this estuary have shown that freshwater fish significantly increase their abundance and species richness in the estuarine zone during periods of higher freshwater discharge, usually associated with excessive rainfall triggered by strong El Niño events (Garcia et al., 2003a, 2004). Fish sampling in the current work was carried under the influence of the weak 2004–2005 El Niño event (Lyon & Barnston, 2005). Therefore, the abundance and diversity of freshwater fishes that we observed in the estuarine zone could be considered higher than normal (non-El Niño) conditions, but lower when compared with conditions influenced by strong El Niño events (Garcia et al., 2004).
Regardless El Niño events, several freshwater fish can also be carried into this estuary during prolonged above-average rainfall episodes associated with local (e.g., topography) and regional (e.g., cold fronts, anomalies of sea surface temperatures off the coast of southern Brazil) factors (Rao & Hada, 1990; Diaz et al., 1998; Kane, 1999). For instance, Garcia et al. (2003a) showed that the excessive rainfall in years between 2000 and 2001, which was not triggered by El Niño event, carried characid species into the estuary, particularly Astyanax eigenmaniorum and O. jenynsii. In addition, they showed that the occurrence of freshwater species from secondary-freshwater families, such as poeciliids and cichlids, which are tolerant to low salinity, are less correlated with hydrological conditions, and seemed to occur through active migration from nearby freshwater habitats draining into this area.
The conspicuous contribution of freshwater fish to the abundance and species richness patterns revealed in the current study seem to reflect the fact that this subtropical estuary is located within the Neotropical zoogeographic region, which has the richest freshwater ichthyofauna in the world (Buckup et al., 2007; Helfman et al., 2009). A similar pattern has also been reported on prior studies conducted in the Neotropics on fish distributions along large scale estuarine–freshwater gradients in tropical (e.g., Winemiller & Leslie, 1992) and subtropical (e.g., Castro et al., 2009; García et al., 2010) latitudes. In contrast, freshwater fish comprised a minor part of fish diversity in temperate estuaries of Southern Africa and Western Australia (Potter et al., 1990), North America (Jung & Houde, 2003; Martino & Able, 2003) and Europe (Akin et al., 2005), which are located within zoogeographic provinces comparatively poorer in freshwater fish species. Future large-scale studies encompassing distinct zoogeographical regions and employing standardized fish sampling protocol would be necessary in order to further evaluate the hypothesis that primary- and secondary-freshwater fish have a greater contribution on species composition and abundance along estuarine–freshwater gradients in the Neotropics when compared to other zoogeographic regions.
Our study also revealed that the highest species richness occurred during warmer months in the São Gonçalo natural channel that connects the Patos Lagoon estuary and the Mirim Lake. Two mechanisms seem to explain this pattern. First, the natural channel has a very narrow and steep shoreline (<2 m) reaching deeper waters (>3 m) only 1–2 m away from the shore, compared to the studied sites of the estuary and the freshwater lake where the distance from shore to deeper waters was >10 m. Consequently, the depth of the beach seine hauls employed during fish collection was significantly higher in this area (P < 0.05). This proximity between shoreline and deeper waters could favor the mixture of fish assemblages from the littoral and limnetic zones of the natural channel. In general, fish faunas of littoral and limnetic zones have distinct fish species compositions (Vieira & Musick, 1994; Lowe-McConnell, 1999; Garcia et al., 2006; Souza et al., 2008). For example, in lakes adjacent to the present study area, Garcia et al. (2006) observed that smaller species (e.g., C. alburnus, J. multidentata, B. iheringii) predominate in the littoral zone, whereas larger species (e.g., Cyphocharax voga, A. fasciatus, O. jenynsii, O. mirinensis) predominate in the limnetic zone. Second, the natural channel is subject to the sporadic inflow of saltwater during the year, usually in warmer months, especially at its northern reaches (site C1), which is nearer to the estuary. During these salinity intrusions, it is common to catch in this region several estuarine-dependent species (e.g., the sardine B. pectinata, the anchovy L. grossidens, the mullets M. liza and M. curema, the Whitemouth croaker Micropogonias furnieri). Hence, this natural channel can harbor freshwater and marine fish species alternately during the year. Moreover, an occurrence of a subsurface dam built in 1977 in the natural channel (approximately 7.5 km south of site C1) could favor the concentration and retention of freshwater and marine and estuarine-related fish fauna in this region. This could also have influenced the higher species richness observed in the natural channel. Therefore, the species richness in the natural channel seemed to be the result of a combination of factors related to bathymetry (i.e., the confluence of littoral and limnetic taxocenoses due to a steeper slope) and fish movement associated with salinity intrusion (i.e., the confluence of guilds with different degrees of salinity tolerance originated from marine/estuarine and limnic environments). Therefore, although estuarine–freshwater ecotones are known to support few species, mainly salinity tolerant (Day et al. 1989), our results suggest that a steeper slope favoring the confluence of fish from the littoral (<2 m) and limnetic (>2 m) zones and seasonal fish movement associated with salinity intrusion could lead to more diverse fish assemblages in our studied transitional zone.
As pointed out by Burns et al. (2006), a subsurface dam seems to disfavor movement of estuarine-related shallow-water fish along the natural channel toward Mirim Lake. Our findings revealed that ecological fish guilds showed a gradual substitution along the gradient in terms of %F suggesting that the dam is not a complete barrier for estuarine-related fish movement. For instance, water from upstream and downstream the dam can mix during the daily operation of the dam’s lock, which allows the passage of fishing boats and amateur sailboats along the São Gonçalo natural channel. Therefore, these lock operations could favor the movement of estuarine-related fish between both regions. However, such estuarine-related fish movement during lock operation seemed to be very restricted, since we observed an abrupt difference in the abundance of these fish guilds between the sampling sites located north (C1) and south (C2) of the dam.
It is also worth noting that water flows from Mirim Lake into Patos Lagoon during most of the year (CLM, 1970; Machado, 2007; Hirata et al., 2010), overflowing in these occasions the subsurface dam in the natural channel. Hence, in contrast to the estuarine-related fish, the movement of freshwater fish over the subsurface dam can occur much more often and independently of the lock operation. This could be one of the reasons why primary-freshwater fish were numerically dominant in the sampling site located north of the dam (C1). Another reason is the fact that this region remained fresh during almost the 12 months of the fish sampling, with the exception of a salinity intrusion observed on February 2005.
It seems reasonable to assume that the abundance of the ecological guilds would shift more gradually between the Patos Lagoon estuary and the Mirim Lake in the absence of this subsurface dam. Other studies on species distributions across large-scale estuarine–freshwater gradients not interrupted by dams seem to corroborate this assumption. For example, when analyzing fish assemblages from the adjacent marine area to the limnic zone of Patos Lagoon, Raseira (2003) observed that the abundance of estuarine-related species was gradually replaced by freshwater species. Likewise, García et al. (2010) observed that marine species gradually replaced the freshwater ichthyofauna along a marine–freshwater gradient in the La Plata River (Argentine), with no conspicuous transitional area (ecotone) in terms of fish abundance.
Relationships between fish abundance and abiotic factors
In general, salinity, slope, substrate granulometry (percentage of fine sediment), and water transparency were the factors that best explained the variability in the abundance of dominant fish species in the study area. With a few exceptions, the salinity gradient segregated our five fish guilds into two groups: One associated with the brackish estuarine waters, which was composed of estuarine-dependent, estuarine-resident, and marine-vagrant fish guilds, and other restricted to the limnic region, which was composed of primary- and secondary-freshwater species. Previous studies highlighted the predominant role of salinity in structuring estuarine fish assemblages (Martino & Able, 2003; Akin et al., 2005; Barletta et al., 2005). However, as discussed earlier, the marked segregation of guilds along the salinity gradient in the present study was probably strengthened by the presence of a subsurface dam in the middle portion of the gradient, which partially impaired the free movement of water and fish between the estuary and the limnic region upstream.
The importance of physicochemical factors (e.g., salinity, temperature) has been extensively investigated in previous studies (Martino & Able, 2003, Akin et al., 2005). Our results suggest the structuring role of sediment granulometry on structuring fish assemblages along estuarine–freshwater gradients. We found that the abundance of many species of the primary- and secondary-freshwater guilds was associated with the predominance of fine sediment (silt and clay) and with higher percentages of organic matter in the substrate. Some of the most abundant species in these guilds (e.g., L. anus, Geophagus brasiliensis, Gymnogeophagus gymnogenys, Corydoras paleatus, and Rineloricaria longicauda) exhibit morpho-anatomical adaptations, such as inferior mouth, sensory barbels, taste buds, to feed on epifaunal and infaunal prey, which are usually abundant in sediment with higher content of fine particles and organic matter of this region (Bemvenuti, 1997).
In summary, our study revealed that primary- and secondary-freshwater fish makes a significant contribution to abundance and species richness patterns along the estuarine–freshwater gradient of a subtropical estuary emptying in the Western Atlantic. Marine-vagrant species were restricted to sites under the influence of salinity intrusion, whereas estuarine-related and freshwater guilds were found along the entire estuarine–freshwater gradient. Primary- and secondary-freshwater fish, in particular, had the most widespread spatial distribution along the gradient. The abundance of ecological fish guilds along the gradient shifted more abruptly than expected due to the presence of a human interference (dam). Species richness was higher at the middle portion of the gradient, where a 75 km-long natural channel connects the brackish waters of Patos Lagoon estuary with the fresh waters of Mirim Lake. The unexpected higher species richness in this region probably results from two mechanisms: the confluence of littoral and limnetic fish species due to a steeper slope and to the mixture of marine/estuarine and freshwater guilds at different periods along the year that could be more concentrated in this region due to the presence of a dam. Abiotic factors seemed to structure fish abundance and composition across two main environmental CCA axes, one related with salinity, water transparency, and sediment granulometry (higher content of silt, clay and organic matter) and the other with habitat slope.
References
Akin, S., E. Buhan, K. O. Winemiller & H. Yilmaz, 2005. Fish assemblage structure of Koycegiz Lagoon-Estuary, Turkey: spatial and temporal distribution patterns in relation to environmental variation. Estuarine Coastal and Shelf Science 64: 671–684.
Barletta, M., A. Barletta-Bergan, U. Saint-Paul & G. Hubold, 2003. Seasonal changes in density, biomass, and diversity of estuarine fishes in tidal mangrove creeks of the lower Caete Estuary (northern Brazilian coast, east Amazon). Marine Ecology-Progress Series 256: 217–228.
Barletta, M., A. Barletta-Bergan, U. Saint-Paul & G. Hubold, 2005. The role of salinity in structuring the fish assemblages in a tropical estuary. Journal of Fish Biology 66: 45–72.
Bemvenuti, C. E., 1997. Benthic invertebrates. In Seeliger, U., C. Odebrecht & J. P. Castello (eds), Subtropical Convergence Environments. The Coast and Sea in the Southwestern Atlantic. Springer, Berlin: 43–46.
Blaber, S. J. M., 2000. Tropical estuarine fishes: ecology, exploitation and conservation. Blackwell, Oxford.
Buckup, P. A., N. A. Menezes & M. S. Ghazzi, 2007. Catálogo das espécies de peixes de água doce do Brasil. Museu Nacional da Universidade Federal do Rio de Janeiro, Rio de Janeiro. (in Portuguese).
Burns, M. D. M., A. M. Garcia, J. P. Vieira, M. A. Bemvenuti, D. M. L. M. Marques & V. Condini, 2006. Evidence of habitat fragmentation affecting fish movement between the Patos and Mirim coastal lagoons in southern Brazil. Neotropical Ichthyology 4: 69–72.
Calliari, J. L., 1997. Geological setting. In Seeliger, U., C. Odebrecht & J. P. Castello (eds), Subtropical convergence marine ecosystem. The coast and the sea in the warm temperate southwestern Atlantic. Springer, Berlin: 13–17.
Calliari, J. L., G. Griep & H. Vieira, 1977. Sedimentologic characteristics of the second profile of Benthos – Lagoa dos Patos Southern Portion. Atlântica 2: 63–82.
Castro, M. G., J. M. D. Astarloa, M. B. Cousseau, D. E. Figueroa, S. M. Delpiani, D. O. Bruno, J. M. Guzzoni, G. E. Blasina & M. Y. D. Antoni, 2009. Fish composition in a south-western Atlantic temperate coastal lagoon: spatial–temporal variation and relationships with environmental variables. Journal of the Marine Biological Association of the United Kingdom 89: 593–604.
Clarke, K. R. & R. N. Gorley, 2006. PRIMER v6: User Manual/Tutorial. PRIMER-E, Plymouth.
CLM, 1970. Comissão da Lagoa Mirim. Barragem do São Gonçalo: estudo preliminar de viabilidade. (in Portuguese).
Conover, W. J., 1999. Practical Nonparametric Statistics, 3rd ed. Wiley, New York.
Day, J. W. J., C. A. S. Hall, W. M. Kemp & A. Yañez-Arancibia, 1989. Estuarine Ecology. Wiley, New York.
Diaz, A. F., C. D. Studzinski & C. R. Mechoso, 1998. Relationships between precipitation anomalies in Uruguay and southern Brazil and sea surface temperature in the Pacific and Atlantic Oceans. Journal of Climate 11: 251–271.
Figueiredo, J. L. & N. A. Menezes, 1978. Manual de peixes marinhos do sudeste do Brasil. II Teleostei (1). Museu de Zoologia, Universidae de São Paulo, São Paulo. (in Portuguese).
Figueiredo, J. L. & N. A. Menezes, 1980. Manual de peixes marinhos do sudeste do Brasil. III Teleostei (2). Museu de Zoologia, Universidade de São Paulo, São Paulo. (in Portuguese).
Garcia, A. M., J. P. Vieira & K. O. Winemiller, 2001. Dynamics of the shallow-water fish assemblage of the Patos Lagoon estuary (Brazil) during cold and warm ENSO episodes. Journal of Fish Biology 59: 1218–1238.
Garcia, A. M., M. B. Raseira, J. P. Vieira, K. O. Winemiller & A. M. Grimm, 2003a. Spatiotemporal variation in shallow-water freshwater fish distribution and abundance in a large subtropical coastal lagoon. Environmental Biology of Fishes 68: 215–228.
Garcia, A. M., J. P. Vieira & K. O. Winemiller, 2003b. Effects of 1997–1998 El Nino on the dynamics of the shallow-water fish assemblage of the Patos Lagoon Estuary (Brazil). Estuarine Coastal and Shelf Science 57: 489–500.
Garcia, A. M., J. P. Vieira, K. O. Winemiller & A. M. Grimm, 2004. Comparison of 1982–1983 and 1997–1998 El Niño effects on the shallow-water fish assemblage of the Patos Lagoon Estuary (Brazil). Estuaries 27: 905–914.
Garcia, A. M., M. A. Bemvenuti, J. P. Vieira, D. M. L. M. Marques, M. D. M. Burns, A. Moresco & M. V. L. Condini, 2006. Checklist comparison and dominance patterns of the fish fauna at Taim Wetland, South Brazil. Neotropical Ichthyology 4: 261–268.
García, M. L., A. J. Jaureguizar & L. C. Protogino, 2010. From fresh water to the slope: fish community ecology in the Rio de la Plata and the sea beyond. Latin American Journal of Aquatic Research 38: 81–94.
Helfman, G. S., B. B. Collette, D. E. Facey & B. W. Bowen, 2009. The Diversity of Fishes: Biology, Evolution, and Ecology. Wiley-Blackwell, Hoboken.
Hill, M. O., 1973. Diversity and evenness – unifying notation and its consequences. Ecology 54: 427–432.
Hirata, F. E., O. O. Möller & M. M. Mata, 2010. Regime shifts, trends and interannual variations of water level in Mirim Lagoon, southern Brazil. Pan-American Journal of Aquatic Sciences 5: 254–266.
Jaureguizar, A. J., R. Menni, C. Bremec, H. Mianzan & C. Lasta, 2003. Fish assemblage and environmental patterns in the Rio de la Plata estuary. Estuarine, Coastal and Shelf Science 56: 921–933.
Jung, S. & E. D. Houde, 2003. Spatial and temporal variabilities of pelagic fish community structure and distribution in Chesapeake Bay, USA. Estuarine Coastal and Shelf Science 58: 335–351.
Kane, R. P., 1999. Rainfall extremes in some selected parts of central and south America: ENSO and other relationships reexamined. Journal of Climatology 19: 423–455.
Kjerfve, B., 1986. Comparative oceanography of coastal lagoons. In Wolfe, D. A. (ed.), Estuarine Variability. Academic Press, New York: 63–81.
Leps, J. & P. Smilauer, 2003. Multivariate analysis of ecological data using CANOCO. Cambridge University Press, New York.
Lowe-McConnell, R. H., 1999. Estudos ecológicos de comunidades de peixes tropicais (A.E.A.M. Vazzoler, A.A. Agostinho & P.T.M. Cunningham, trad.). Editora da Universidade de São Paulo, São Paulo. (in Portuguese).
Ludwig, J. A. & J. F. Reynolds, 1988. Statistical Ecology: A Primer on Methods and Computing. Wiley, New York.
Lyon, B. & A. G. Barnston, 2005. The evolution of the weak El Niño of 2004–2005. U.S. CLIVAR Variations, 3(2), US CLIVAR Office, Washington, DC: 1–4.
Machado, G., 2007. Demanda e disponibilidade hídrica no sistema Lagoa Mirim-São Gonçalo – Rio Grande do Sul. Revista Discente Expresões Geográficas 3: 61–82.
Martino, E. J. & K. W. Able, 2003. Fish assemblages across the marine to low salinity transition zone of a temperate estuary. Estuarine Coastal and Shelf Science 56: 969–987.
Menezes, N. A. & J. L. Figueiredo, 1980. Manual de peixes marinhos do sudeste do Brasil. IV. Teleostei (3). Museu de Zoologia, Universidade de São Paulo, São Paulo. (in Portuguese).
Menge, B. A. & A. M. Olson, 1990. Role of scale and environmental-factors in regulation of community structure. Trends in Ecology & Evolution 5: 52–57.
Potter, I. C., L. E. Beckley, A. K. Whitfield & R. C. J. Lenanton, 1990. Comparisons between the roles played by estuaries in the life cycles of fishes in temperate western Australia and southern Africa. Environmental Biology of Fishes 28: 143–178.
Rao, V. B. & K. Hada, 1990. Characteristics of rainfall over Brazil: annual variations and connections with the southern oscillation. Theoretical and Applied Climatology 42: 81–91.
Raseira, M. B., 2003. Análise espaço-temporal da associação de peixes das zonas rasas da Lagoa dos Patos e região costeira adjacente, Pós Graduação em Oceanografia Biológica, Dissertação de mestrado, Universidade Federal do Rio Grande, Rio Grande, 113. Unpublished. (in Portuguese).
Reis, R. E., S. O. Kullander & C. J. Ferraris, 2003. Check List of the Freshwater Fishes of South and Central America. EDIPUC, Porto Alegre.
Rundle, S. D., M. J. Attrill & A. Arshad, 1998. Seasonality in macroinvertebrate community composition across a neglected ecological boundary, the freshwater–estuarine transition zone. Aquatic Ecology 32: 211–216.
Sanders, H. L., 1968. Marine benthic diversity – a comparative study. American Naturalist 102: 243–282.
Seeliger, U., C. Odebrecht & J. Castello, 1997. Subtropical Convergence Environments: The Coast and Sea in the Southwestern Atlantic. Springer, New York.
Sokal, R. R. & F. J. Rohlf, 1995. Biometry: The Principles and Practice of Statistics in Biological Research, 3rd ed. W.H. Freeman, New York.
Sosa-López, A., D. Mouillot, J. Ramos-Miranda, D. Flores-Hernandez & T. D. Chi, 2007. Fish species richness decreases with salinity in tropical coastal lagoons. Journal of Biogeography 34: 52–61.
Souza, U. P., R. C. Costa, I. A. Martins & A. Fransozo, 2008. Associações entre as biomassas de peixes Sciaenidae (Teleostei: Perciformes) e de camarões Penaeoidea (Decapoda: Dendrobranchiata) no litoral norte do Estado de São Paulo. Biota Neotropica 8: 83–92. (in Portuguese).
ter Braak, C. J. F. & P. F. M. Verdonschot, 1995. Canonical correspondence analysis and related multivariate methods in aquatic ecology. Aquatic Sciences 57: 255–289.
Vega-Cendejas, M. & M. H. Santillana, 2004. Fish community structure and dynamics in a coastal hypersaline lagoon: Rio Lagartos, Yucatan, Mexico. Estuarine Coastal and Shelf Science 60: 285–299.
Vieira, J. P. & J. A. Musick, 1994. Fish faunal composition in warm-temperate and tropical estuaries of western Atlantic. Atlântica 16: 31–53.
Wentworth, C. R., 1922. A scale of grade and class terms of clastic sediments. Journal of Geology 3: 377–392.
Winemiller, K. O. & M. A. Leslie, 1992. Fish assemblages across a complex, tropical fresh-water marine ecotone. Environmental Biology of Fishes 34: 29–50.
Acknowledgments
We thank colleagues of the Ichthyology Laboratory of the Oceanographic Institute of the Federal University of Rio Grande (FURG), especially Vinícius Condini and Marcelo Burns, for their assistance in the field, and Kirk Winemiller and two anonymous reviewers for their comments, and the Coordenadoria de Aperfeiçoamento de Pessoal de Nível Superior CAPES that provided a student fellowship for PMM. The study received financial support of the Conselho Nacional de Desenvolvimento Científico e Tecnológico, CNPq (Research Grant No. 150868/2003-0) and from the Brazilian Long-Term Ecological Research (PELD—Phase I) program (sites 7 and 8).
Author information
Authors and Affiliations
Corresponding authors
Additional information
Handling editor: Odd Terje Sandlund
Rights and permissions
About this article
Cite this article
de Moura, P.M., Vieira, J.P. & Garcia, A.M. Fish abundance and species richness across an estuarine–freshwater ecosystem in the Neotropics. Hydrobiologia 696, 107–122 (2012). https://doi.org/10.1007/s10750-012-1187-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-012-1187-9