
Abstract
The aim of the study was to evaluate the long-term development of a hard bottom benthic assemblage over a period of 20 years in an area off the mouth of a large river. The artificial reef of Fregene was selected because benthic assemblage data were available for the period 1981–1992. This artificial reef is located in the mid Tyrrhenian Sea, 5 nautical miles north of the two mouths of the Tevere River (Latium, Italy) and 1.5 nautical miles offshore from Fregene (Rome, Italy). The artificial reef was deployed in March 1981 for fisheries enhancement in 10–14 m of water on a sandy-silty seabed. The Tevere River carries suspended materials and a heavy load of organics since it transports Rome’s effluent, resulting in the eutrophic state of area waters. Benthic sampling was conducted in 2001 by SCUBA diving; two standard surfaces of 400 cm2 were scraped from the vertical walls of the same uppermost block in four different periods. All organisms were identified and counted. The methodology used is the same as that adopted in the previous periods, so that the 2001 data could be compared with past collected data. The benthic assemblage was analysed by cluster analysis using the Bray-Curtis index and clustered using the group average clustering algorithm. The SIMPER procedure was used to identify those taxa that characterize each station group identified by cluster analysis. Changes in benthic assemblages and hydrological trends of the Tevere River were investigated using the cumulative sum series method. The 20-year development of the benthic community, starting from the new substratum, is composed of different phases characterised by different benthic assemblages. In particular five different phases were distinguished: 1. Pioneer species recruitment (May 1981–June 1981); 2. Mytilus galloprovincialis (mussel) dominance (August 1981–November 1983); 3. M. galloprovincialis regression (July 1984–October 1985); 4. M. galloprovincialis absence (91–92); 5. Bryozoans bioconstruction dominance (2001). The dynamic succession of the observed benthic assemblages exhibited a good relation with the Tevere River flow. The Tevere River flow, and the subsequent sedimentation process, seems to have strongly influenced the benthic assemblage succession of the Fregene artificial reef.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Knowledge of long-term benthic assemblages development, and in particular assemblages associated with hard bottom, is poor. Many research projects have been undertaken that describe benthic succession using different experimental approaches involving artificial reefs or defaunated natural substrata (Bourget et al., 1994; Brown & Swearingen, 1998; Kocak & Zamboni, 1998; Glasby & Connell, 2001). Most of these studies were concerned with benthic primary colonisation patterns and therefore were short-term studies. On the other hand, long-term investigations that follow later development phases of benthic assemblages have rarely been conducted. A long-term study, including origination on a newly deployed substratum, could offer insight into understanding the most important changes that occur in the benthic assemblage structure during and after the colonisation process (Wiens, 1997). Due to their sessile habit, benthic fauna facilitates the evaluation of environmental quality, as modification of such communities may be directly traceable to natural or anthropogenic environmental variations (Pearson & Rosenberg, 1978; Gray, 1981; Gray et al., 1990; Warwick & Clarke, 1991). A long-term investigation can help to elucidate the response that benthic communities manifest to environmental changes in terms of sedimentation and turbidity. The objective of this study is to evaluate the development of a hard-bottom benthic assemblage over a period of 20 years in an area off the mouth of a large river. We chose to evaluate the benthic assemblage associated with an artificial substratum which, due to its structural simplicity, facilitated quantification relative to area and time (Riggio, 1995; Glasby & Connell, 2001; Smith & Rule, 2002). In particular, this study analysed the benthic assemblage of the Fregene artificial reef, placed in an area where the Tevere River strongly influences the water conditions through a significant organic and inorganic load. The Tevere River is the main river of the central Tyrrhenian Sea and has an effect on the eutrophication process, in particular on the north coast. In fact, the principal direction of the current, and thus of suspended sediment, is northwest along the coast (Bellotti & Tortora, 1985; La Monica & Raffi, 1996). In the last few decades, coastal erosion along the Latium coast of Italy has been particularly evident and different factors have contributed to this process. One of these was the decrease in the transportation of solids due to dam construction, sand extraction from riverbeds, etc. The river’s solid loads vary with river flow which is also influenced by climatic events that affect the drainage basin (Bellotti & Tortora, 1996; Bencivenga & Ranieri, 1997). The changes in sedimentation and turbidity in the deltaic areas off large rivers can lead to alteration in associated benthic community. An extended time series of data is necessary to evaluate changes in both benthic communities and river flows.
Materials and methods
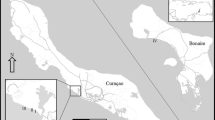
The artificial reef off Fregene is located in the mid Tyrrhenian Sea, 5 nautical miles North of the two mouths of the Tevere River (Latium, Italy) and 1.5 nautical miles offshore Fregene (Rome, Italy) (Fig. 1). The artificial reef was placed at 10–14 m depth on a sandy-silty seabed. The reef is composed of 280 concrete cube-shaped units (2 m on each side) arranged in a pyramid consisting of five units to a set, four at the base and one on the top. A total of 60 sets were deployed as a reef group that covered 6 ha. Each cube contains hollows and cavities moulded into the shape in order to increase the surface/volume ratio (Bombace, 1977). The Tevere River carries suspended materials and a significant organic load, resulting in eutrophic conditions. Benthic sampling was performed in 2001 (February, June, September and December) using SCUBA. During each sampling event, two standard surfaces of 400 cm2 were scraped from the vertical wall of the same top block. Different parts of the vertical wall were chosen in order to avoid a re-sampling on surfaces previously scraped. The methodology used is the same as that adopted in the previous periods, so that the 2001 data could be compared with past collected data (Ardizzone et al., 1989; Somaschini et al., 1997). Samples were fixed in buffered formaldehyde (10%). All organisms were sorted under a stereomicroscope. Polychaetes, molluscs, crustaceans and echinoderms were identified to species level and counted. The Bray-Curtis index was used as faunal similarity coefficients calculated using species/abundance matrix. This index was clustered using the group average clustering algorithm. The SIMPER procedure (similarity percentage analysis) was used to identify those taxa that characterize each station group identified by cluster analysis (Clarke, 1993). Hydrological trends of the Tevere River were investigated using the cumulative sum series method (Ibanez et al., 1993; Salen-Picard et al., 2003).
Results
Composition of benthic assemblage in 2001
In 2001 a total of 15,306 benthic invertebrate individuals and a total of 172 taxa were collected. Most benthic species were polychaetes (109 species and 6,918 individuals), followed by crustaceans (22 species and 7,257 individuals) and molluscs (24 species and 1,148 individuals). Among the polychaetes the most abundant species were sessile and suspension-feeder species such as Sabellaria spinulosa, Hydroides pseudouncinatus pseudouncinatus, Serpula concharum, and soft bottom deposit-feeder species such as Aphelocheta marioni. Balanus perforatus was the most abundant crustacean species. The molluscs Striarca lactea and Gastrochaena dubia, characteristic sessile species of hard-bottoms, were very abundant. The benthic assemblage is also characterized by the presence of colonies of two bryozoans, Schizoporella errata and Turbicellepora magnicostata, which cover the whole reef surface by three-dimensional colonies.
Temporal trend in assemblage structure and composition
The number of species showed a progressive increase during benthic assemblages’ temporal succession (Fig. 2). The number of individuals increased from 1981 to May 1983 and then decreased until December 1991. Subsequently, the number of individuals increased again until 2001. Cluster analysis indicates that the year is a chief factor in faunal similarity (Fig. 3). The benthic assemblage sampled in May 1981, 2 months after the artificial reef deployment, when the community was composed of very few pioneer species, is least similar to all other assemblages. A second cluster of assemblages (similarity value 20%) is distinct from the assemblages collected from the 1980s to the ones of the 1990s and 2001. The faunal assemblage collected from the 1980s indicates that the assemblage collected in June 1981 was less similar (30%) to the others. This period also represents a time when the assemblage on the reef was still represented by pioneer species and few individuals. The benthic faunal samples collected between 1981 and 1985 were similar to each other at level of 60%. At the 65% level of similarity the assemblages formed three distinct clusters relative to the periods August 1981–February 1982, March 1982–November 1983 and July 1984–October 1985. The assemblages from 1991–1992 and 2001 were different from the previous assemblages; their similarity was 50% and their respective stations appeared homogeneous. Table 1 shows the results of SIMPER analysis for the identified assemblages clusters.
The monthly cumulative sum curve of monthly flow shows a clear upward trend between 1981 and 1985 and consequently an increase of quantity of sediment in the sea waters. Subsequently (1990–2001) this curve displays a reversed trend (Fig. 4) interpreted as a decrease of the terrigenous particles distributed from the Tevere to sea waters.
Discussion
Five different time-phases as determined by faunal similarity can be distinguished in the cluster analysis.
-
1.
Pioneer species recruitment (May 1981–June 1981). During this phase the reef was colonised only by hydroids (Obelia dichotoma, Bougainvillia ramosa), serpulid polychaetes (Pomatoceros triqueter, Hydroides elegans), barnacles (Balanus eburneus and B. perforatus) and molluscs (Anomia ephippium and Mytilus galloprovincialis). Three months after artificial reef establishment, the mussel M. galloprovincialis was the most abundant species in the assemblage. Notably, macroalgae were absent.
-
2.
Mytilus galloprovincialis dominance (August 1981–November 1983). The mussel M. galloprovincialis dominated the benthic assemblage influencing the settlement of other epibenthic species. Mussel valves and barnacles altered the topography of substratum that became more heterogeneous, this gave origin to an increase in both number of individuals and species (Ardizzone et al., 1982a, b; Ardizzone & Chimenz, 1982).
-
3.
Mytilus galloprovincialis regression (July 1984–October 1985). Gradual reduction of mussel density was observed and soft bottom species became frequent next to the hard bottom ones. Over-sedimentation and “mussel mud” production caused a decrease in the availability of hard substrata for epibenthic species colonization (Gravina et al., 1989).
-
4.
Mytilus galloprovincialis absence (1991–1992). A new benthic assemblage, characterised by the absence of M. galloprovincialis and the numerical abundance of soft bottom species, established itself on the hard and very muddy artificial substratum after 10 years of artificial reef deployment. Bryozoans Schizoporella errata and Scrupocellaria reptans were the only sessile organism extensively distributed on the reef (Ardizzone et al., 1996).
-
5.
Bryozoans bioconstruction dominance (2001). The benthic assemblage, 20 years after artificial reef deployment, is characterised by the presence of colonies of the bryozoans S. errata and Turbicellepora magnicostata. These bryozoans are strong space competitors and are able to build rigid frameworks. They are important “bioconstructors” sensu Bianchi (2001). The benthic assemblage present in 2001 is different from the ones observed in the previous periods when the bioconstructions were absent and there is a strong increase in the number of species typical of both hard and soft substrates.
The artificial reef at Fregene is located in a transitional zone, characterised by abiotic factors typical of the infralittoral zone (high hydrodynamism and eutrophic water) and the circalittoral zone (reduced illumination). During the second period of succession, a mussel dominated assemblage, characteristic of shallow exposed and eutrophic environments, was present. The colonisation of the artificial reef at Fregene was similar to the colonisation observed on other artificial reefs in the Adriatic Sea. These other artificial reefs in the Adriatic were also dominated by M. galloprovincialis (Fabi et al., 1986, 1989). The increased sediment recorded during 1981–1985 (Fig. 4) can be offered as one of the reasons that led to the decline in mussels. Other species, already present as a small percentage of the assemblage on the muddy substrate, became dominant. Later, from 1991 to 1992, the assemblage was characterised by the presence of sciaphilous species typical of muddy infralittoral hard bottoms. Subsequently, the ensuing dramatic decrease of the terrigenous particles from the Tevere (Fig. 4) may have favoured the high presence of suspension-feeding organisms, such as bryozoans. In 2001 these organisms, together with serpulids, barnacles and vermetids, gave rise to bioconstructions which changed the edaphic reef structure. Consequently, habitat heterogeneity increased providing new habitat and thus facilitating the increase in biological diversity (Bianchi & Morri, 1985; Ferdeghini et al, 2000, 2001; Cocito et al, 2000). The presence of these biocontructions allowed the presence of many species typical of “coralligenous” biocoenosis (i.e., polychaetes Palola siciliensis, Serpula vermicularis, Eunice schizobranchia, Eunice oerstedi, Haplosyllis spongicola and the crustacean Leptochelia savignyi). In fact, the bioconstruction present on the artificial reef is similar to a “coralligenous” bioconstruction in morphology and composition, thereby favouring the settlement of many typical coralligenous habitat species. The coexistence of these species with infralittoral species favours the maintenance of a high biodiversity. Due to the high heterogeneity of substratum the benthic settlement was also characterized by the presence of soft bottom species that occupied the interstices filled with mud in the bryozoan bioconstruction and the spaces left by dead organisms. A schematic representation of the cumulative sum series of the river juxtaposed with the different phases of the benthic assemblage succession is reported in Fig. 4.

Map showing the study area and location of the artificial reef at Fregene

Raw data and cumulative sum series of mean monthly flow of the Tevere River from 1980 to 2001. Raw data are compared to the average monthly flow calculated over the period 1980–2001. The reference value used to calculate the cumulative sum series is the long-term mean flow of the river (193 m3 s−1)

Cluster diagram by group-average clustering based on the Bray-Curtis similarity index using the abundance of species

Temporal trend of species richness (S) and abundance (N)
Conclusion
This study underlines the importance of long-term investigations when attempting to understand benthic community succession. Long-term studies allow detection of the effects of rare events and slow acting processes, revealing subtle but consistent trends and environmental changes. Study results from short-term studies using static “snap-shots” of reality may not be able to detect these changes. As a consequence, short-term study results may lead investigators to arrive at incorrect conclusions regarding the dynamics of ecological systems. This 20 years study provided information on the succession of benthic assemblages in relation to the Tevere River flow variability. A benthic community during any point in time is determined by the interaction of biotic and abiotic factors. Among the abiotic factors, the Tevere River flow, and the subsequent sedimentation process, seemed to have influenced the benthic assemblage succession on the artificial reef at Fregene. The present analysis of a benthic community succession provides a contribution that is useful in understanding the dynamics of ecological systems. The analysis of historical data thereby allows a better understanding of how ecosystems behave and develop.
References
Ardizzone, G. D., G. Bombace & P. Pelusi, 1982. Settlement and growth of Mytilus galloprovincialis Lamk on an artificial reef in the Tyrrhenian Sea. Journée Etud. Recifs Artif. et Maricult. suspend., Cannes, Rapp. CIESM: 59–61.
Ardizzone, G. D., & C. Chimenz, 1982a. Primi insediamenti bentonici della barriera artificiale di Fregene. Atti Conv. Ris. Biol. Inq. Mar. P. F. Oceanogr.: Fondi Marini. Roma, 10–11 Novembre 1981: 165–181.
Ardizzone, G. D., C. Chimenz & A. Belluscio, 1982b. Benthic community on the artificial reef of Fregene (Latium). Journée Etud. Récifs artif. Et Maricult. Suspend. – Cannes, C.I.E.S.M.: 55–57.
Ardizzone, G. D., M. F. Gravina & A. Belluscio, 1989. Temporal development of epibenthic communities on artificial reefs in the central Mediterranean Sea. Bulletin of Marine Science 44(2): 592–608.
Ardizzone, G. D., A. Belluscio, M. F. Gravina & A. Somaschini, 1996. Colonization and disappearance of Mytilus galloprovincialis Lamk on an artificial habitat on the Mediterranean Sea. Estuarine, Coastal and Shelf Science 43:665–676.
Bellotti, P. & P. Tortora, 1985. Il delta del Tevere: lineamenti batimetrici, morfologici e tessiturali della conoide sommersa e delle aree limitrofe. Bollettino della Società Geologica Italiana 104: 65–80.
Bellotti, P. & P. Tortora, 1996. I sedimenti sul fondale del delta del Fiume Tevere. Bollettino della Società Geologica Italiana 115: 449–458.
Bencivenga, M. & E. Ranieri, 1997. Il regime dei deflussi del Tevere a Roma. Acqua 3: 51–60.
Bianchi, C. N., 2001. La biocostruzione negli ecosistemi marini e la biologia marina Italiana. Biologia Marina Mediterranea 8(1): 112–130.
Bianchi, C. N. & C. Morri, 1985. I policheti come descrittori della struttura trofica degli ecosistemi marini. Oebalia 11: 203–214.
Bombace, G., 1977. Aspetti teorici e sperimentali concernenti le barriere artificiali. In Cinelli, F., E. Fresi & L. Mazzella (eds), Atti del IX Congresso della Società Italiana di Biologia Marina. Ischia, Naples: 29–41.
Bourget, E., J. DeGuise & G. Daigle, 1994. Scales of substratum heterogeneity, structural complexity, and the early establishment of a marine epibenthic community. Journal of Experimental Marine Biology and Ecology 181(1): 31–52.
Brown, K. M. & D. C. Swearingen, 1998. Effects of seasonality, length of immersion, locality and predation on an intertidal fouling assemblage in the Northern Gulf of Mexico. Journal of Experimental Marine Biology and Ecology 225: 107–121.
Clarke, K. R., 1993. Non-parametric multivariate analyses of changes in community structure. Australian Journal of Ecology 18: 117–143.
Cocito, S., F. Ferdeghini, C. Morri & C. N. Bianchi, 2000. Patterns of bioconstruction in the cheilostome Schizoporella errata: the influence of hydrodynamics and associated biota. Marine Ecology Progress Series 192: 153–161.
Fabi, G., L. Fiorentini & S. Giannini, 1986. Growth of Mytilus galloprovincialis Lamk on a suspended and immersed culture in the Bay of Portonovo (central Adriatic sea). FAO Fisheries Report 357: 144–154.
Fabi, G., L. Fiorentini & S. Giannini, 1989. Experimental shellfish culture on an artificial reef in the Adriatic Sea. Bulletin of Marine Science 44 (2): 923–933.
Ferdeghini, F., S. Cocito, C. Morri & C. N. Bianchi, 2000. Living Bryozoan Buildups: Schizoporella errata (Waters, 1848) (Cheilostomatida, Ascophorina) in the Northwestern Mediterranean (Preliminary Observations). Proceedings of the 11th International Bryozoology Association Conference: 238–244.
Ferdeghini, F., S. Cocito, L. Azzaro, S. Sgorbini & F. Cinelli, 2001. Bryozoan biocostructions in the coralligenous formations of S. M. Leuca (Apulia, Italy). Biologia Marina Mediterranea 8(1): 238–245.
Glasby, T. M. & S. D. Connell, 2001. Orientation and position of substrata have large effects on epibiotic assemblages. Marine Ecology Progress Series 214: 127–135.
Gray, J.S., 1981. The Ecology of Marine Sediments. Cambridge University Press, Cambridge: 185 pp.
Gray, J. S., K. R. Clarke, R. M. Warwick & G. Hobbs, 1990. Detection of initial effects of pollution on marine benthos: an example from the Ekofisk and Eldfisk oilfields, North Sea. Marine Ecology Progress Series 66: 285–299.
Gravina, M. F., G. D. Ardizzone & A. Belluscio, 1989. Polychaetes of an artificial reef in the Central Mediterranean Sea. Estuarine, Coastal and Shelf Science 28: 161–172.
Ibanez, F., J. M. Fromentin & J. Castel, 1993. Application de la méthode des sommes cumulées à l’analyse des séries chronologiques en océanographie. Comptes Rendus de l’Academie des Sciences Paris 316: 745–748.
La Monica, G. B. & R. Raffi, 1996. Morfologia e sedimentologia della spiaggia e della piattaforma continentale interna. In: Il Mare del Lazio. Università degli Studi di Roma “La Sapienza”, Regione Lazio Assessorato Opere e Reti di Servizi e Mobilità: 62–105.
Kocak, F. & N. Zamboni, 1998. Settlement and seasonal changes of sessile macrobenthic communities on the panels in the Loano artificial reef (Ligurian sea, NW Mediterranean). Oebalia XXIV: 17–37.
Pearson, T. H. & R. Rosenberg, 1978. Macrobenthic succession in relation to organic enrichment and pollution of the marine environment. Oceanography and Marine Biology: An Annual Review 16: 229–311.
Riggio, S., 1995. Le barriere artificiali e l’uso conservativo della fascia costiera: risultati dei “Reefs” nella Sicilia N/O. Biologia Marina Mediterranea 2(1): 129–164.
Salen-Picard, C., D. Arlhac & E. Alliot, 2003. Responses of a Mediterranean soft bottom community to short-term (1993–1996) hydrological changes in the Rhone River. Marine Environmental Research 55:409–427.
Smith, S. D. A. & M. J. Rule, 2002. Artificial substrata in a shallow sublittoral habitat: do they adequately represent natural habitats or the local species pool? Journal of Experimental Marine Biology and Ecology 277: 25–41.
Somaschini, A., G. D. Ardizzone & M. F. Gravina, 1997. Long-term changes in the structure of a polychaete community on artificial habitats. Bulletin of Marine Science 60(2): 460–466.
Warwick, R. M. & K. R. Clarke, 1991. A comparison of some methods for analysing changes in benthic community structure. Journal of Marine Biological Association of the United Kingdom 71: 225–244.
Wiens, J. A., 1997. Lengthy ecological studies. Trends in Ecology and Evolution 12(12): 499.
Acknowledgements
In particular we would like to thank Dr. Andrea Belluscio for his support in conducting the field work and Dr. Paola La Valle (Molluscs), Dr. Loretta Lattanzi and Dr. Monica Targusi (Crustaceans) for their time and invaluable dedication to the laboratory activities.
Author information
Authors and Affiliations
Corresponding author
Additional information
Guest editors: G. Relini & J. Ryland
Biodiversity in Enclosed Seas and Artificial Marine Habitats
Rights and permissions
About this article
Cite this article
Nicoletti, L., Marzialetti, S., Paganelli, D. et al. Long-term changes in a benthic assemblage associated with artificial reefs. Hydrobiologia 580, 233–240 (2007). https://doi.org/10.1007/s10750-006-0450-3
Issue Date:
DOI: https://doi.org/10.1007/s10750-006-0450-3