
Abstract
In Mediterranean climate areas, the great seasonal variability in temperature and rainfall is considered to be an evolutionary pressure that constrains plant and animal communities and their biological traits. Droughts alter habitat availability (changes of flow alter riffle-pool sequences), although habitat characteristics may also exacerbate drought to some extent. Using a simple quantitative index based on the proportion of conglomerate bedrock versus gravel and cobbles, pools versus riffles and winter versus summer flow, we show how habitat characteristics (in terms of substratum and flow) may influence the permanency of a stream site and how flow permanence constrains macroinvertebrate community structure and biological traits. Annual and seasonal macroinvertebrate richness, and the EPT (Ephemeroptera, Plecoptera and Trichoptera) and OCH (Odonata, Coleoptera and Hemiptera) metrics differed between permanent, intermittent and ephemeral sites, but not between permanent and intermittent sites. In contrast, distinct biological traits were observed in the three flow categories, although permanent sites presented few significant traits which was attributed to the stability of the habitat. Intermittent sites were dominated by taxa with pool-like strategies, while ephemeral sites were characterized by fauna with life-history adaptations to floods and droughts. In contrast to most traits (e.g., dissemination, reproduction, substrate relation), which were more constrained by local flow and substrate characteristics, life-cycle characteristics did not differ significantly among flow categories. This pattern can be explained by the features of the Mediterranean climate and particularly its high seasonal predictability, which serves as a large-scale filter of life-cycle traits, independently of local hydromorphological characteristics. Our findings indicate that drought is related to habitat characteristics and that local habitat variability favours organisms with certain traits, while other traits are independent of habitat variability on this scale and are probably affected by other large-scale habitat characteristics.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Several scale-dependent environmental factors are responsible for spatial and temporal heterogeneity in ecosystems and, at the same time, constrain the distribution patterns and responses of organisms (Milne, 1991; Poff, 1997). The relationships between habitat heterogeneity and organisms’ response are summarized in the Habitat Templet Theory (Southwood, 1977; 1988). This proposes that habitat and its spatio-temporal variability is the framework against which evolution selects organisms with characteristic life-history strategies and, simultaneously, determines the presence and abundance of these organisms at distinct scales. Over the last decade, this theory has been applied to both terrestrial and aquatic ecosystems and has shown matches and mismatches between habitat characteristics and the biological traits of organisms (e.g., Statzner et al., 1994; Ribera et al., 2001; Thuiller et al., 2004). Applied to river ecosystems, examples from the Habitat Templet Theory (River Habitat Templet by Townsend & Hildrew, 1994—RHT) include a wide array of aquatic organisms, from fungi to amphibians and fish (e.g., Joly & Morand, 1994; Muotka & Virtaten, 1995; Pattee & Chergui, 1995; Biggs et al., 1998; Townsend et al., 1997; Lamouroux et al., 2002; 2004; Poff et al., 2006).
Thus, the association between habitats and the traits of their matched species can be established in two-dimensional space (spatial and temporal heterogeneity). Commonly, these two dimensions are associated with disturbance (Poff & Ward, 1990; Townsend & Hildrew, 1994). The rationale behind this association is that natural disturbances are large sources of heterogeneity in ecosystems because they alter the structure of the environment and the distribution of organisms (White & Harrod, 1997; but see Wiens, 2000). In stable environments, specialist species are favoured while in more unstable conditions generalist strategies dominate (Southwood et al., 1974; Southwood, 1988; Poff & Allan, 1995). In Mediterranean climatic areas around the world, seasonal variations in temperature and rainfall patterns are considered to be evolutionary pressures that constrain plant and animal communities and their biological traits (di Castri, 1981; Stamou, 1998). Rivers in these areas show seasonal and predictable disturbances such as floods and droughts (McElravy et al., 1989; Gasith & Resh, 1999) and three different general flow categories can be distinguished: permanent (flowing waters), intermittent (isolated pools during the dry season that may persist to the end of summer) or ephemeral (dry stream beds in the dry season or even for longer periods). Overall, we could consider that these three flow categories differ in their spatio-temporal characteristics, and this framework makes our system very suitable for assessing the predictions of the RHT: temporal heterogeneity decreases along a gradient from ephemeral over intermittent to permanent because flow stability increases. Conversely, spatial heterogeneity may increase along this gradient because water permanency may tend to be associated with higher water levels and coverage of land, representing habitats that are structurally more diverse. While several studies have described the way in which droughts alter habitat availability (Boulton & Lake, 1992b; Stanley et al., 1997; Lake, 2000; Boulton, 2003), few report how habitat may exacerbate drought (but see Smith et al., 2003).
The responses of macroinvertebrate communities to flow permanence have been widely reported. In many cases, differences in the faunal composition of permanent and intermittent sites are slight and there is a high overlap between assemblages (Boulton & Suter, 1986; Delucchi, 1988; Delucchi & Peckarsky, 1989; Boulton & Lake, 1992a; Feminella, 1996; Williams, 1996; del Rosario & Resh, 2000). While taxonomic richness and composition are the features most studied, research on biological traits under a range of flow conditions are required to elucidate the environmental constraints that determine the characteristics of organisms (Williams, 1991, 1996) and few studies exist on this topic. Moreover, the use of biological traits to characterize environmental conditions has advantages because traits are independent of taxonomic constraints (Statzner et al., 1997), which may allow global predictions of organisms’ responses to flow disturbance.
Flow interacts with substrate to create dynamic patterns in, for example, the documented movement and deposition of fines even at base flow (Wood & Armitage, 1997). In this study, we checked how habitat characteristics, such as substrate and water flow determine the extent of permanence in a Mediterranean river network. A site is supposed to be less temporary if it is dominated by bedrock, large pools and high winter and summer flows at the time of sampling. In contrast, a site with a habitat dominated by gravel or cobbles, riffles and low winter and summer flows is expected to dry at any time during that summer. We expected that these habitat characteristics are associated with macroinvertebrates of sites and we wanted to test the prediction that permanent streams are dominated by organisms with traits adapted to dominant riffle conditions (i.e., riffles and pools are always present), that intermittent streams are mainly inhabited by organisms with traits adapted to dominant pool conditions (i.e., riffles are lost during summer), whereas ephemeral streams mainly contain communities of species that have traits which make them resistant or resilient to the absence of water during the dry period.
Methods
Study area
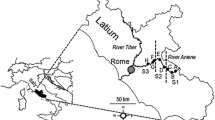
The study was carried out in the Sant Llorenç del Munt Natural Park (Catalonia, NE Spain) (Fig. 1). This park, located north of the metropolitan area of Barcelona (NE Spain) and occupying 9,630 Ha, has benefited from conservation practices for 20 years. The mountain ranges in which the park is located have a typical Mediterranean climate, with irregular and intense rainfall, mostly in winter, but with some rainfall in spring and autumn, while summer is normally very dry. The park has a dominant karstic geology with highly permeable substrates and, therefore, surface flow in streams may cease in hours or days after rainfall. However, the area also has several permanent streams, mostly linked to springs that discharge from the karstic aquifer (the most permanent ones are situated at low altitudes). Vegetation is dominated by evergreen oak trees (Quercus ilex L.) and white pines (Pinus halepensis Miller), except in the steepest areas or in places with rock outcrops. Altitude ranges from 280 m to 1,100 m, and 20% of the reaches studied extend beyond the limits of the park. The streams sampled belong to two main catchments that discharge to the Mediterranean, the Besòs and Llobregat river catchments. The Besòs catchment drains from three Natural Parks (Sant Llorenç del Munt, Montseny and Corredor-Montnegre) and has a surface of 1,007 km2. The Llobregat catchment drains from the Pre-Pyrenees and has a surface of 4,923 km2. More information of these two catchments can be found in Prat et al. (1984), Prat & Ward (1994) and Prat et al. (2000).

Sampling area and sites (white circles), indicating channel status in winter and summer period. Solid circles indicate dry sites
Habitat characteristics
The length of the river network in the park was measured on a 1:50000 scale map that defined the main channels. This network was examined in detail in winter (February) and summer (August) 1996, exploring all streams and springs that were close to the main and minor routes and, in some cases, far form these. Overall, we visited 78 sites, resulting in almost one site per tributary and up to 2.7 sites visited per km of stream length (with an average of 1.5). These 78 sites were characterized in winter and summer period according to three different flow categories: permanent (flow > 1 l s−1), intermittent (only pools) or ephemeral (dry). From them, only 25 sites, covering an altitudinal range of 280–565 m a.s.l., had flow in winter and were chosen for this study (Fig. 1). At each of these 25 sites, winter and summer flow was measured. Habitat structure was recorded in winter by estimating the percentage of gravel, cobbles, or bedrock and the percentage of pools versus riffles in a 50 m reach. All sampled sites could be considered as reference because no man-made impacts are affecting the area (Rieradevall et al., 1999).
The study was performed in 1996, a very wet year (1,144 mm of annual rainfall) that was preceded by several years in which the mean annual rainfall had been lower (610–884 mm between 1992 and 1995). Rainfall in 1996 fell mainly in January and during the autumn, while spring presented medium to low levels. Consequently, samples from winter and summer were collected after a period of high and low precipitation, respectively.
According to our assumptions, a stream site should be more permanent in the following circumstances: (1) over large conglomerate bedrock (no infiltration) versus sites where gravel or cobbles dominate, (2) when large pools are present (more water accumulated), (3) when high flow is present in winter (delayed drought) and (4) when high flow is present in summer at the time of sampling (less probability of drying at any time during that summer). Thus, we quantified these site attributes and estimated a site permanence score (SPS) as follows, and in a similar way to that of Feminella (1996). We first ranked the 25 sites in function of the importance of four attributes separately (See Electronic Supplementary Material): (1) the percentage of bedrock; (2) the percentage of pools; (3) the relative winter flow (winter flow at site relative to the maximum winter flow measured in all sites); and (4) the relative summer flow (summer flow at site relative to the maximum summer flow measured in all sites). As we used 25 sites, for each parameter, the first site with the highest percentage for this parameter received a maximum score of 25, the next 24 and successively, until the last site received a score of 1. For each parameter, percentages were grouped in classes (i.e., 0.1–9%, 10–19%, up to 100%). When several sites had the same percentage for one of the parameters, the mean of their rank scores was assigned to each one (e.g., if three sites had 90% hard substrata and their rank was between 24 and 22, a score of 23 was given to all three sites). After ranking the sites, the four scores were added for each site and the final SPS was obtained, which could potentially vary between 100 (highest possible value if a site ranked first in all attributes) and 4 (the lower value possible if a site had the lowest and single value in all attributes). Thus, we expected permanent conditions for high SPS values (mostly pools over hard substrata and maximum flow recorded both in winter and summer), intermittent conditions for intermediate SPS values and ephemeral conditions for low SPS values (mostly riffles over gravel or cobbles, low winter flow and low or null summer flow). To determine how well this SPS classify sites into permanent, intermittent and ephemeral conditions, we divided the obtained SPS in three groups using a k-means method (Legendre & Legendre, 1998), which maximizes the differences among groups of sites and minimizes the differences among sites within a group. The resulting groups of sites were compared with the observed flow category in summer and the error of classification (e.g., number of observed intermittent sites classified in the predicted permanent group) was calculated. The SPSS statistical package (version 10, SPSS Inc., Chicago, US) was used for this analysis.
Taxonomic composition and biological traits of macroinvertebrates
Macroinvertebrate samples were taken with a 250 μm mesh size net, using the kick method in all available substrates proportionally and with a similar effort at each site. The samples were preserved in formalin, sorted and identified at genus level. In some cases, several taxa were kept at family level because of difficulty in identification (small organisms or with complex taxonomy).
Taxon richness was compared between the groups of sites with distinct flow regimes and seasons using Kruskal-Wallis non-parametric ANOVA tests as the values differed from normality. The STATISTICA Program was used to perform the analyses (version 1999, Stat Soft Inc., Tulsa, UK). The same analyses were applied to EPT (sum of number of Ephemeroptera, Plecoptera and Trichoptera taxa) and OCH (sum of number of Odonata, Coleoptera and Hemiptera taxa). These two metrics are good indicators of the seasonal changes in macroinvertebrate communities in Mediterranean rivers (Rieradevall et al., 1999; Bonada et al., 2006).
The biological characteristics of macroinvertebrate taxa were obtained from Usseglio-Polatera et al. (2000) and include 11 traits with a total of 63 modalities. These traits are related to life-cycle features (maximum size, life-cycle duration, potential number of reproduction cycles per year, aquatic stages), resistance or resilience (dispersal, substrate relation, resistance forms), physiology and morphology (respiration, locomotion) and feeding and reproduction behaviour (reproduction, food and feeding habits). Several of these studies that have focused on traits report a mismatch between habitat characteristics and biological traits as predicted by the RHT (e.g., Usseglio-Polatera, 1994; Persat et al., 1994). Methodological problems related to trade-offs between traits, the scale of study, or the biological data used, may explain this non-conformity with theoretical assumptions (Resh et al., 1994). To overcome this problem, several authors have proposed testing habitat-trait theories using groups of organisms with similar characteristics (Statzner et al., 1997; Usseglio-Polatera et al., 2000). Thus, for example, Usseglio-Polatera et al. (2000) assigned each taxon to a group and subgroup, both of which include taxa that share similar traits. To build the trait matrix with our data, each taxon was checked for the Usseglio-Polatera’s subgroups (or for group when subgroup was not available) and the mean distribution frequency of the affinities of each subgroup for each modality of the 11 traits was used (Usseglio-Polatera et al., 2000, See Electronic Supplementary Material; Usseglio-Polatera, unpubl. data). For example, Mesovelia belonged to the g2 subgroup (Usseglio-Polatera et al., 2000) and had a mean frequency of affinity of 0.433 for life-cycle duration ≤ 1 year and of 0.567 for life duration > 1 year (Usseglio-Polatera, unpubl. data). For some taxa, information on biological traits was not available and these were excluded from the analysis (e.g., Hydracarina). When identification to genus was not possible, the biological traits of the family were used according to the group in which most of the genera of that family were assigned. That was the case of, for example, Corduliidae or Coenagrionidae.
The “4th Corner method” (Legendre et al., 1997) was used to check for differences in biological traits between the different flow conditions. This method, which is similar to the RLQ proposed by Dolédec et al. (1996), uses a taxon distribution (taxa vs. sites), a behavioural (taxa vs. traits) and an environmental (sites vs. environment or habitat) matrix to develop a new matrix that relates the distinct kinds of habitat to biological traits (i.e., a habitat vs. traits matrix). In our case, the taxon distribution matrix was the annual macroinvertebrate matrix (winter and summer matrices) transformed to presence/absence because of requirements of the program (sites × taxa: 25 × 68). The behavioural matrix included the degree of affinity of each taxon to each trait category (trait categories × taxa: 63 × 68); and the environmental-habitat matrix was a multi-state nominal variable recoded as three dummy variables: permanent, intermittent or ephemeral (sites × habitat: 25 × 3). Firstly, to check if there was global association between behavioural and habitat matrices (i.e., if significant differences were found between all habitats and a behavioural condition), an analysis of variance statistic (i.e., the global F-statistic) was calculated with the program for each trait category. If F is significant, at least one habitat differed from the others in terms of each trait category. Furthermore, to explore the specific relationship between the two matrices (i.e., if significant differences were found between a specific habitat and a behavioural condition), the statistics used in the “4th Corner method” depend on the type of variables. When using a quantitative behavioural matrix and a qualitative habitat matrix coded into dummy variables, as in this study, Legendre et al. (1997) provided two options: the use of the homogeneity statistic (d) or the Pearson product-moment correlation coefficient (r). We used the latter to obtain the strength and direction of association between the two matrices. The r-value indicates a positive or negative correlation between each trait category and each flow category (permanent-intermittent-ephemeral). The significance of r and F was obtained by random permutations (999 runs) and included Holm’s correction to avoid Type I error. From all the permutation models available in the “4th Corner method”, Model-1 (environmental control over individual species) was used because we assumed that a species is found in a particular habitat when environmental conditions are appropriate for its development (Legendre et al., 1997).
Results
Flow permanence
The flow conditions of the drainage network of the Sant Llorenç del Munt Natural Park for winter and summer are shown in Fig. 1. Three conditions observed during each sampling period are illustrated: (1) Flowing water (from 1 l s−1 to 600 l s−1) (continuous line), (2) River courses with pools but without surface flow (dashed lines) and (3) Dry watercourses (dotted line). These three flow categories represent permanent, intermittent and ephemeral streams, respectively. In winter, 63% of the total network length had continuous flow, while in summer this flow regime accounted for only 26%. The rest of the river length was intermittent (only pools) or ephemeral (dry). The main streams with permanent flow in summer were outside or at the limits of the park area, downstream of permanent springs. Although 1996 was a relatively wet year, 8 of the 25 sites sampled in February were totally dry in summer.
The 25 sampling sites were ranked using the method described above (SPS). The final SPS (the sum of the four values for each sampling site, see Methods) ranged from 16.5 to 84 (Table 1). Using the SPS values, k-means method classified sites as follows: the first 7 with high SPS (i.e., presumably permanent), the next 11 with intermediate SPS values (i.e., presumably intermittent) and the last 7, with low SPS values (i.e., presumably ephemeral). Overall, when comparing predicted SPS to the observed condition, we obtained an 84% of classification success, with the permanent and ephemeral conditions being the best classified, indicating that attributes used in SPS influence site flow permanence. Although our method could be also sensitive to error, misclassified sites do not necessarily mean that our SPS is not reflecting permanence. The degree of classification success was performed comparing the SPS value and the observed flow category in the exact sampling time (Fig. 1), but some sites could have flow categories in one condition during the time of sampling and change to another flow category at the end of summer. Thus, to avoid circularity, information on flow permanence of each site at the end of summer should ideally be obtained to correctly calculate the degree of misclassification of SPS, which was impossible in our case. On the other hand, other factors not measured in this study, but locally important in determining permanence (e.g., riparian vegetation or forest reaching the river banks), could exacerbate summer drought even when habitat conditions in the channel were satisfactory.
Macroinvertebrate structure and biological traits
Significant differences in winter and summer taxon richness along the SPS gradient were found among permanent, intermittent and ephemeral sites, for total (H = 14.43, P < 0.001), winter (H = 12.45, P = 0.002) and summer taxon richness (H = 13.52, P = 0.001) (Fig. 2). There was a clear gap between ephemeral sites and those corresponding to the other two flow regimes. Taxon richness was lower in winter than in summer for both intermittent and permanent sites, but winter samples added an average of 3 new taxa to total richness. No differences were found when comparing permanent and intermittent richness in summer (H = 0.05, P = 0.819), winter (H = 0.21, P = 0.647) or overall (H = 0.10, P = 0.750).

Variation in the number of taxa (total, winter and summer) in the sites along the SPS gradient, from permanent to ephemeral sites. For location of sites see Fig. 1
In permanent and intermittent sites, a higher EPT than OCH was found in winter whereas in summer, OCH richness increased (Fig. 3). The EPT and OCH metrics showed significant differences in winter among permanent, intermittent and ephemeral sites (EPT: H = 11.33, P = 0.003; OCH: H = 8.31, P = 0.015). However, significant differences were not detected between permanent and intermittent sites in winter (EPT: H = 1.54, P = 0.214; OCH: H = 0.52, P = 0.468) and summer (EPT: H = 0.20, P = 0.651; OCH: H = 0.06, P = 0.804) (Fig. 3).

Mean and standard deviation of the EPT and OCH (see text for definition) values for permanent, intermittent and ephemeral sites in winter and summer period
Results from the “4th Corner method” analysis showed distinct patterns of biological traits in permanent, intermittent and ephemeral streams (Table 2). Permanent sites had few significant trait categories compared with the other two flow categories. Life-history characteristics were not significant in any habitat group. Other categories related to size and aquatic stages also presented non-overall significance (see F-values). The remaining traits were significant for one or several flow categories, although correlation values were low. Thus, isolated eggs and asexual reproduction were positively correlated with ephemeral sites but negatively with intermittent ones. In contrast, intermittent sites were characterized by clutches in vegetation. Dispersion mechanisms were aerial-active in intermittent sites but aquatic-passive in ephemeral ones; the latter also presented a negative correlation with non-resistant forms, while cocoons were a significant strategy at these sites. Diapause was positively associated with intermittent sites and negatively with ephemeral ones but without an overall significance. Respiration through skin was positively associated with ephemeral sites whereas the possession of spiracles was associated with the intermittent sites. Organisms in intermittent sites were substantially fliers and surface swimmers, whereas ephemeral sites had significant and positive relationships with burrowers and interstitial organisms but negative ones with fliers and swimmers. Food and feeding habits in ephemeral sites were characterized by absorbers (e.g., some Oligochaeta—Tachet et al., 2002) and deposit feeders that consume detritus and microorganisms. In contrast, intermittent sites were associated with organisms that feed on living microinvertebrates.
Discussion
Spatial and temporal heterogeneity affect organisms independently, but the interaction between the two can determine patterns and processes (Keddy, 1991). Relationships between these two types of heterogeneity are difficult to examine because of their complexity and because of methodological constraints (Kolasa & Rollo, 1991). Some studies have addressed these two types of heterogeneity (Resh & Rosenberg, 1989; Watling & Press, 2000; Wiens, 2000), but few others report the interaction between them and their relationship with biological traits (e.g., Townsend et al., 1997). Using a simple quantitative index (SPS), we show how habitat characteristics (substrate and flow) influence flow permanence of a stream site and constrain macroinvertebrate structure and biological traits. Several methods of characterizing river types using hydrological variables have been described (including recent and historical descriptors, e.g., Poff & Ward, 1989; Poff, 1996; Uys & O’Keefe, 1997; Olden & Poff, 2003; Robles et al., 2004) and some of them related these classifications to community characteristics (e.g., see Stanley et al., 1997; 2004). Recently, Fritz & Dodds (2005) proposed an index to characterize sites and streams on a continuum of flow intermittency, but we did not have the information in quantity and quality to apply it. The SPS presented here is a simple and a broad outline method to characterize site permanence in Mediterranean river ecosystems. Overall, we found that habitat characteristics determined temporal heterogeneity (i.e., flow stability). Thus, as winter and summer flows increases and bedrock and pools dominate, temporal heterogeneity decreases (i.e., we move from ephemeral to intermittent and to permanent). In addition, in this framework, habitat characteristics also influenced spatial heterogeneity. Higher winter and summer flows results in a more wetted habitat and likely in an increase of available refuges and substrate diversity. Thus, it is reasonable to think that spatial heterogeneity increases from ephemeral to intermittent and to permanent sites (Fig. 4). These three river flow types established in this spatio-temporal framework differed in macroinvertebrate community structure and biological traits. Droughts and natural disturbances in general have been previously considered as determinants of community structure in freshwater ecosystems (Resh et al., 1988; Stanley & Fisher, 1992). Comparisons between permanent and temporary rivers are contradictory, from lower richness in temporary sites (Wright et al., 1984; del Rosario & Resh, 2000) to similar richness (Legier & Talin, 1973; Boulton & Suter, 1986; Miller & Golladay, 1996). In our study, intermittent and permanent sites showed similar taxonomic richness, but higher richness than ephemeral sites. The length of the dry season is crucial in determining diversity in temporary streams (Williams & Hynes, 1976; Abell, 1984; Williams, 1996) and could explain this pattern of species richness in the three flow conditions examined. Fritz & Dodds (2005), in characterizing harshness in several intermittent prairie streams, found that harsher sites (i.e., long dry period, low flow predictability, high flood frequency and low surface connectivity) had lower taxon richness.

Categorization of habitat by flow and substrate, for permanent, intermittent and ephemeral sites. Note the increase in spatial heterogeneity and the decrease in temporal heterogeneity from ephemeral to permanent sites
In a study of the relationship between a gradient of permanence of riffle habitat and macroinvertebrate assemblages, Feminella (1996) found a high correlation between EPT and an increase in flow permanence. A similar pattern was reported by Smith et al. (2003). Our results showed not significant differences in EPT values between permanent and intermittent sites in both seasons, but an overall significant difference in winter when considering ephemeral sites. On the other hand, seasonal changes in OCH values were more evident, and could be attributed to the high proportion of pool conditions in permanent sites and the absence of riffles in intermittent sites in summer (Williams, 1996). During the wet season, riffle-like taxa (EPT) may dominate river beds and some may be replaced by pool-like taxa (OCH) in summer, jointly with a group of core species (e.g., Diptera) with a broader niche and which are present throughout the year (Langton & Casas, 1999; Rieradevall et al., 1999; Bogan & Lytle, 2007). This clear seasonal pattern enhances the use of the relationship between EPT and OCH metrics in the characterization of communities according to flow permanence in Mediterranean rivers, which is also coincident with studies in other Mediterranean climate areas (i.e., in California, Bonada et al., 2006). The relationship between EPT and OCH may be probably used as a worldwide index of quantification of temporary rivers.
Williams (1996) modelled expected biological traits in three types of temporary conditions: seasonal with long periodicity, annually seasonal and unpredictable non-seasonal. Adults with short life duration, intermediate fecundity, good dispersal ability and generalist feeding larvae with some desiccation resistance were proposed as characteristics of annual seasonal conditions in this model. In our study, although several traits (e.g., respiration and locomotion) were consistent with the predictions made by Williams (1996), life-history traits were not significant. The high interannual variability of Mediterranean rivers, with sites drying out one year but possibly not the next, depending on the annual climatic conditions (Gasith & Resh, 1999), may explain our results. Because life-cycle characteristics and organism size are strongly associated with latitude and climatic conditions (e.g., Johansson, 2003), a dominance of multivoltine life-cycles and organisms with a short life-span is to be expected in Mediterranean climate areas, independently of the characteristics of the habitat and its spatial and temporal variability. In fact, when the timing of disturbances is predictable, life-cycles are adapted to long-term dynamics of these disturbances rather than to specific flow events (Lytle & Poff, 2004). In Mediterranean rivers, droughts and floods are mainly seasonal and predictable, occurring twice or three times a year. Under these conditions, multivoltinism and short life-cycles are favoured, particularly for those organisms with low migratory capacity (most of the highly frequent and abundant Ephemeroptera, Plecoptera, Trichoptera). Thus, at least one generation will be ensured before a possible early drought in ephemeral sites or late drought in intermittent or permanent sites. In fact, in comparison with rivers under temperate climatic conditions, Mediterranean areas are highly dominated by multivoltine aquatic organisms (e.g., Caenis or Nebrioporus—Tachet et al., 2002; Bonada et al., in press).
Other traits, such as respiration, locomotion and dispersal, among others, should differ between permanent, intermittent and ephemeral sites. In accordance with our results, changes in habitat structure at intermittent sites when riffles are lost enhance the presence of aerial breathers, fliers and swimmers (Williams, 1996). The rationale for this phenomenon is that these traits are less constrained by climatic characteristics or by other large-scale variables, and are probably less dependent on long-term flow dynamics. However, these traits would be more related to local-scale characteristics such as individual year flow conditions (e.g., winter and summer flow) and local substrate (e.g., presence of bedrock vs. gravel and cobbles and riffles vs. pools). Thus, organisms at a site would be selected from the regional pool of species, in order to match their biological traits with the local flow conditions of a site in a given year; however, most of these organisms (especially those without high migratory capacity) would display similar life-cycles in order to adapt to the short stable periods of floods and droughts over the annual cycle. It does not necessarily mean that all organisms present in all flow categories display multivoltinism and have short life-cycles (e.g., Odonata can be semivoltine even in temporary streams—Ferreras-Romero & García-Rojas, 1995), but that most of them do.
Correlation values from the “4th Corner method” were very low, which is consistent with the findings of similar studies (Legendre et al., 1997). However, with the exception of some traits, such as maximal size and life-cycle characteristics, trends in this Mediterranean river system indicated that our empirical results matched the theoretical predictions of the RHT. Permanent sites did not present any significant traits because riffles and pools are present throughout the year, allowing the presence of a mix of organisms some characterizing ephemeral and intermittent sites (Fig. 5). Intermittent sites were dominated by pool species that colonized this habitat and may colonize others in the event of pool desiccation. The importance of specific drought adaptations (e.g., cocoons) or the hyporheos as a refuge (Magoulick & Kobza, 2003; Hose et al., 2005; Bogan & Lytle, 2007) has not been investigated in this paper, but probably this was one of the main reasons of rapid colonization of the streams studied together with the dispersal abilities of many species. In addition, this rapid colonization by organisms from each site or coming for neighbour sites would allow that all sites in February (i.e., after several months of permanent flow since the last dry period) were comparable, independently of their spatial position.

Application of permanent, intermittent and ephemeral conditions to the River Habitat Templet Theory proposed by Townsend & Hildrew (1994) in the Mediterranean basin studied
In summary, our results show that the effect of drought, and thus its intensity, also depends on local habitat characteristics. Permanent sites were present in reaches with constant (from springs) winter and summer flow over a variety of substrates. Intermittent sites were found in low flow conditions over bedrock, thereby facilitating the persistence of pools in summer. Finally, the most ephemeral sites occurred with low winter flow over gravel and cobbles. We found that habitat exerts a strong constrain on macroinvertebrate communities by constraining their richness, structure and traits. While some biological traits were more affected by local habitat characteristics, others were more influenced by regional characteristics related to the predictability of flood and drought timing of the Mediterranean climate.
References
Abell, D. L., 1984. Benthic Invertebrates of some California Intermittent Streams. In Jain, S. & P. Moyle (eds), Vernal Pools and Intermittent Streams. Institute of Ecology Publication, UC Davis, Davis, 46–60.
Biggs, B. J. F., R. J. Stevenson & R. L. Lowe, 1998. A habitat matrix conceptual model for stream periphyton. Archiv fur Hydrobiologie 143: 21–56.
Bogan, M. T. & D. A. Lytle, 2007. Seasonal flow variation allows ‘time-sharing’ by disparate aquatic insect communities in montane desert streams. Freshwater Biology 52: 290–304.
Bonada, N., M. Rieradevall, N. Prat & V. H. Resh, 2006. Benthic macroinvertebrate assemblages and macrohabitat connectivity in Mediterranean-climate streams of northern California. Journal of the North American Benthological Society 25: 32–43.
Bonada, N., S. Dolédec & B. Statzner, in press. Taxonomic and biological trait differences of stream macroinvertebrate communities between mediterranean and temperate regions: implications for future climatic scenarios. Global Change Biology .
Boulton, A. J., 2003. Parallels and contrasts in the effects of drought on stream macroinvertebrate assemblages. Freshwater Biology 48: 1173–1185.
Boulton, A. J. & P. J. Suter, 1986. Ecology of Temporary Streams—an Australian Perspective. In De Deckker, P. & W. D. Williams (eds), Limnology in Australia. Junk Publishers, Melbourne, 313–327.
Boulton, A. J. & P. S. Lake, 1992a. The ecology of two intermittent streams in Victoria, Australia. II. Comparisons of faunal composition between habitats, rivers and years. Freshwater Biology 27: 99–121.
Boulton, A. J. & P. S. Lake, 1992b. The ecology of two intermittent streams in Victoria, Australia. III. Temporal changes in faunal composition. Freshwater Biology 27: 123–138.
del Rosario, R. B. & V. H. Resh, 2000. Invertebrates in intermittent and perennial streams: is the hyporheic zone a refuge from drying? Journal of the North American Benthological Society 19: 680–696.
Delucchi, C. M., 1988. Comparison of community structure among streams with different temporal flow regimes. Canadian Journal of Zoology 66: 579–586.
Delucchi, C. M. & B. L. Peckarsky, 1989. Life history patterns of insects in an intermittent and a permanent stream. Journal of the North American Benthological Society 8: 308–321.
di Castri, F., 1981. Mediterranean-type Shrublands of the World. In di Castri, F., D. W. Goodall & R. L. Specht (eds), Mediterranean-type Shrublands. Elsevier Scientific Publishing Company, Amsterdam, The Netherlands, 1–643.
Dolédec, S., D. Chessel, C. J. F. Ter Braak & S. Champely, 1996. Matching species traits to environmental variables: a new three-table ordination method. Environmental and Ecological Statistics 3: 143–166.
Feminella, J. W., 1996. Comparision of benthic macroinvertebrates assemblages in small streams along a gradient of flow permanence. Journal of the North American Benthological Society 15: 651–669.
Ferreras-Romero, M. & A. M. García-Rojas, 1995. Life-history patterns and spatial separation exhibited by the Odonates from a Mediterranean inland catchment in southern Spain. Vie Milieu 45: 157–166.
Fritz, K. M. & W. K. Dodds, 2005. Harshness: characterization of intermittent stream habitat over space and time. Marine and Freshwater Research 56: 13–23.
Gasith, A. & V. H. Resh, 1999. Streams in mediterranean climate regions: abiotic influences and biotic responses to predictable seasonal events. Annual Review of Ecology, Evolution and Systematics 30: 51–81.
Hose, G. C., P. Jones & R. P. Lim, 2005. Hyporheic macroinvertebrates in riffle and pool areas of temporary streams in south eastern Australia. Hydrobiologia 532: 81–90.
Johansson, F., 2003. Latitudinal shifts in body size of Enallagma cyathigerum (Odonata). Journal of Biogeography 30: 29–34.
Joly, P. & A. Morand, 1994. Theoretical habitat templets, species traits, and species richness: amphibians in the Upper Rhône River and its floodplain. Freshwater Biology 31: 455–468.
Keddy, P. A., 1991. Working with Heterogeneity: an Operator’s Guide to Environmental Gradients. In Kolasa, J. & T. A. Pickett (eds), Ecological Heterogeneity. Springer-Verlag, New York, New Jersey, 181–201.
Kolasa, J. & C. D. Rollo, 1991. The Heterogeneity of Heterogeneity: a Glossary. In Kolasa, J. & T. A. Pickett (eds), Ecological Heterogeneity. Springer-Verlag, New York, New Jersey, 1–23.
Lake, P. S., 2000. Disturbance, patchiness, and diversity in streams. Journal of the North American Benthological Society 19: 573–592.
Lamouroux, N., N. L. Poff & P. Argermeier, 2002. Intercontinental convergence of stream fish community traits along geomorphic and hydraulic gradients. Ecology 83: 1792–1807.
Lamouroux, N., S. Dolédec & S. Gayraud, 2004. Biological traits of stream macroinvertebrate communities: effects of microhabitat, reach, and basin filters. Journal of the North American Benthological Society 23: 449–466.
Langton, P. H. & J. Casas, 1999. Changes in chironomid assemblage composition in two Mediterranean mountain streams over a period of extreme hydrological conditions. Hydrobiologia 390: 37–49.
Legendre, P. & L. Legendre, 1998. Numerical Ecology. Elsevier, Amsterdam.
Legendre, P., R. Galzin & M. Harmelin-Vivien, 1997. Relating behavior to habitat: solutions to the fourth-corner problem. Ecology 78: 547–562.
Legier, P. & J. Talin, 1973. Comparaison de ruisseaux permanents et temporaries de la Provence calcarie. Annales de Limnologie 9: 273–292.
Lytle, D. A. & N. L. Poff, 2004. Adaptation to natural flow regimes. Trends in Ecology and Evolution 19: 94–100.
Magoulick, D. D. & R. Kobza, 2003. The role of refugia for fishes during drought: a review and synthesis. Freshwater Biology 48: 1186–1198.
McElravy, E. P., G. A. Lamberti & V. H. Resh, 1989. Year-to-year variation in the aquatic macroinvertebrate fauna of a northern California stream. Journal of the North American Benthological Society 8: 51–63.
Miller, A. M. & S. W. Golladay, 1996. Effects of spates and drying on macroinvertebrate assemblages of an intermittent and a perennial prairie stream. Journal of the North American Benthological Society 15: 670–689.
Milne, B. T., 1991. Heterogeneity as a Multiscale Characteristic of Landscapes. In Kolasa, J. & T. A. Pickett (eds), Ecological Heterogeneity. Springer-Verlag, New York, 32–69.
Muotka, T. & R. Virtaten, 1995. The stream as a habitat templet for bryophytes: species’ distributions along gradients in disturbance and substratum heterogeneity. Freshwater Biology 3: 141–160.
Olden, J. D. & N. L. Poff, 2003. Redundancy and the choice of hydrologic indices for characterizing streamflow regimes. River Research and Applications 19: 101–121.
Pattee, E. & H. Chergui, 1995. The application of habitat templets and traits to hyphomycete fungi in a mid-European river system. Freshwater Biology 33: 525–539.
Persat, H., J.-M. Olivier & D. Pont, 1994. Theoretical habitat templets, species traits, and species richness: fish in the Upper Rhône River and its floodplain. Freshwater Biology 31: 439–454.
Poff, N. L., 1996. A hydrogeography of unregulated streams in the United States and an examination of scale-dependence in some hydrological descriptors. Freshwater Biology 36: 71–91.
Poff, N. L., 1997. Landscape filters and species traits: towards mechanistic understanding and prediction in stream ecology. Journal of the North American Benthological Society 16: 391–409.
Poff, N. L. & J. V. Ward, 1989. Implications of streamflow variability for lotic community structure: a regional analysis of streamflow patterns. Canadian Journal of Fisheries and Aquatic Sciences 46: 1805–1817.
Poff, N. L. & J. V. Ward, 1990. Physical habitat template of lotic systems: recovery in the context of historical pattern of spatiotemporal heterogeneity. Environmental Management 14: 629–645.
Poff, N. L. & J. D. Allan, 1995. Functional organization of stream fish assemblages in relation to hydrological variability. Ecology 76: 606–627.
Poff, N. L., J. D. Olden, N. K. M. Vieira, D. S. Finn, M. P. Simmons & B. C. Kondratieff, 2006. Functional trait niches of North American lotic insects: traits-based ecological applications in light of phylogenetic relationships. Journal of the North American Benthological Society 25: 730–755.
Prat, N., M. A. Puig, G. Gonzàlez, M. F. Tort & M. Estrada, 1984. Llobregat. In Whitton B. A. (ed.), Ecology of European Rivers. Blackwell Scientific Publications, Oxford: 527–551.
Prat, N. & J. V. Ward, 1994. The Tamed River. In Margalef, R. (ed.), Lymnology now a Paradigm of Planetary Problems. Elsevier Science: 219–236.
Prat, N., A. Munné, M. Rieradevall, F. Carcellar, J. Fons, G. Chacón, C. Ibañez, X. Font, J. M. Carmona & A. Romo, 2000. Biodiversity of a Mediterranean stream drainage network. Verhandlungen International Verein Limnology 27: 137–139.
Resh, V. H. & D. M. Rosenberg, 1989. Spatial-temporal variability and the study of aquatic insects. The Canadian Entomologist 121: 941–963.
Resh, V. H., A. V. Brown, A. P. Covich, M. E. Gurtz, H. W. Li, G. W. Minshall, S. R. Reice, A. L. Sheldon, J. B. Wallace & R. C. Wissmar, 1988. The role of disturbance in stream ecology. Journal of the North American Benthological Society 7: 433–455.
Resh, V. H., A. G. Hildrew, B. Statzner & C. R. Townsend, 1994. Theoretical habitat templets, species traits, and species richness: a synthesis of long-term ecological research on the Upper Rhône River in the context of concurrently developed ecological theory. Freshwater Biology 31: 539–554.
Ribera, I., S. Dolédec, I. S. Downie & G. N. Foster, 2001. Effect of land disturbance and stress on species traits of ground beetle assemblages. Ecology 82: 1112–1129.
Rieradevall, M., N. Bonada & N. Prat, 1999. Community structure and water quality in the mediterranean streams of a natural park (St. Llorenç del Munt, NE Spain). Limnetica 17: 45–56.
Robles, S., M. Toro, C. Nuño, J. Avilés, J. Alba-Tercedor, M. Álvarez, N. Bonada, J. Casas, P. Jáimez-Cuéllar, A. Mellado, A. Munné, I. Pardo, N. Prat, M. L. Suárez, M. R. Vidal-Abarca, S. Vivas, G. Moyá & G. Ramon, 2004. Descripción de las cuencas mediterráneas seleccionadas en el proyecto GUADALMED. Limnetica 21(2002): 35–61.
Smith, H., P. J. Wood & J. Gunn, 2003. The influence of habitat structure and flow permanence on invertebrate communities in karst spring systems. Hydrobiologia 510: 53–66.
Southwood, T. R. E., 1977. Habitat, the templet for ecological strategies? Journal of Animal Ecology 46: 337–365.
Southwood, T. R. E., 1988. Tactics, strategies and templets. Oikos 52: 3–18.
Southwood, T. R. E., R. M. May, M. P. Hassell & G. R. Conway, 1974. Ecological strategies and population parameters. American Naturalist 108: 791–804.
Stamou, G. P., 1998, Arthropods of Mediterranean-type Ecosystems. Springer-Verlag, Berlin, 135.
Stanley, E. H. & S. G. Fisher, 1992. Intermittency, Disturbance, and Stability in Stream Ecosystems. In Robarts, R. D. & M. L. Bothwell (eds), Aquatic Ecosystems in Semiarid Regions: Implications for Resource Management. Environmental Canada, Saskatoon: 271–280.
Stanley, E. H., S. G. Fisher & N. B. Grimm, 1997. Ecosystem expansion and contraction in streams. BioScience 47: 427–435.
Stanley, E. H., S. G. Fisher & J. B. J. Jones, 2004. Effects of water loss on primary production: a landscape-scale model. Aquatic Sciences 66: 130–138.
Statzner, B., V. H. Resh & S. Dolédec (eds), 1994. Ecology of the Upper Rhône River: a Test of Habitat Templet Theories. Special Issue Freshwater Biology 31: 253–554.
Statzner, B., K. Hoppenhaus, M. F. Arens & P. Richoux, 1997. Reproductive traits, habitat use and templet theory: a synthesis of world-wide data on aquatic insects. Freshwater Biology 38: 109–135.
Tachet, H., P. Richoux, M. Bournaud & P. Usseglio-Polatera, 2002. Invertébrés d’Eau Douce (2nd corrected impression). CNRS éditions, Paris.
Thuiller, W., S. Lavorel, G. Midgley, S. Lavergne & T. Rebelo, 2004. Relating plant traits and species distributions along bioclimatic gradients for 88 Leucadendron taxa. Ecology 85: 1688–1699.
Townsend, C. R. & A. G. Hildrew, 1994. Species traits in relation to a habitat templet for river systems. Freshwater Biology 31: 265–275.
Townsend, C. R., M. R. Scarsbrook & S. Dolédec, 1997. Quantifying disturbance in streams: alternative measures of disturbance in relation to macroinvertebrate species traits and species richness. Journal of the North American Benthological Society 16: 531–544.
Usseglio-Polatera, P., 1994. Theoretical habitat templets, species traits, and species richness: aquatic insects in the Upper R¨hône and its floodplain. Freshwater Biology 31: 417–437.
Usseglio-Polatera, P., M. Bournaud, P. Richoux & H. Tachet, 2000. Biological and ecological traits of benthic freshwater macroinvertebrates: relationships and definition of groups with similar traits. Freshwater Biology 43: 175–205.
Uys, M. C. & J. H. O’Keefe, 1997. Simple words and fuzzy zones: early directions for temporary research in South Africa. Environmental Management 21: 517–531.
Watling, J. R. & M. C. Press, 2000. Light Heterogeneity in Tropical Rain Forests: Photosynthetic Responses, their Ecological Consequences. In Hutchings, M. J., E. A. John & A. J. A. Stewart (eds), The Ecological Consequences of Environmental Heterogeneity. Blackwell Science, Oxford, 131–154.
White, P. S. & J. Harrod, 1997. Disturbance and Diversity in a Landscape Context. In Bissonette, J. A. (ed.), Wildlife and Landscape Ecology: Effects of Pattern and Scale. Springer-Verlag, New York, 128–159.
Wiens, J. A., 2000. Ecological Heterogeneity: an Ontogeny of Concepts and Approaches. In Hutchings, M. J., E. A. John & A. J. A. Stewart (eds), The Ecological Consequences of Environmental Heterogeneity. Blackwell Science, Oxford, 9–32.
Williams, D. D., 1991. Life history traits of aquatic arthropods in springs. Memoirs of the entomological society of Canada 155: 63–87.
Williams, D. D., 1996. Environmental constraits in temporary fresh waters and their consequences for the insect fauna. Journal of the North American Benthological Society 15: 634–650.
Williams, D. D. & H. B. N. Hynes, 1976. The ecology of temporary streams. I. The fauna of two Canadian streams. International Review Gesamten Hydrobiologie 61: 761–787.
Wood, P. J. & P. D. Armitage, 1997. Biological effects of fine sediment in the lotic environment. Environmental Management 31: 203–217.
Wright, J. F., P. D. Hiley, D. A. Cooling, A. C. Cameron, M. E. Wigham & A. D. Berrie, 1984. The invertebrate fauna of a small chalk stream in Berkshire, England, and the effect of intermittent flow. Archiv fur Hydrobiologie 99: 176–199.
Acknowledgements
We thank P. Usseglio-Polatera for kindly providing the data on the degree of affinity of trait groups. We are very grateful to L. Bêche, J.D. Olden, V.H. Resh and D.D. Williams for their constructive comments on previous drafts of the manuscript, to M.À. Rodríguez and S. Vives for statistical advice. This research was supported by the “Diputació de Barcelona—Servei de Parcs Naturals” and a grant to N.B. from the “Ministerio de Ciencia y Tecnología” (Spain). We also thank Steven Declerck and two anonymous reviewers for their comments on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: S. Declerck
Electronic supplementary material
Below are the electronic supplementary materials.
Rights and permissions
About this article
Cite this article
Bonada, N., Rieradevall, M. & Prat, N. Macroinvertebrate community structure and biological traits related to flow permanence in a Mediterranean river network. Hydrobiologia 589, 91–106 (2007). https://doi.org/10.1007/s10750-007-0723-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-007-0723-5