
Abstract
We investigated the abundance and taxonomic composition of the aquatic predatory insect fauna, with focus on adult diving beetles (Coleoptera: Dytiscidae), in eight temporary flooded wet meadows and two alder swamps in the River Dalälven floodplains, central Sweden from 2002 to 2006. Diving beetles are generalist predators and often abundant in various waters, including temporary wetlands. In the River Dalälven floodplains, recurrent floods induce massive hatching of flood-water mosquitoes (Diptera: Culicidae), which constitute a superabundant patchy and irregular food resource for aquatic predatory insects. Our aims were (1) to characterize the assemblage of adult diving beetles occurring in the wetlands during floods in relation to time and wetland type and (2) to evaluate the effect on the aquatic predator assemblage of strongly reducing the abundance of a potential prey, flood-water mosquito larvae with Bacillus thuringiensis var. israelensis (Bti) during floods. We found diving beetles to be the dominating aquatic predatory insect taxa in all 10 wetlands. There was a difference in Dytiscidae species richness but not in diversity between wet meadows and alder swamps after rarefaction. The cluster analysis based on dytiscid species and abundances showed very high similarities between the wetlands. The variance component analysis was unable to distinguish any factor that could explain more than 7.4% of the variation in the dytiscid species assemblages. The only effect of Bti-treatment against flood-water mosquito larvae, potential food for the predatory dytiscids, was a slight increase in abundance of the medium-sized dytiscid species. Our results are in accordance with previous studies, suggesting that irregular and recurrent flood dynamic structure the dytiscid fauna more than food limitations and environmental factors.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Recurrent but irregular floods along large rivers create temporary wetlands that provide opportunities for many species to proliferate and develop. Temporary flooded wetlands have long been neglected in studies of the aquatic insect fauna. Instead, the scientific focus has been on insect communities in permanent waters (Nilsson et al., 1994; Tate & Hershey, 2003), temporary pools (Kiflawi et al., 2003; Blaustein et al., 2004) and prairie pot holes (Anderson & Vondracek, 1999; Euliss et al., 2002). Ephemeral wetlands are, however, important for many aquatic and semi-aquatic species and newly flooded wetlands are often nutrient-rich and provide habitats for aquatic insects (Wiggins et al., 1980; Larson, 1985). Abundant food resources in temporary flooded wetlands may provide suitable feeding and reproduction possibilities for aquatic predatory insects (e.g. Coleoptera: Dytiscidae, Hemiptera: Corixidae and Notonectidae) (Batzer & Wissinger, 1996; Schneider, 1999; Wilcox, 2001). Flood duration is the most important factor structuring the aquatic insect fauna (Schneider & Frost, 1996). Long hydroperiods lead to a fauna more characteristic of permanent waters with higher degrees of biotic interactions including increased number of predators and density-dependent regulating factors (Wiggins et al., 1980; Wellborn et al., 1996; Jenkins & Buikema, 1998). Short hydroperiods are on the other hand, associated with species with fast developmental time, abundant food resources, less impact of predators and species adaptations to a fast changing environment (Nilsson, 1986; Wellborn et al., 1996; Williams, 1996). Flood-water mosquitoes (Diptera: Culicidae) are typical of short hydroperiod fauna, being highly adapted to temporary wetlands.
In addition to hydrology, the presence of predators may often constitute the greatest factor structuring invertebrate communities in wetlands (Wellborn et al., 1996). Temporary flooded wetlands are usually fishless and top predators are often dytiscids (Coleoptera: Dytiscidae) and other insects classified as generalist predators (Nilsson & Holmen, 1995; Batzer & Wissinger, 1996). Dytiscids occur in almost any type of freshwater aquatic environments, such as wetlands, snow pools, lakes, streams, rivers and bogs (Nilsson & Holmen, 1995). Dytiscids are generalist aquatic predators that will feed on any suitable aquatic prey, however, food preference seem to occur for at least some species (Deding, 1988; Kehl & Dettner, 2003; Lundkvist et al., 2003).
The floodplains of River Dalälven, central Sweden, are subjected to recurrent but irregular floods of variable amplitude and duration (http://www.vattenreglering.se). These recurrent floods have shaped a complex wetland landscape with a mixture of wet meadows, marshes and swamps. Although water flow regulations of River Dalälven have reduced the amplitude and duration of floods in most of the floodplains, the wetlands around Lake Färnebofjärden still have a semi-natural flood regime. Floods during May to August induce massive emergence of the flood-water mosquito Aedes sticticus, a species causing massive nuisance to humans in large areas around the wetlands (Schäfer et al., 2008). After decades of complaints, mosquito control operations were commenced in the summer of 2002, using the biological mosquito larvicide VectoBac G©, with protein crystals produced by Bacillus thuringiensis var. israelensis (Bti) as active ingredient to reduce populations of Aedes sticticus and other flood-water mosquito species. Bti is specific and efficient against mosquitoes (Becker et al., 2003) with few or no direct negative effects on non-target insects (Boisvert & Boisvert, 2000; Lundström et al., 2009). Flood-water mosquito larvae are abundant in the flooded areas from about 24 h after the flood reaches its maximum level, and for about 10 days depending on the water temperature.
Our aims were (1) to characterize the assemblage of adult diving beetles occurring in the wetlands during floods in relation to time and wetland type and (2) to evaluate the effect on the aquatic predator assemblage of strongly reducing the abundance of a potential prey, flood-water mosquito larvae (Diptera: Culicidae) with Bti during floods.
Materials and methods
Study areas
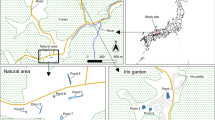
The aquatic predatory insect assemblages, especially adult dytiscid assemblages, were studied in eight open temporary wetlands (wet meadows) and two forested wetlands (alder swamps) around Lake Färnebofjärden in the River Dalälven floodplains, central Sweden during spring and summer floods from 2002 to 2006. Flood frequency, duration and magnitude have profound effects on the vegetation (Keddy, 2000). Wet meadows characterized by grasses and herbs with a few or no woody plants form in areas subjected to relatively frequent floods of relatively long duration. Swamps characterized by the abundance of woody plants and trees forming a canopy form in areas subject to less frequent floods of shorter duration. Floods in the River Dalälven floodplains are induced by snow-melt in spring and by heavy rain in summer. The geographical locations of the wetlands are given in Fig. 1 and the descriptions of individual wetland in Table 1.

Location of the River Dalälven floodplains in central Sweden and the 10 temporary wetlands where the aquatic predatory fauna was investigated from 2002 to 2006. In the experimental (Exp) wetlands, abundance of mosquito larvae was reduced by Bti-treatment, while no treatment was performed in reference (Ref) wetlands. The arrows indicate the flow direction of the River Dalälven
Reduction of flood-water mosquito larvae abundance in temporary flooded wetlands in the River Dalälven floodplains is based on aerial application of VectoBac G© granules. The aerial application of VectoBac G© at 13–15 kg ha−1 (0.4 kg Bti ha−1), using a helicopter sling system with a bucket spreader, gives close to 100% reduction of flood-water mosquito larvae (M. L. Schäfer and J. O. Lundström, unpublished). Four experimental wetlands (Laggarbo, Nordmyra, Norrån and Valmbäcken) were treated during periods of floods in 2002, 2003, 2005 and 2006 (Table 1). The remaining six wetlands (Fågle, Lusmyren, Gysinge, Ista, Stavnäs and Koversta) were reference wetlands, and untreated throughout the study.
Predatory insect sampling and identification
We used submerged activity traps to catch active aquatic predatory insects including adult Dytiscidae, Corixidae and Notonectidae (Jansson, 1996; Nilsson, 1996). The activity traps same as in Lundkvist et al. (2001) consisted of a 1.5 l plastic cylinder with a funnel (100 and 23 mm at the widest and narrowest points) inserted in the centre of the bottom, and a fine net lid (0.5 mm mesh) covering the other end to allow water exchange. Activity traps are commonly used in studies of dytiscids as they provide a relative clean sample (i.e. minimizing sorting time) and trap over night when many predators are active (Hilsenhoff, 1987; Hilsenhoff, 1991; Nilsson et al., 1994; Lundkvist et al., 2001). Each of the 10 wetlands was sampled for aquatic predatory insects during at least two consecutive years, in the 2002–2006 study period. On each sampling occasion, three to ten traps were placed in each wetland for 24, 48 or 72 h, and the water level at each trap position was measured. One trap used 24 h is one trapping period and two traps are two periods etc. The traps were usually placed horizontally on the bottom but occasionally also in mid water if the water depth exceeded 20 cm. The activity traps were used during floods and required a minimum water depth of 10 cm to remain submerged. Thus, the number of 24 h trapping periods varied between years and wetlands (Table 2). Upon emptying, the catch was sorted in the field and the predatory insects were transferred to plastic vials containing >70% ethanol.
Identification of the collected insects to family and genus followed the keys in Jansson (1996), Andersen (1996) and Solem and Gullefors (1996). Species identification of Dytiscidae followed the keys in Nilsson and Holmen (1995).
Statistical analyses
Statistical analyses were performed on Dytiscidae abundance and species data. We defined species richness as the number of dytiscid species found in each wetland and year. Rarefaction was calculated in EcoSim v7.68 (Gotelli & Entsminger, 2004) to compensate for differences in sampling effort between wetlands and years. Calculations of dytiscid diversity were made with Hurlbert’s PIE (Hurlbert, 1971). Hurlbert’s PIE diversity index gives the probability that two randomly sampled individuals from the assemblage represent two different species, and this index is relatively little influenced by sample size. The range of Hurlbert’s PIE is 0–1, where 0 represents low and 1 represents high diversity.
Rarefaction was calculated based on all dytiscid species, but to minimize the effect of variation in abundance of common species, we removed the most abundant species, Hydroporus striola, in 2002, 2003 and 2004. In total, 3,731 individuals of 59 species were used for the rarefaction analysis. We tested for differences in dytiscid species richness after rarefaction between wet meadows and alder swamps, and between experimental and reference wetlands from 2002 to 2003. Abundances were rarefied to 41 individuals from 2002 and 5 individuals from 2003, respectively. The rarefied number of species was analysed in relation to habitat and Bti-treatment in a MIXED MODEL (PROC MIXED, SAS Institute, 2004) with wetland as a random factor to account for the repeated sampling, while year was used as a fixed factor. Due to relative few individuals and species from 2005 and 2006, the cumulative number of dytiscid species richness was calculated for each individual wetland and used for testing of Bti-treatment effects between the experimental and reference wet meadows from 2005 to 2006. Abundances were rarefied to 20 individuals. This analysis was evaluated with ANOVA. The year 2004 was excluded from the analyses because no Bti-treatments were carried out.
Dytiscids are generalist aquatic predators feeding on the prey they can find, capture and handle. The body size range of Dytiscidae is from 2.0 to 44 mm in length, indicating a similar variation in size of suitable prey organisms (Deding, 1988). Individuals of Dytiscidae were grouped in three size classes irrespective of the species owing to species data was not sufficient (to few) for analyses at species level. The three dytiscid size classes were small (2–6.9 mm), medium-sized (7–15.9 mm) and large (≥16 mm), and followed the size distribution of dytiscid species in Nilsson & Holmen (1995). Moreover, species in all three size classes have been observed to be predators on mosquito larvae (Nilsson & Söderström, 1988; Nilsson & Holmen, 1995; Lundkvist et al., 2003). Number of individuals in all three size classes was tested for effects due to Bti-treatments between experimental and reference wetlands from 2002 to 2006. In addition, we tested what effect the interaction between dytiscid size classes and Bti-treatment had on dytiscid abundances between experimental and reference wetlands. The prerequisite was that the mosquito control with Bti caused a mosquito larvae reduction close to 100% (M. L. Schäfer and J. O. Lundström, unpublished). All tests were evaluated by ANCOVA, with water depth and time of year as covariates.
A cluster analysis based on McQuitty similarity analysis (SAS Institute, 2004) was performed and a dendogram constructed to visualize the similarities and dissimilarities in dytiscid assemblages between wetlands. The dytiscid assemblage data were controlled for variation in number of trapping periods between years before the dendogram was constructed. The McQuitty similarity analysis range from zero and upwards, depending on the similarities in the groups defined.
A variance component analysis was used to analyse the relative explanatory importance of each environmental and temporal variable, with respect to the variation in the weekly abundance of dytiscids in activity traps. The variance components included in the analysis were: dytiscid size class (the same three size classes that were used for the ANCOVA analyses), year of sampling, week of sampling, wetland, water depth at trap (range 10–50 cm) and Bti-treatment. The importance of each of the variance components were estimated in PROC VARCOMP using restricted maximum likelihood.
Statistical analyses were performed using the SAS statistical software, version 9.1 (SAS Institute, 2004).
Results
Composition of the aquatic predatory insect community
A total of 6,863 aquatic predatory insects were caught in activity traps in temporary flooded wetlands of the River Dalälven floodplains, during 2002–2006. The 1,707 trapping periods provided four insect orders with aquatic predatory insect species occurred: Coleoptera (3 families, 19 genera and 61 species), Odonata (5 families, 7 genera and 7 species), Hemiptera suborder Heteroptera (5 families, 10 genera and >20 species) and Trichoptera (1 family, 1 genera and 1 species) (Tables 3, 4).
Adult Dytiscidae had the largest number of individuals and species. Most common was H. striola which occurred in 0.2 to 7.7 individuals per 24 h trapping period including all wetlands and years. Only H. striola and Ilybius guttiger were recorded from all the 10 study wetlands. Small species, mainly Hydroporus spp. and Hygrotus spp., dominated the fauna from the wet meadows, while the medium-sized Ilybius subtilis dominated in the alder swamps. Large species comprised less than 1% of all dytiscids and were caught in very low numbers in all study wetlands during all years.
For the whole study period and all wetlands combined, the five most abundant small Dytiscidae species were H. striola (3,165 individuals), Hydroporus umbrosus (597), Hydroporus rufifrons (503), Hygrotus inaequalis (182) and Hydroporus palustris (172). The most abundant medium-sized species were I. guttiger (164), Ilybius ater (154), Rhantus exsoletus (149), Hydaticus aruspex (123), and Agabus congener (114). The most abundant large species were Dytiscus circumcinctus (44), Colymbetes striatus (9), Dytiscus marginalis (7) and Colymbetes paykulli (5). One species of particular protection concerns was recorded, viz Graphoderus bilineatus, (Natura 2000 species listed in the Council Directive 92/43/EEC on the Conservation of natural habitats and of wild fauna and flora). It was found in the experimental wet meadow Nordmyra 2003 and 2005 and in the reference wet meadows Gysinge 2004 and Ista 2005.
The highest abundances of dytiscids were observed during the spring floods, which usually occurred during May to June each year, by weeks 18 to 23 (Fig. 2). However, a deviation from this general temporal pattern occurred in 2002 when the dytiscid peak activity was recorded in August, by week 32. The year 2004 had the largest annual catch (71.4% of the total number of dytiscids). Two species, H. striola and H. umbrosus, dominated the dytiscid catches from 2003 to 2006 (Fig. 2a, c), while H. striola and I. ater dominated in 2002 (Fig. 2b).

Weekly dytiscid catches, with the two most abundant dytiscid species, collected with submerged activity traps in temporary wetlands in the River Dalälven floodplains in central Sweden. In a the annual catch of 2002, b the annual catch of 2004, and c the total catch of 2003, 2005 and 2006
Dytiscid richness and diversity
In total, 6,702 dytiscids including 61 species were collected in this study (Table 4). The highest number of species was caught in Fågle (reference wet meadow) and Laggarbo (experimental wet meadow), each with 36 species, and the lowest in Valmbäcken (experimental alder swamp) with 10 species. After rarefaction, Lusmyren (reference wet meadow) had the highest richness with 20 species, followed by Laggarbo with 18 species, and the lowest richness was found in Norrån (experimental wet meadow) with 3 species (Table 5).
To evaluate the sampling effort the cumulative number of Dytiscidae species was plotted against the cumulative number of 24 h trapping periods. The cumulated species curve asymptotically levelled out after 3 years of consecutive sampling and no additional species was added during the last 2 years of sampling (Fig. 3).

The cumulated number of Dytiscidae species plotted against the cumulated number of 24 h trapping periods, for the Dytiscidae sampling in 10 temporary wetlands in the River Dalälven floodplains central Sweden, 2002 to 2006
After rarefaction, we found differences in the dytiscid species richness between the wet meadows and alder swamps for the years 2002 to 2003 (P = 0.020, df = 1, F = 20.9, MIXED MODEL), and a significant year*wetland interaction (P = 0.032, df = 1, F = 14.3). There was higher species richness in the wet meadows than in the alder swamps in 2002, but not in 2003. We found no differences in dytiscid species richness between experimental wetlands and reference wetlands for the years 2002 and 2003 (P = 0.445, df = 1, F = 0.77, MIXED MODEL). In addition, there was no difference in the cumulative dytiscid richness after rarefaction between experimental and reference wet meadows for the years 2005 and 2006 (P = 0.615, df = 1, F = 0.29, ANOVA).
The Hurlbert’s PIE index of diversity varied between 0.35 and 0.90 for all wetlands and all years (Table 5). We found no differences in dytiscid diversity neither between years (P = 0.749, df = 4, F = 0.48, ANOVA), between wet meadows and alder swamps (P = 0.991, df = 1, F = 0, ANOVA), nor between experimental and reference wetlands (P = 0.392, df = 1, F = 0.75, ANOVA).
Dytiscid assemblage similarities between wetlands
The cluster analysis based on McQuitty similarity analysis (Fig. 4) of all wetlands and all years, based on dytiscid assemblages data, showed high similarity for 32 annual samples at a linking distance of 1.0, while Nordmyra 2005, Fågle 2004 and Stavnäs 2004 were solitaires. At a linking distance of 3.0, two clusters were formed; one small cluster containing Fågle 2004 and Stavnäs 2004, and one large cluster containing the remaining 33 annual samples. This clearly shows the high dytiscid species assemblage similarity between the study wetlands, both wet meadows and alder swamps.

McQuitty similarity analysis dendogram of the dytiscid species abundance composition for 10 temporary wetlands in the River Dalälven floodplains, central Sweden sampled from 2002 to 2006. Grey lines show the clusters formed at linking distances of 1.0 and 3.0
Variation in relation to temporal, environmental and biological factors
The variance component analysis showed that the measured factors together explained 20.6% of the variance in the dytiscid species assemblages. In more detail, the variance was explained by dytiscid size-class (7.4%), year (5.5%), week (3.5%), wetland (3.9%), water depth at trap (0.3%) and Bti-treatment (0.01%). Reduction of mosquito larvae abundance with Bti had no significant effect for any of the three dytiscid size classes (small P = 0.509, df = 1, F = 0.44, medium P = 0.468, df = 1, F = 0.53 and large P = 0.410, df = 1, F = 0.70, ANCOVA). However, the interaction between treatment and dytiscid size class showed significant pair-wise differences (P = 0.035) for medium-sized dytiscids. Medium-sized dytiscids were more abundant in experimental wetlands than in reference wetlands. There were no differences in the abundances of small-sized or large-sized dytiscids.
Discussion
We found 61 dytiscid species which is 68% of the number of dytiscids known from the regional species pool of the Swedish counties of Västmanland and Gästrikland (Nilsson & Holmen, 1995), in which this study was performed. In all wetlands studied, small dytiscids (<7 mm) were by far the most abundant, and medium-sized dytiscids occurred in smaller numbers, which is in accordance with results from similar studies in Alberta, Canada (Larson, 1985). We assume that the majority of the species that occur in wet meadows and swamps in the River Dalälven floodplains are included in this study, since no additional species were found during the last 2 years. Thus, our data on dytiscid species and abundance provide a good platform for characterizing the dytiscid assemblages and analysing the indirect effect on the predators of reducing the mosquito larvae abundance with Bti.
In our study, the number of dytiscid individuals varied greatly between years, with peak abundances in 2004. Even though 2004 was a dry year, there was an early spring flood in April, in which the majority of the dytiscids were collected. These large annual variations in dytiscid numbers will probably lead to variations in the potential dytiscid predation pressure, not only on mosquito larvae but on all potential prey co-occurring with dytiscids. As generalist predators, dytiscids may feed on all available prey taxa, and mosquito larvae will be the only prey among others, however, superabundant. Many dytiscids are known to include mosquito larvae in their diet (Deding, 1988; Nilsson & Söderström, 1988; Nilsson & Svensson, 1994; Lundkvist et al., 2003; Medlock & Snow, 2008), and a few studies show that dytiscids may lower the mosquito larvae abundance in lab and in the field (Lundkvist et al., 2003; Aditya et al., 2006; Chandra et al., 2008).
Nevertheless, it is not clear if dytiscids are also capable of naturally controlling mosquito populations. High predation pressure can be expected in small temporary ponds that form when the flood recedes, where “concentrations” of both predators and prey occur. Low predation pressure may on the other hand prevail during extensive floods when there instead will be a “dilution” effect of both predators and prey. In addition, water permanence may have a positive effect on the aquatic predator community, with an increased predation pressure with longer hydroperiod (Williams, 1997; Schneider, 1999; Schäfer et al., 2006). In most cases in the temporary wetlands of the River Dalälven floodplains, flood-water mosquito larvae will be a super-abundant food resource for a short period, and probably neither controlled by dytiscids nor any other predatory insect, as defined by Price (1984) for a pulsing resource type. In these short lived habitats, dytiscid larval development will probably be controlled by the short time available for growth instead of food limitations (Nilsson, 1986; Bellini et al., 2000).
There are many advantages for a dispersal behaviour, including inbreeding avoidance, competition avoidance and the ability to leave unfavourable conditions (Bilton et al., 2001). We found mainly spring-migrating dytiscid species in our study, which explains the abundance peaks found during spring flood. The most common dytiscid life cycle in north-western Europe is to over-winter as adult and with larvae developing in spring or summer and with the newly hatched adults in late summer or autumn (Nilsson & Holmen, 1995). Thus, the majority of the dytiscid species probably leave their overwintering sites early in spring to find suitable waters for reproduction, while the next generation of adult dytiscids leaves the water before the winter. Colonizing behaviour may be genera or species specific, with e.g. some Agabus spp. and Ilybius spp. re-colonizing the waters each spring (Nilsson & Svensson, 1995), and several Hydroporus spp. have been recorded flying around in the landscape in the spring colonizing new aquatic habitats in wetlands both in south-east Sweden (Lundkvist et al., 2002), and northern Delaware, USA (Fairchild et al., 2003). Other strategies of colonizing new aquatic habitats include following the flood, which eventually covers the wet meadows, or to break the reproductive diapause as found for Agabus disintegratus (Garcia & Hagen, 1987).
In a temporary wetland, alternating between aquatic and terrestrial conditions, the hydroperiod is a major force structuring the aquatic insect assemblages (Wiggins et al., 1980; Wellborn et al., 1996; Williams, 1996; Wissinger et al., 1996). Aquatic insects capable of flying could move between aquatic habitats, and Coleoptera can persist within seasonally flooded pools mainly due to re-colonization from adjacent permanent water bodies (Batzer & Resh, 1992). As an adaptation to a rapidly diminishing habitat, the dytiscid species that first colonize temporary waters in spring will leave for permanent water bodies when these temporary waters dry up later in the season (Batzer & Wissinger, 1996; Davy-Bowker, 2002). Permanent aquatic habitats are available within a few hundred meters from the wet meadows and swamps of the River Dalälven floodplains. Our findings are in agreement with the earlier studies that dytiscids due to good colonizing abilities are early inhabitants of temporary waters (Bellini et al., 2000; Davy-Bowker, 2002; Miguélez & Valladares, 2008), occur in large numbers and are relative species rich in Sweden as in other countries in northern areas.
We found high level of similarity for the dytiscid species assemblages in the wet meadows and alder swamps of the River Dalälven floodplains, with almost all wetlands forming one large cluster (Fig. 4). The higher species richness found in wet meadows than in alder swamps for 2002 may be an effect of shading, with migrating species reacting negatively to the increased shading in the alder swamps (Gee et al., 1997). The wet meadows in our study were deliberately chosen based on hydrological and biological similarities. The two alder swamps were partly chosen to diversify the habitat types in our study. Our results are in agreement with Lundkvist et al. (2001), who found environmentally similar wetlands containing similar dytiscid species compositions. The number of floods and flood duration varied between years and this had certainly a great effect on the observed dytiscid species assemblages. Other habitat conditions than water regime may have had less importance in structuring the dytiscid fauna as found in Lundkvist et al. (2001). Factors that positively influence dytiscid species richness are water permanence and absence of shading effects from trees (Nilsson & Svensson, 1994; Lundkvist et al., 2001; Rundle et al., 2002; Schäfer et al., 2006). Habitat complexity provided by vegetation may contribute positively to both the dytiscid species richness and diversity (Nilsson et al., 1994), and the most diverse dytiscid assemblages are found in small stagnant water bodies with at least some seasonality in the water fluctuations (Larson, 1985). Thus, the wetlands in our study relatively well resemble the habitats in which high species richness would be expected, even though few annual samples were from habitats with long hydroperiod.
We investigated the indirect effect of reducing the abundance of a potential prey, flood-water mosquito larvae, using the biological mosquito larvicide VectoBac G©. Dytiscids and some aquatic Hemiptera as generalist predators have been claimed as predators on mosquito larvae (Nilsson & Svensson, 1994; Blaustein, 1998; Lundkvist et al., 2003; Medlock & Snow, 2008), but no direct Bti-effect is known on these orders (Boisvert & Boisvert, 2000). In our analyses, Bti against mosquito larvae seemed to play only a minor role in structuring the dytiscid species assemblage, and the only significant result was a possible positive effect on the medium-sized dytiscids. The relatively long-term view (five consecutive years) of our study made it possible to also reveal less pronounced effects.
Even though mosquito larvae and dytiscids may occur simultaneously in high abundances our analyses could not find any clear relationship between them. Temporary wetlands will, however, be inhabited by many potential alternative preys including small crustaceans, aquatic insect larvae (mainly different Diptera larvae), plant material and algae all of which have been found in the diet of dytiscids (Deding, 1988; Kehl & Dettner, 2003). In fact, Deding (1988) found Chironomidae and small crustaceans to dominate as recognizable food items for the majority of the dytiscid species studied. Lundkvist et al. (2003) found different prey preference of three medium-sized dytiscid species on two alternative preys (mosquito larvae and Daphnia) in laboratory tests. Flood-water mosquito larvae may constitute an example of an unpredictable super-abundant food resource that varies strongly within and between years. Thus, flood-water mosquito larvae are probably preyed upon by dytiscids during floods with massive mosquito hatching, while in times of lower mosquito production, alternative prey are fed upon.
We are aware that catches from activity traps may not always produce representative data for all aquatic insect groups (Turner & Trexler, 1997). The traps are catching insects that actively move in the water and recordings of more sluggish and immobile insects may be under represented. However, in the temporary wetlands of River Dalälven, the hydroperiod is often short and occurs on an irregular basis, and this certainly affects the aquatic insect species composition. Dragonflies and caddis flies are rarely collected in activity traps (Elmberg et al., 1992). However, both dragonflies and caddis flies occur in aquatic habitats with longer hydroperiods than the short lived wetlands in this study. Corixidae may be under represented in activity traps placed in mid water, since they usually swim close to the bottom (Elmberg et al., 1992; Hyvönen & Nummi, 2000). In addition, Elmberg et al. (1992) found that both bottom and mid-water activity trap positions were equally good when investigating mobile aquatic invertebrate fauna, but mid-water traps may generate a more balanced material in respect of nektonic and benthic organisms (Hyvönen & Nummi, 2000). However, our aim was to characterize the dominant aquatic predatory fauna, and compare wetlands and treatments and therefore the use of activity traps was a relevant method.
Conclusion
Diving beetles dominated the aquatic predatory insect fauna in the temporary flooded wetlands of the River Dalälven floodplains, and peak activity usually occurred in connection to the spring flood. A total of 61 diving beetle species, 68% of the regional species pool, and 10–36 species per wetland show a rather diverse diving beetle fauna. Species richness but not diversity after rarefaction differed significantly between wetland types and years. However, the cluster analysis based on McQuitty similarity analysis showed one major cluster including almost all wetlands during all years, and one minor cluster indicating high dytiscid assemblage similarities. Abundance of flood-water mosquito larvae, a superabundant potential prey for diving beetles in flooded wetlands, was strongly reduced by Bti-treatment in experimental wetlands, while no treatment was performed in reference wetlands. Although the abundance of flood-water mosquito larvae was reduced by close to 100%, there was no general significant effect on diving beetle abundance. Our results indicate that hydrology is the most important factor shaping the diving beetle assemblages of temporary flooded wetland environments, while variation in the abundance of a dominant prey taxon had no major influence.
References
Aditya, G., A. Ash & G. K. Saha, 2006. Predatory activity of Rhantus sikkimensis and larvae of Toxorhynchites splendens on mosquito larvae in Darjeeling, India. Journal of Vector Borne Diseases 43: 66–72.
Andersen, N. M., 1996. Heteroptera Gerromorpha, semiaquatic bugs. In Nilsson, A. (ed.), Aquatic Insects of North Europe—A Taxonomic Handbook, Vol. 1. Apollo Books Aps, Stenstrup: 77–90.
Anderson, D. J. & B. Vondracek, 1999. Insects as indicators of land use in three ecoregions in the prairie pothole region. Wetlands 19: 648.
Batzer, D. P. & V. H. Resh, 1992. Macroinvertebrates of a California seasonal wetland and responses to experimental habitat manipulation. Wetlands 12: 1–7.
Batzer, D. P. & S. A. Wissinger, 1996. Ecology of insect communities in nontidal wetlands. Annual Review of Entomology 41: 75–100.
Becker, N., M. Zgomba, D. Petrić, C. Dahl, C. Boase, J. Lane & A. Kaiser, 2003. Mosquitoes and Their Control. Kluwer Academic/Plenum Publishers, New York.
Bellini, R., F. Pederzani, R. Pilani, R. Veronesi & S. Maini, 2000. Hydroglyphus pusillus (Fabricius) (Coleoptera Dytiscidae): its role as a mosquito larvae predator in rice fields. Bollettino dell’Istituto di Entomologia ‘G. Grandi’ dell’Università di Bologna 54: 155–163.
Bilton, D. T., J. R. Freeland & B. Okamura, 2001. Dispersal in freshwater invertebrates. Annual Review of Ecology and Systematics 32: 159–181.
Blaustein, L., K. Moshe, A. Eitam, M. Marc & J. E. Cohen, 2004. Oviposition habitat selection in response to risk of predation in temporary pools: mode of detection and consistency across experimental venue. Oecologia 138: 300–305.
Boisvert, M. & J. Boisvert, 2000. Effects of Bacillus thuringiensis var. israelensis on target and nontarget organisms: a review of laboratory and field experiments. Biocontrol Science and Technology 10: 517–561.
Chandra, G., S. K. Mandal, A. K. Ghosh, D. Das, S. S. Banerjee & S. Chakraborty, 2008. Biocontrol of larval mosquitoes by Acilius sulcatus (Coleoptera: Dytiscidae). BMC Infectious Diseases 8:138.
Davy-Bowker, J., 2002. A mark and recapture study of water beetles (Coleoptera: Dytiscidae) in a group of semi-permanent and temporary ponds. Aquatic Ecology 36: 435–446.
Deding, J., 1988. Gut content analysis of diving beetles (Coleoptera: Dytiscidae). Natura Jutlandica 22: 177–184.
Elmberg, J., P. Nummi, H. Pöysä & K. Sjöberg, 1992. Do intruding predators and trap position affect the reliability of catches in activity traps? Hydrobiologia 239: 187–193.
Euliss, N. H. J., D. M. Mushet & D. H. Johnson, 2002. Using aquatic invertebrates to delineate seasonal and temporary wetlands in the prairie pothole region of North America. Wetlands 22: 256–262.
Fairchild, G. W., J. Cruz, A. M. Faulds, A. E. Z. Short & J. F. Matta, 2003. Microhabitat and landscape influences on aquatic beetle assemblages in a cluster of temporary and permanent ponds. Journal of the North American Benthological Society 22: 224–240.
Garcia, R. & K. S. Hagen, 1987. Summer dormancy in adult Agabus disintegratus (Crotch) (Coleoptera: Dytiscidae) in dried ponds in California. Annales of the Entomological Society of America 80: 267–271.
Gee, J. H. R., B. D. Smith, K. M. Lee & S. W. Griffiths, 1997. The ecological basis of freshwater pond management for biodiversity. Aquatic Conservation: Marine and Freshwater Ecosystems 7: 91–104.
Gotelli, N. J. & G. L. Entsminger, 2004. EcoSim: Null Models Software for Ecology. Version 7. Acquired Intelligence Inc. & Kesey-Bear, Jericho.
Hilsenhoff, W. L., 1987. Effectiveness of bottle traps for collecting Dytiscidae (Coleoptera). The Coleopterists Bulletin 41: 377–380.
Hilsenhoff, W. L., 1991. Comparison of bottle traps with a D-frame net for collecting adults and larvae of Dytiscidae and Hydrophilidae (Coleoptera). The Coleopterists Bulletin 45: 143–146.
Hurlbert, S., 1971. The nonconcept of diversity: a critique and alternative parameters. Ecology 52: 577–586.
Hyvönen, T. & P. Nummi, 2000. Activity traps and the corer: complementary methods for sampling aquatic invertebrates. Hydrobiologia 432: 121–125.
Jansson, A., 1996. Heteroptera Nepomorpha, aquatic bugs. In Nilsson, A. (ed.), Aquatic insects of North Europe—A Taxonomic Handbook, Vol. 2. Apollo Books Aps, Stenstrup: 91–104.
Jenkins, D. G. & A. L. J. Buikema, 1998. Do similar communities develop in similar sites? A test with zooplankton structure and function. Ecological Monographs 68: 421–443.
Keddy, P. A., 2000. Wetland Ecology—Principles and Conservation. Cambridge University Press, Cambridge.
Kehl, S. & K. Dettner, 2003. Predation by pioneer water beetles (Coleoptera, Dytiscidae) from sandpit ponds, based on crop-content analysis and laboratory experiments. Archiv fuer Hydrobiologie 158: 109–126.
Kiflawi, M., A. Eitam & L. Blaustein, 2003. The relative impact of local and regional processes on macro-invertebrate species richness in temporary pools. Journal of Animal Ecology 72: 447–452.
Larson, D. J., 1985. Structure in temperate predaceous diving beetle communities (Coleoptera: Dytiscidae). Ecography 8: 18–32.
Lundkvist, E., J. Landin & P. Milberg, 2001. Diving beetle (Dytiscidae) assemblages along environmental gradients in an agricultural landscape in southeastern Sweden. Wetlands 21: 48–58.
Lundkvist, E., J. Landin & F. Karlsson, 2002. Dispersing diving beetles (Dytiscidae) in agricultural and urban landscapes in south-eastern Sweden. Annales Zoologici Fennici 39: 109–123.
Lundkvist, E., J. Landin, M. Jackson & C. Svensson, 2003. Diving beetles (Dytiscidae) as predators of mosquito larvae (Culicidae) in field experiments and in laboratory tests of prey preference. Bulletin of Entomological Research 23: 219–226.
Lundström, J. O., M. L. Schäfer, E. Petersson, T. Z. P. Vinnersten, J. Landin & Y. Brodin, 2009. Production of wetland Chironomidae (Diptera) and the effects of using Bacillus thuringiensis israelensis for mosquito control. Bulletin of Entomological Research Jun 5: 1–9. doi:10.1017/S0007485309990137.
Medlock, J. M. & K. R. Snow, 2008. Natural predators and parasites of British mosquitoes—a review. European Mosquito Bulletin 25: 1–11.
Miguélez, D. & L. F. Valladares, 2008. Seasonal dispersal of water beetles (Coleoptera) in an agricultural landscape: a study using Moericke traps in northwest Spain. Annales de la Société Entomologique de France : revue d'entomologie générale et appliquée 44: 317–326.
Nilsson, A. N., 1986. Community structure in the Dytiscidae (Coleoptera) of a northern Swedish seasonal pond. Annales Zoologici Fennici 23: 39–47.
Nilsson, A. N., 1996. Coleoptera, introduction. In Nilsson, A. (ed.), Aquatic Insects of North Europe—A Taxonomic Handbook, Vol. 1. Apollo Books Aps, Stenstrup: 115–122.
Nilsson, A. N. & M. Holmen, 1995. The aquatic Adephaga (Coleoptera) of Fennoscandia and Denmark. II. Dytiscidae. E. J. Brill, Leiden.
Nilsson, A. N. & O. Söderström, 1988. Larval consumption rates, interspecific predation, and local guild composition of egg-overwintering Agabus (Coleoptera, Dytiscidae) species in vernal ponds. Oecologia 76: 131–137.
Nilsson, A. N. & B. W. Svensson, 1994. Dytiscid predators and culicid prey in two boreal snowmelt pools differing in temperature and duration. Annales Zoologici Fennici 31: 365–376.
Nilsson, A. N. & B. W. Svensson, 1995. Assemblages of dytiscid predators and culicid prey in relation to environmental factors in natural and clear-cut boreal swamp forest pools. Hydrobiologia 308: 183–196.
Nilsson, A. N., J. Elmberg & K. Sjoberg, 1994. Abundance and species richness patterns of predaceous diving beetles (Coleoptera, Dytiscidae) in Swedish lakes. Journal of Biogeography 21: 197–206.
Price, P. W., 1984. Alternative paradigms in community ecology. In Price, P. W., C. N. Slobodchikoff & S. Gaud (eds), A New Ecology. Novel Approcahes to Interactive Systems. Wiley, New York: 353–383.
Rundle, S. D., A. Foggo, V. Choiseul & D. T. Bilton, 2002. Are distribution patterns linked to dispersal mechanism? An investigation using pond invertebrate assemblages. Freshwater Biology 47: 1571–1581.
SAS Institute, 2004. SAS version 9.1, Statistical Software. Cary, NC, USA.
Schäfer, M. L., E. Lundkvist, J. Landin, T. Z. Persson & J. O. Lundström, 2006. Influence of landscape structure on mosquitoes (Diptera: Culicidae) and dytiscids (Coleoptera: Dytiscidae) at five spatial scales in Swedish wetlands. Wetlands 26: 57–68.
Schäfer, M. L., J. O. Lundström & E. Petersson, 2008. Comparison of mosquito (Diptera: Culicidae) populations by wetland type and year in the lower River Dalälven region, Central Sweden. Journal of Vector Ecology 33: 150–157.
Schneider, D. W., 1999. Influence of hydroperiod on invertebrate community structure. In Batzer, D. P., R. B. Rader & S. A. Wissinger (eds), Invertebrates in Freshwater Wetlands of North America. Wiley, New York: 299–318.
Schneider, D. W. & T. M. Frost, 1996. Habitat duration and community structure in temporary ponds. Journal of the North American Benthological Society 15: 64–86.
Solem, J. O. & B. Gullefors, 1996. Trichoptera, caddisflies. In Nilsson, A. (ed.), Aquatic Insects of North Europe—A Taxonomic Handbook, Vol. 1. Apollo Books Aps, Stenstrup: 223–255.
Tate, A. W. & A. E. Hershey, 2003. Selective feeding by larval dytiscids (Coleoptera: Dytiscidae) and effects of fish predation on upper littoral zone macroinvertebrate communities of arctic lakes. Hydrobiologia 497: 13–23.
Turner, A. M. & J. C. Trexler, 1997. Sampling aquatic invertebrates from marshes: evaluating the options. Journal of the North American Benthological Society 16: 694–709.
Wellborn, G. A., D. K. Skelly & E. E. Werner, 1996. Mechanisms creating community structure across a freshwater habitat gradient. Annual Review of Ecology and Systematics 27: 337–363.
Wiggins, G. B., R. J. Mackay & I. M. Smith, 1980. Evolutionary and ecological strategies of animals in annual temporary pools. Archiv für Hydrobiologie Suppl 58: 97–206.
Wilcox, C., 2001. Habitat size and isolation affect colonization of seasonal wetlands by predatory aquatic insects. Israel Journal of Zoology 47: 459–475.
Williams, D. D., 1996. Environmental constraints in temporary fresh waters and their consequences for the insect fauna. Journal of the North American Benthological Society 15: 634–650.
Williams, D. D., 1997. Temporary ponds and their invertebrate communities. Aquatic Conservation: Marine and Freshwater Ecosystems 7: 105–117.
Wissinger, S. A., G. B. Sparks, G. L. Rouse, W. S. Brown & H. Steltzer, 1996. Intraguild predation and cannibalism among larvae of detritivorous caddisflies in subalpine wetlands. Ecology 77: 2421–2430.
Acknowledgements
We appreciated the grants from the Swedish Environmental Protection Agency to J.O.L, and the invaluable help from Martina Schäfer and Yngve Brodin for comments on the manuscript, Örjan Östman for discussions about statistics and rarefaction analyses, Anna Hagelin, Björn Forsberg, Gustav Carlander, Anna-Sara Liman assisted with field sampling and identification, and Gunnar Sjödin with identification of dytiscid species.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: B. Oertli
Rights and permissions
About this article
Cite this article
Vinnersten, T.Z.P., Lundström, J.O., Petersson, E. et al. Diving beetle assemblages of flooded wetlands in relation to time, wetland type and Bti-based mosquito control. Hydrobiologia 635, 189–203 (2009). https://doi.org/10.1007/s10750-009-9911-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-009-9911-9