
Abstract
Legislation governing strict-protection nature reserves in Brazil in principle precludes human habitation, but virtually all Amazon reserves are nonetheless inhabited. Historical ecology research reported herein assesses the impacts of occupation and resource use by beiradeiros (forest peasants) on the forests of a strictly designated nature reserve in the Iriri River basin (Brazilian Amazon). The hypothesis is that traditional beiradeiros activities are congruent with the aims of conservation of the reserve because their impacts are either neutral or enhance diversity of forest landscapes and biota. We designed the methodology underlying data collection to integrate forest inventory with archaeological techniques in two contrasting forest types (terra firme and seasonally inundated forest), faunal surveys, freelisting of tree terms by beiradeiros, and participant observation, in order to determine biological diversity indices, forest age and the parameters of traditional knowledge that encode such diversity in local vocabulary. Our research results lead us to reject the premise that traditional peasant activities lead to ecological degradation or impoverishment, and suggest that the rationale underpinning strictly protected nature reserves should be re-examined.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The de facto territory of forest peasants—who call themselves beiradeiros - living along the middle Iriri River, Pará state in the Brazilian Amazon, was included in a Brazilian government designated strict nature reserve, the Terra do Meio Ecological Station (henceforth ESEC-TM), even though many beiradeiros can trace genealogies in the area back six generations to the initial migration of rubber tappers from northeastern Brazil in the last decade of the nineteenth century. We demonstrate through forest inventory and archaeological techniques in two contrasting forest types, faunal surveys, freelisting of tree terms by beiradeiros and participant observation, that the human transformation of the forest on well-drained ground (terra firme) began before the oldest known occupation by forest peasants and that it is an anthropogenic landscape that is more utilized than an adjacent seasonally inundated forest. The principal finding is that current and historical forest peasant populations have not degraded the territory, either terra firme or seasonally inundated forest, after more than a century of occupation and use. Community-managed tropical forests and indigenous territories may be more effective in halting deforestation and reducing fires than strictly enforced nature reserves (Nepstad et al. 2006; Nelson and Chomitz 2011; Porter-Bolland et al. 2012; cf. Terborgh 1999). Our research supports this view. We examine the length of human occupation, the impact of traditional utilization of resources, and the implications of traditional knowledge (TK) on diversity of plants and animals over time to evaluate the effects – negative or positive – of traditional forest peasant occupation on the ecological integrity of the ESEC-TM.
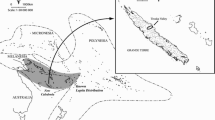
The approximately 3.4 million hectare ESEC–TM was decreed in 2005 (Brasil 2005) as part of a mosaic of protected areas in the Terra do Meio region (Fig. 1) (Schwartzman et al. 2010). ESEC-TM is somewhat larger than Belgium, and covers much of the interfluvial forest between the Xingu and Iriri rivers. It is mostly situated east of the right bank of the Rio Iriri (Fig. 1). The only activities allowed in the ESEC-TM by Brazilian environmental legislation are occupation by government environmental agency staff and scientific researchers’ visits (Brasil 2000 also see Barreto Filho 2004), though in December 2018 in response to a technical report commissioned by the Brazilian federal prosecutor’s office (Ministério Público Federal) and prepared by an interdisciplinary research team (Almeida et al. 2018) government issued a temporary permit for the local forest peasants to remain on the lands they have traditionally occupied inside the ESEC-TM.

The mosaic of conservation units of the Xingu and Iriri River basins (map by Juan Doblas, courtesy Instituto Socioambiental, Altamira)
The initial survey for the mosaic conducted by the Instituto Socioambiental (ISA) for the Environment Ministry proposed the creation of an extractive reserve (Reserva Extrativista, hereafter “Resex”) on the Iriri River including all of the non-indigenous forest peasant communities south of the Cachoeira Seca indigenous territory up to Mekragnoti Kayapô indigenous territory (Fig. 1) (Villas-Bôas et al. 2003). Extractive reserves are, in contrast to the ESEC-TM, sustainable use areas, established at the request of traditional local communities (Santilli 2005). At present, there are 74 forest peasants living inside the ESEC-TM along the Iriri River (ICMBio 2015), for many others abandoned the area after the decree of 2005 (Alarcon and Torres 2014; Almeida et al. 2018). To date, the region has been little studied scientifically, in part because of its remoteness and difficulty of access.
Recent Human Occupation of the Area
Indigenous people historically in the area include Tupí-Guaraní groups, macro-Tupian peoples, and Kayapó societies (Nimuendaju 1948; 1980; Grenand 1982; Viveiros de Castro 1992; Thieme 1993); Rodrigues and Cabral 2012). Although there has been no prior archaeological research on the middle and upper reaches of the Iriri, earlier work on the prehistory of its lower course and within its principal tributary, the Curuá, respectively, points to long term habitation (Perota 1979; Nimuendaju 2004; Roosevelt et al. 2009). In terms of ethnography, Von den Steinen in 1884 and Coudreau in 1896 independently reached and recorded the mouth of the Iriri. Coudreau’s visit coincided with a period of sharply rising prices for Acre fino rubber. The Hevea brasiliensis found in the Xingu basin was considered close in quality to Acre fino (Wagley 2014: 85). In the 1890s, nonindigenous rubber tappers began to populate the Iriri River margins and numbered more than 1000 by the beginning of the twentieth century (Snethlage 2002). Contemporary beiradeiro communities of the Iriri basin are descended largely from these rubber tappers and their families who migrated from the Northeast (o Nordeste) of Brazil, although they have also acquired local traditional knowledge (TK) through interactions with indigenous populations.
The vocabulary of beiradeiros along the Rio Iriri today, specifically in relation to the landscape and its biota, reflects these origins (Table 1), and distinguishes them historically, culturally, and linguistically from the forest peasants of the more central areas of the Amazon River region who are known as “caboclos.”
Since the collapse of an earlier patron-client system beiradeiros have been general extractivists and small-scale farmers (Wagley 2014). Diversity is central to their livelihoods. They collect and market forest resources including rubber, Brazil nuts, babaçu fruits (kernels and mesocarp), tonka beans, and animal hides (Torres 2008; Alarcon and Torres 2014; Villas-Bôas et al. 2017) . As with peasant societies elsewhere, their production strategies are determined by household requirements and the availability of household labor; at the same time, they are, generally indirectly, connected to global markets (Shanin 1984, 2008) and have at times generated income from timber extraction and placer gold mining.
Anthropogenic Landscapes
Levis et al. (2018) address mechanisms of prehistoric landscape transformation in Amazonia, describing cultural or anthropogenic forests as having been domesticated. Primary landscape transformation (Balée 2018) occurs when human agency fundamentally alters, by increase or decrease, species composition and diversity in an area, usually with modification of the substrate, as in structural changes to the environmental gradients of soil or elevation or both. Secondary landscape transformation includes “protection of useful plants,” “attraction of non-human dispersers of useful plants,” “fire management,” “planting,” and “removal of plants not used or of those that interfere with agroforestry traditions and practices,” such as removing lianas from Brazil nut trees (Levis et al. 2018). For example, when people spit out cacao beans into their household gardens after sucking off the sweet pulp around them they inadvertently sow seeds that, after the village moves to a new site, may sprout to become fruiting trees that attract various game animals, including Cebus (organ-grinder) monkeys that, in turn, spread the beans throughout the seemingly abandoned village and surrounding fields—all of which becomes emergent forest resulting from secondary landscape transformation (Balée 1994, 2013).
Rubber tappers were known to plant rubber trees along trails and at campsites (Schroth et al. 2003) that they used when they collected rubber (without destroying the trees) until prices fell in the 1980s, when they abandoned that practice (Schwartzman et al. 2013). They may have felled some caucho trees (Castilla ulei) for white latex in an earlier period (e.g., Hecht 2013), but because the region had a valuable species of rubber (Hevea brasiliensis) in relative abundance, it is unlikely that market demand encouraged the practice. Caucho is also a common species in the anthropogenic forest at the top of one of the geoglyphs in Acre (Balée et al. 2014).
Cacao beans (Theobroma spp.) used for making chocolate for the European market were the most important export from Amazonia in the eighteenth century, including from well-established groves of cacao trees—both domesticated and semi-domesticated--in the Xingu River basin (Betendorf 1910; Maxwell 1973; Alden 1976). Arroyo-Kalin (2016: 11), following Brookfield (1984), considers that this kind of landscape constitutes “landesque capital,” which he defines as “fixed agricultural installations, field systems, and major modifications to the soil ... because they have been intentionally produced to endure beyond the cropping cycle ... fruit tree groves associated with long-fallow slash-and-burn, field systems, anthrosols. ..” We postulate that the indigenous primary landscape transformations in ESEC-TM, which resulted in forests of Brazil nut trees (Bertholletia excelsa), babaçu palms (Attalea speciosa), and cacao groves (Theobroma spp.), together with patches of Amazon Dark Earth (ADE), have been modified only by secondary landscape transformations initiated by beiradeiros and their immediate antecedents.
Methodology
The methodology of historical ecology is by definition interdisciplinary. In this study, in order to understand the influence of beiradeiros on environments of the ESEC-TM, we designed quantitative forest inventories to assess botanical (trees and lianas) alpha and beta diversity as well as absolute dominance, relative density, and relative frequency of trees and lianas across two habitats: forest that is actively and historically in use (terra firme) versus forest that is less used, i.e., seasonally inundated forests, due to current extractivist preferences and activities. Because tree and liana flora are only part of the biota, we carried out a brief complementary survey of the fauna. To understand the long-term human impact on the diversity of the terra firme forest, we conducted archaeological surveys and excavations appropriate to tropical forest (Neves 2000; Machado 2005; Moraes 2006; Lima 2008; Tamanaha 2012). To assess use, management, and knowledge of forest resources and landscapes by beiradeiros, we conducted participant observation and collection of freelist data reflective of widely shared local traditional knowledge (TK) of trees. All field data were collected between August 2016 and April 2018. The zone where forest inventories and archaeological excavations were carried out is located in the center-south of the ESEC-TM, near the right bank of the Igarapé (also called Rio) do Bala, which is itself a tributary of the right bank of the Rio Iriri (Fig. 2).

Area of study, middle Iriri River, showing archaeological site, inventory locations, and Brazil nut grove trails
The area is known locally as “Grota do Cachorro” (Dog Run), which is also the name of an even smaller creek that during the rainy season drains to the right bank of the Igarapé do Bala. The area was selected because initial inspection in August 2016 revealed anthropogenic indicators in the terra firme, ADE and surface potsherds of indigenous (not beiradeiro) manufacture (as reported in Almeida et al. 2018). The forest selected for inventory research was also economically utilized by local beiradeiros (ICMBio 2015). It is specifically part of the territory inhabited since after about 1890 by generations of a beiradeiro family that still lives in the Iriri basin and annually extracts and sells Brazil nuts from the area. That activity does not involve felling the trees; the nuts are collected from the ground.
The climate of the study site and environs is tropical monsoon (Alvares et al. 2013), as per the Köpen classificaton used in Brazil, with a rainy season mostly from February to May (Eletrobras 2009; ICMBio 2015). Average annual temperatures range between 24.60 C (the minimum in July) and 25.40 C (the maximum in September). Average relative atmospheric humidity is 68% with the lowest levels recorded between July and December. Most common soil types on well-drained soils are podsols but there is significant presence of anthropogenic soils or ADE (Fig. 2). In Brazil, the two principal forest types are usually classified physiognomically as Open Tropical Submontane Broadleaf Forest with emergent canopy and Alluvial Broadleaf Forest (along rivers only) (ICMBio 2015).
We carried out the first forest inventory of one hectare in the Tropical Submontane Forest (ICMBio 2015), which we identified as well-drained terra firme, anthropogenic forest (because of the presence of indicators such as ADE). Locally this forest type is known as terra alta. We needed a control site, but untouched primary forest is difficult to find on terra firme, given the extent of indicators of anthropogenesis such as ADE, surface sherds, domesticated trees (such as Brazil nut), semidomesticated, protected, promoted, and occasionally cultivated trees (such as Theobroma speciosum, Castilla ulei, and Hevea brasiliensis) and other disturbance indicator taxa also in that biome (Fig. 3) (also see Clement et al. 2015). Historical ecologists have collected increasing evidence of human disturbance in even apparently unlikely places in the Amazon, including sites far from major waterways (e.g., Levis et al. 2012; Levis et al. 2017).

Measuring a Brazil nut tree (Bertholletia excelsa) on Brazil nut grove trail (Grota do Cachorro), November 2017
The counter-argument, that such anthropogenic or managed forests would be unlikely in the interfluves, is part of the null hypothesis; its proponents (McMichael et al. 2012; McMichael et al. 2014; Piperno et al. 2015, 2019), however, offer limited evidence to rule out prehistoric manipulation in both archaeological and biotic contexts (Levis et al. 2017). In this context, a local option as control site in the area of resource extraction is the riverine, or seasonally flooded forest (baixāo) (Alluvial Broadleaf Forest in the Brazilian classification system), where people do not plant or burn swiddens. We carried out one one-hectare inventory of this forest type, referring to it as the seasonally inundated forest.
We compiled the inventories between August and October 2017. The hectare was used as a standard forest inventory unit, as in other studies of botanical diversity in Amazonia (ter Steege et al. 2013). Each inventory measured 10 m by 1000 m. All trees and lianas ≥10 cm at breast height (dbh) (as in Campbell et al. 1986; ter Steege et al. 2013) were measured, recorded and tagged with numbers for later collection and identification in the field. All collections of specimens were taken to the herbarium at the Universidade Federal do Oeste do Pará in Santarém for identification. In many cases taxa were identified to genus only and in a few cases only to family or even to indeterminate taxon and then grouped into morpho-categories for comparison with other individuals both in the terra firme and seasonally inundated forest inventories. Forest inventories were divided into 10 m X 25 m sampling units (SU), 40 in each inventory, for the purpose of estimating tree and liana species absolute density and to generate species/area curves (Fig. 4). We identified and compared species richness (number of species) and absolute dominance (measured by basal area in square meters) of all trees and lianas in the appropriate size class in both transects. The proportion of species shared by the two transects in relation to species total was compared using the Jaccard index (Sj). Data on dominance (total basal area occupied on the plot per species) were also compared in both transects. In addition, in order to estimate the age of the terra firme forest in the sample, we collected a large piece of xylem from the center near the base of the trunk of an enormous fallen Brazil nut tree about 20 m north of the terra firme transect. Its location was S 050 39.812′, W 0540 14.773′, and its diameter at roughly 1.7 m from its enormous base was approximately 150 cm (which is what its dbh would have been when the tree was standing). A C-14 date was obtained for this specimen (recorded as Iriri PN 1001, Beta – 498,822) from Beta Analytic, Inc. in July 2018.

Species/area curve of two inventories (“terra alta” and “baixão”)
We obtained data concerning abundance of fauna, especially in reference to target game species in local subsistence, as part of the Programa de Monitoramento Participativo da Biodiversidade (Participatory Monitoring of Biodiversity Program) under direction of the Instituto Chico Mendes de Biodiversidade (ICMBio), which has administrative oversight of the ESEC-TM. This includes photographic monitoring following the TEAM network protocol (www.teamnetwork.org) (per Beaudrot et al. 2016). We also conducted transect surveys with mapping of animals detected including their spoor and debris. We also carried out a parallel study mapping spoor during a rapid-assessment trip in March–April 2018 on several trails in use by the beiradeiros for collection of Brazil nuts, açaí (Euterpe oleracea), and access to swiddens and hunting grounds.
We conducted the archaeological survey and excavations over 3 weeks, in October–November 2017. We used forest inventory transects as a reference for sub-surface sampling. The sub-surface characterization of the site was undertaken using a posthole auger, which yielded basic site stratigraphy. Observations included how strata are characterized, the depth and horizontal extension of ADE, any shifting of ADE deposits for construction of structures, annotation of soil color, soil texture, and level of compactness. We also inspected the ground surface adjacent to each augered posthole.
Having initially identified archaeological remains, we delimited the spatial extent of archaeological evidence that extrapolated the inventory’s one hectare perimeter. This enabled us to understand the horizontal extent of the site. We refer to the archaeological site, which overlaps the inventories, as Grota do Cachorro. This stage involved further augering, by excavating 1 m-deep postholes along lines perpendicular to the original transect, as well as surface inspection.
On the basis of the information we gathered we selected locales for 1m2 test pit excavations. The test pits were excavated in 10 cm arbitrary levels, until an archaeologically sterile layer was encountered. This type of excavation allows for a more detailed observation of the vertical and horizontal distributions of archaeological evidence within the test pit and the limits between strata, as well as collection of samples. We retrieved lithic and ceramic remains, carbonised seeds, fragments of wood charcoal, and soil samples; the profiles of the excavations were drawn, photographed, and described. Areas surrounding both transects and the ADE patch were also inspected. We also mapped the seasonally flooded stream, as well as the Brazil nut trails and the Brazil nut trees situated along them (Fig. 2).
In addition to the forest inventories and archaeological excavations, we also surveyed local ethnobiological knowledge (TK, “traditional knowledge” or “traditional ecological knowledge) through a study of vocabulary for referencing the biota, specifically the trees, to understand prehistoric land and forest management practices. The freelist survey was carried out by asking adult beiradeiros in Portuguese to “tell me all the tree names you know.” We aimed to determine in an extensional way what is a tree in local parlance, and by inference, what are the most common, specific trees in the habitat; this was accomplished by free recall interviews (as in Balée and Cebolla Badie 2009). Thirty adult informants participated. All participants reside on the Iriri River along a riverine distance of more than 500 km, stretching across four degrees of latitude (02 to 06) and most reside inside the ESEC-TM. The data were cleaned as follows: spelling idiosyncrasies arising from free variation in speech were eliminated; attachment of noun classifier morphemes was retained if most respondents used them depending on the term; where most did not, the noun classifiers were also abandoned. For example, the postposed bound morpheme [−eiro/a] or [zeiro/a], which means “tree,” depending on whether the term is preceded by a consonant or not, is abandoned from words like “limeira” (lime tree) and “goloseira” (a sapotaceous tree). Species names are given where known. Many of the species names are taken from the inventory data. We calculated frequencies and salience values (based on Smith’s s) of all terms listed by informants.
Results
Forest Inventories
The total number of species from the two inventories was 282, with 32 shared species. There were 119 total species in riverine forest and 195 total species in terra firme forest, showing that the tree species richness of the terra firme is higher than the seasonally inundated forest (Fig. 4), which is unsurprising. Terra firme forests tend to be more diverse than floodplain forests due to environmental gradients of slope and seasonality. The number of shared species is 32, which yields an index of 11.34% sharedness by the Jaccard coefficient (the number of shared species divided by the total number of species in two different alpha-type plots, here 32/282). This value is comparatively low for adjacent forests in eastern Amazonia (e.g., Balée 1994), and suggests that the two forests of the two inventories represent different landscape types.
The most important trees in terms of absolute dominance (or total basal area) include several indicators of disturbance, especially Attalea speciosa (babaçu), Castilla ulei (caucho), Theobroma speciosum (nondomesticated cacao), and Handroanthus sp. (ipê) (Table 2). Each can be understood in terms of mechanisms of dispersal that help account for why they are located in terra firme forests disturbed at some distant point in the past by human agrarian activities (involving swidden gardening and agroforestry combined). What is perhaps most remarkable is the tremendous basal area occupied by babaçu palm (3.398 m2), which accounts for about 20% of all the basal area of all species on the plot. It is clearly a very dominant species in the entire habitat; this is no doubt true also of the Brazil nut, but the only Brazil nut on the plot were fallen trees, and these cannot be counted as individuals in the inventory. In any event, both species are for the most part confined to the terra firme forest. A. speciosa also has high values of relative density and relative frequency (Table 2). Relative density of a species is the number of subplots (out of a total of 40) that it occurs in divided by the total number of all such occurrences. The total number of occurrences in the terra firme forest inventory was 362 and in the seasonally inundated forest 346. All relative densities of all species sum to 1.0 (exclusive of rounding errors). The number of individuals in the terra firme inventory was 429, and in the seasonally inundated inventory, 473. Relative frequency of a species refers to the number of individuals of that species divided by the total number of individuals of all species on the plot. All relative frequencies sum to 1.0 (exclusive of rounding errors). Total basal area (the basal area of all species added together) was considerably higher in the seasonally inundated forest, at 20.303 square meters than in the terra firme forest at 16.881 m2. The most dominant species of the seasonally inundated forest are not useful or commercialized by way of comparison. The date obtained of the fallen Brazil nut’s tree xylem corroborates the charcoal in the soil of the site, with a date of approximately AD 1780, showing that the forest, though anthropogenic, is older than the forest peasants’ occupation since the late nineteenth century, by about 100 years at least. One of the conclusions to be drawn from this finding is that the forest peasants of the Iriri River have not degraded the prehistoric landscape; on the contrary, they are maintaining it. Brazil nut groves and babassu palm forests were useful to indigenous inhabitants, even if not commercialized by them.
Our results for the major dominant species in the baixão (Table 3) are significantly different from the comparable results for the terra firme (Table 2). For example, there are no disturbance indicators, such as Brazil nut, babaçu palm, cacao or wild cacao, or other trees found in anthropogenic forests elsewhere in eastern Amazonia (e.g., Balée 2013; Levis et al. 2018). In addition the only potential fruit trees are Caryocar sp. (the genus includes edible species), Lecythis pisonis (also known as monkey pot tree, which has small edible fruits and seeds), and Manilkara sp. (which produces small edible fruits as well as small amounts of sweet latex). In other words, these results suggest the baixão is indicative of a landscape not impacted, or minimally impacted, by human activity.
Fauna Survey
Fauna surveys on transects and trails of forest peasants inside the ESEC-TM indicated vestiges of 61 species of mammals and large ground dwelling birds. The vestiges include tracks, partially eaten fruits, feces, clawings, scratch marks, digging, hides, and feathers. Tapirs (Tapirus terrestris), deer (Mazama spp.), and peccaries (Tayassu pecari and Pecari tajacu)—which are large, preferred game species—are abundant in all the areas sampled, including those in close proximity to the dwellings of the forest peasants. Jaguars (Panthera onca), pacas (Cuniculus paca), agoutis (Dasyprocta leporina), monkeys, curassows, and guans (several species) are also abundant in the forests that surround the small homesteads dispersed along the Iriri; our observations imply substantial faunal diversity. Travelling from Altamira to the middle Iriri, the research team also observed numerous (although mostly unquantified) animals along the banks of the river.
Also during the March 2018 ascent of the river, we observed a herd of white-lipped peccaries (Tayassu pecari), which included 46 individuals, crossing the river, three kilometers downriver from the boundary between the Extractivist Reserve of the Iriri and the ESEC-TM. These and related sightings correspond to findings from previous fauna monitoring research done inside the extractive reserve, which suggests comparatively high faunal diversity.
Results of monitoring faunal diversity from the TEAM protocol indicate that the ESEC-TM exhibits the highest number of observations and species richness among all the TEAM network sites worldwide, including the 15 such sites located in the tropics (South America, Africa, and Southeast Asia) (Beaudrot et al. 2016; Carvalho Jr. 2017). Monitoring of game in the Rio Iriri and Riozinho do Anfrísio Extractive reserves, along with the forest peasants who live inside the ESEC-TM, shows that hunting activity is selective; the game taken consists almost exclusively of ungulates and large birds (Carneiro and Pezzuti, unpublished report).
Data that pertain to 4 years of game taken in the community of Rio Novo in the north of the ESEC-TM, show that hunting is in any case limited and sporadic (with only 58 animals taken in 4 years of study) and that it is effected opportunistically, usually in connection with canoe trips along the Rio Iriri for fishing or Brazil nut gathering (Table 4). Most of the species taken, such as pacas, capyvaras (Hydrochoerus hydrochaeris), tapirs, and curassows (especially Crax spp.) are found along riverbanks (Carneiro and Pezzuti 2009).
Archaeological Surveys and Excavations
Fieldwork consisted mainly of excavation of test pits and augering of postholes. We excavated 69 auger postholes, 40 of which followed the terra firme transect and 24 of which followed the riverine forest transect. We dug an additional five auger postholes within the ADE at Grota do Cachorro. The augered postholes that followed the terra firme forest transect and that roughly accompany the Brazil nut tree trail point to archaeological material up to SU 08 (Sampling Unit 08, that is, about 200 m from beginning of the one kilometre-long transect—there are 40 sampling units numbered consecutively 00–40, with 00 the beginning and 40 the end of the transect--in linear arrangement on each of the two one-hectare inventory plots). We observed both lithics and ceramics at SU 00, 01, and 02 (along the transects shown in Fig. 2). Top soil color varied progressively from very dark at SU 00 to dark brown at SU 08 and 09 (Fig. 5a), after which no further ceramic remains were identified (Fig. 5b). The absence of archaeological remains indicated this to be the NE limit of prehistoric settlement at Grota do Cachorro.

a Augured posthole by terra firme transect point SU 00. The soil heap farthest to the right represents the most superficial levels; the left one represents the deepest levels. b Augured posthole by riverine forest transect point SU 02. The soil heap farthest to the right represents the most superficial levels; the left one represents the deepest levels
As expected, augered postholes along the riverine forest transect present features consistent with seasonal flooding, with soil color ranging from light gray to gray and light brownish-gray. We found pottery at approximately 30 cm depth at one augered posthole only (at SU 02), near the Igarapé do Bala itself (Fig. 5a).
We excavated five postholes and inspected another 30 points, plotted by GPS, outside the forest inventories transects. In relation to the postholes, of interest is the excavation of point N1000/E950, located 50 m SW of terra firme SU 00, which presented a lighter colour soil (brown) and limited occurrence of pottery. At approximately 70 cm depth we encountered a line of charcoal suggesting a possible burning episode and in the 80-100 cm levels we retrieved knapped lithics.
Following posthole excavations along the transects and within the ADE patch, we excavated two 1m2 test pits. Excavation 1 is situated adjacent to the first sampling unit (SU 00) of the terra firme transect, where the posthole excavation indicated a large amount of archaeological pottery and ADE up to c. 40 cm in depth (Fig. 6a and b). Excavation 1 was therefore dug in order to collect the greatest possible amount of ceramic and lithic materials for an initial characterization of these remains and for subsequent comparison with archaeological materials in the wider region (Toney 2012; Honorato de Oliveira 2015; Garcia 2017; Rocha 2017). A charcoal sample identified to be a carbonized babaçu (Attalea speciosa) seed extracted at 24 cm depth was dated to 410 ± 30 C14 BP. This layer contained the greatest quantity of pottery, as well as carbonized botanical material and terra preta, which is the type of ADE found in the archaeological site.

a Test pit near the terra firme forest transect and inside the archaeological site; note darker soil colour near top, indicative of prehistoric human settlement. b Test pit near terra firme forest transect outside nuclear zone of prehistoric settlement
Excavation 2 is located near posthole N950/E1000, where a line of charcoal at approximately 70 cm depth as well as buried lithic vestiges that were not associated with ceramics or ADE had been identified (Fig. 6b). A small quantity of ceramics associated with lithics was evidenced up to 80 cm depth. In layer III (situated between 80 and 98 cm), we noted an increase in the amount of flaked milky quartz and absence of ceramics. We further noted a somewhat darker soil colour (10YR 4/6, dark yellowish brown), if compared to layers above (10YR 5/4, yellowish brown) and below (10YR 5/6, yellowish brown). This archaeological layer was dated to 3850 ± 30 C14 BP (4.300–4.085 cal BP).
Finally, inspection around the site suggests past indigenous activity. We observed stone polishing features at the southern boundary of the terra preta, in the creek, and on the banks of the Igarapé do Bala. The existence of lithic raw materials similar to those observed in finished pieces at other archaeological sites surveyed during the expedition points to intensive production of stone axes in the past. These were probably used in forest management long before the arrival of iron tools from Luso-Brazilian populations. The presence of fixed polishing features within the Grota do Cachorro creek suggests seasonality of stone axe production, since these locales are submerged during the rainy season, as was observed during the March–April 2018 reconnaissance visit. The archaeological data in summary suggest a village of roughly one hectare in size on the edge of the Igarapé do Bala, situated atop terra firme near the beginning of a Brazil nut grove trail. This means the Brazil nut trees were likely cultivated purposefully near but far enough away from permanent habitations in order to pose no danger—from falling fruit capsules, branches, and entire gigantic trees— to the indigenous people and their dwellings in the prehistoric settlement.
Freelists and Traditional Knowledge
The freelists identified 334 distinctive tree names (Table 5) of which 155 are considered valid because they were mentioned by at least two respondents. The most salient trees on the list tend to be tall, enormous hardwoods (Appendix E), and this fact instantiates what is probably a universal feature of the life form ‘tree’ in diverse languages; the pattern is found elsewhere in lowland South American traditional societies (Balée and Cebolla Badie 2009). As such the freelist data are further evidence of the traditional status—in terms of TK or TEK--of beiradeiros in the middle Iriri basin.
In addition, some of these hardwoods together with numerous fruit trees characteristic of anthropogenic forests are also among the most important taxa named by local folk generic terms, such as Brazil nut, Hymenaea spp., Caryocar sp., Sapotaceae sp., Spondias sp., Lecythis pisonis, and the açaí and bacaba palms (Table 5). Palms are only marginally considered trees, therefore, which further corresponds to traditional knowledge reported elsewhere in Amazonia (e.g., Berlin 1992) given that unlike most “trees” in traditional ethnobiological classification systems, palms are monocots lacking growth rings, woody branches, and many of them, in the specific area of study, grow in ways trees normally do not, such as in caespitose format (as with Euterpe oleracea) or stemlessly as with palmetto and palmetto-like individuals (e.g., Geonoma spp.).
According to the inventory data, the richness of trees and lianas together with the dominance values of disturbance indicator species is higher in the terra firme than in the baixão of the study area. This result suggests that the most impactful human activity on the forest occurs in the terra firme rather than in the baixão, which lacks most indicators of human agency found in the terra alta (or terra firme), e.g., no evidence of burning. We observed evidence of more targeted, seasonal activities, such as stone axe production in the prehistoric past. We can postulate further that some of the species on the terra firme originated in the riverine forest (such as Hevea brasiliensis, Handroanthus spp.) and that the terra firme is in part diverse because of factors that induce differences in biotas between the two areas, such as soil water saturation (Pires 1976; McMenamin and McMenamin 1996; Ivanauskas et al. 1997; Montagnini and Muñiz-Miret 1999), even if the inundation in this particular case is only seasonal.
Discussion and Conclusions
Primary Landscape Transformation
The archaeological evidence we identified and present-day beiradeiro activities such as use of small (apparently natural) mounds found inside the seasonally inundated forest for hunting and collecting activities, together with their management practices and deep ecological knowledge suggest a continuous use of this area from the distant past to the present. In other words, species richness and dominance are maintained as part of a general conservation of the indigenous forest legacy of past peoples, to whom the beiradeiros are heirs. The activity of these people inside this forest during the last 100 years or so constitutes the continuity of landscape transformation engineered long ago. The primary landscape transformation dates from 410 ± 30 C14 BP (or even earlier, given the previous occupation of 3850 ± 30 C14 BP or 4300–4085 cal BP), or from about the beginning of the seventeenth century, though the beiradeiro culture and its forebears are present only within the last 130 years.
In one remarkable sense, there appears to be a relationship between the Grota do Cachorro site’s probable habitation area (inferred from the presence of archaeological artifacts and ADE) and the location of the Brazil nut trees and the trail through the Brazil nut grove. There appears to have been a conscious division of space into settlement and possibly herbaceous cultivars versus an agroforestry zone with disturbance indicator (and in some cases domesticated) trees, such as Brazil nut, babaçu palm, and cacao. The greatest concentration of Brazil nut trees is found adjacent to the outer limits of the ADE patch, which seems to indicate a potential correlation between ADE formation and the formation of the Brazil nut tree grove, while the Brazil nut tree trail goes well beyond the limits of what is likely the original area of agrarian habitation in prehistory. Probably, Brazil nut trees were set back from the probable location of the prehistoric village (Levis et al. 2018), which would have been located near the Igarapé do Bala.
Secondary Landscape Transformations and Beiradeiro TK
The beiradeiros who exploit the area for Brazil nuts (and in the recent past for tonka bean, cacao, and rubber) are not essentially modifying the original framework of a prehistoric agrarian and agroforestry system. Rather, they maintain it precisely because they utilize the area, and consciously seek to protect and encourage its valuable non-timber forest resources, especially Brazil nuts at present.
Beiradeiros at an earlier time extracted high quality rubber from both the baixão and the terra firme forest (it occurs in both inventories) (Schwartzman et al. 2013). Their local TK at present exhibits an accurate chronology more or less of the age of the forest, and of earlier landscape transformations. That is because they can explain the different technologies of the past that impacted the landscape one witnesses today. We attested that they note the type of knife used for scoring rubber trees, such as the u-shaped Amazonian knife, which has been in use since about the 1930s. Informants claim its use makes a different type of wound on the rubber tree than the hatchet (called regionally the machadinho, or ‘little axe’), which was used by the first wave of immigrant rubber tappers to the region of the middle Iriri in the period from about 1890 to 1920. The hatchet wounds make distinctive knots not seen to be congruent with the result of the more modern knife, and hence indicates forest usage by beiradeiros since the first arrival of permanent Luso-Brazilian colonization of the area (specifically by rubber tappers originating in Northeast Brazil).
Secondary landscape transformation probably also includes domesticated and semidomesticated cacao trees—specifically Theobroma speciosum, the most dominant, frequent, and densely distributed species on the terra firme site, and has beans and fruit similar to domesticated cacao (Table 2). Betendorf also reported the abundance of cacao groves, mainly Theobroma speciosum, in the late 1600s. Caucho trees are also one of the most dominant species in the well-drained forest of the Rio Iriri basin where the study was conducted. These observations indicate an indirect effect of TK and cultural management of the forest and secondary landscape transformations that have the effect of managing the original, primary landscape transformation, which probably involved the establishment of a managed forest in the first place by prehistoric indigenes.
Folk knowledge, much of it derived from indigenous ancestors who occupied the region, affects the secondary landscape transformations that have not degraded the forest in spite of more than 100 years of forest peasant occupation. Levis et al. (2018: 15) observe that “modern Amazonian peoples who live on pre-Columbian settlements seem to have inherited indigenous knowledge.” Indeed, beiradeiro folk generic terms for the most important trees (Table 5) approximate the richness of indigenous nomenclature, and indicate deep ecological knowledge of the Iriri River forests.
Beiradeiro folk knowledge and its probable indigenous roots are illustrated by their knowledge of the uses of Minquartia guianensis (acariquara, whose wood contains silicates resistant both to termites and to decomposition from flooding), a tree widely used in house posts in the Iriri and elsewhere. One male informant, aged 64 at the time of research, remarked that “nem terra, nem água come ...” (‘neither land nor water consume it’). Tupian indigenous societies of Amazonia, such as the Ka’apor far to the east recognize the same properties of the acariquara (Balée 1994). In August 2017, we observed the informant demonstrate that an above-ground root of the samaúma tree (Ceiba pentandra) contained several liters of potable water. The samaúma is found both in Atlantic Coastal Forest and Amazonia. This traditional knowledge could have derived from historical antecedents in the indigenous Northeast of Brazil or in indigenous Amazonia or both.
New Avenues for Historical Ecology
The evidence we presented here points to the forest activities of beiradeiros—as an ethnograpically and ethnohistorically real and agentive social group-- being sustainable. The beiradeiros exemplify the paradigmatic concept of forest peasants living sustainably in a heavily forested Amazonian biome rich in biodiversity (Carneiro da Cunha and Almeida 2000; cf. Peres 2011). Because virtually all Amazon strictly protected areas are inhabited by humans, traditional occupation by forest peasants is, in this sense, already compatible with protection of biodiversity. Our results thus far show that faunal richness and abundance in the inhabited forest is high. In addition, as in the three extractive reserves of the Terra do Meio (Fig. 1), beiradeiros’ hunting and collecting treks typically do not exceed a radius of about ten kilometers into the forest, away from the river. If one considers this to be standard for the entire mosaic of reserves in the Terra do Meio, the 3.4 million hectares of the ESEC – TM group is surrounded on three sides, for some 80% of its perimeter, by indigenous and beiradeiro occupation that provides the basis for potentially preventing invasion and forest destruction by outside settlers’ incursions, such as wildcat land grabbers (grileiros) and large soybean farms and cattle ranches. From this perspective, supporting the beiradeiros in the ESEC-TM with transportation and communication infrastructure, social services, and investment in technology and capacity for income-generating activities would better protect the biodiversity of the strict nature reserve, than would their removal.
Our findings suggest the need for a more nuanced approach to human habitation in Amazonian strictly protected areas, and for further multidisciplinary field research in historical ecology comparing diversity in both inhabited and uninhabited forests of protected areas, examining archeological as well as ecological and ethnographic evidence to better understand the extent and timescale of ancient and modern occupation of forests, and the legacy of indigenous knowledge in forest peasant communities. Hisotrical ecology, serving as the rubric under which to assemble all of these kinds of evidence, can contribute to more effective forest conservation.
Change history
28 May 2021
The original version of this paper was updated to present the correct family name of Dr. Vinicius Honorato de Oliveira.
References
Alarcon, D.F., and Torres, M. (2014). “Não tem essa lei no mundo rapaz”: A Estaçāo Ecológica da Terra do Meio e a resistência dos beiradeiros do alto rio Iriri. Associaçāo dos Moradores de Reserva Extrativista Riozinho do Anfrísio (Amora) and Instituto Socioambiental (ISA), Altamira, Brazil.
Alden, D. (1976). The significance of cacao production in the Amazon region during the late colonial period: An essay in comparative economic history. Proceedings of the American Philosophical Society 120(2): 103–135.
Almeida, M.W.B., Guerrero, N.R., Francesco, A., Postigo, A., Rocha, B.C., Nepomuceno, I., Doblas, J., Carneiro da Cunha, M., Salazar, M., Amaral, M., Torres, M., Santos, R.R., Scoles, R., Rezende, R.S., Honorato, V., and Balée, W. (2018). Laudo pericial sobre a situaçāo das famílias residentes no interior da Estação Ecológica da Terra do Meio (Expert report). São Paulo.
Alvares, C. A., Stape, J. L., Sentelhas, P. C., Gonçalves, J. L. M., and Sparovek, G. (2013). Köppen’s climate classification map for Brazil. Meterologische Zeitschrift 22: 711–728. https://doi.org/10.1127/0941-2948/2013/0507.
Arroyo-Kalin, M. (2016). Landscaping, landscape legacies, and landesque capital in pre-Columbian Amazonia. In Isendahl, C., and Stump, D. (eds.), The Oxford handbook of historical ecology and applied archaeology, Oxford University Press, Oxford. https://doi.org/10.1093/oxfordhb/9780199672691.013.16.
Balée, W. (1994). Footprints of the Forest: Ka’apor Ethnobotany –the historical ecology of plant utilization by an Amazonian people, Columbia University Press, New York.
Balée, W. (2013). Cultural forests of the Amazon: A historical ecology of people and their landscapes. University of Alabama Press, Tuscaloosa.
Balée, W. (2018). Brief review of historical ecology. Les Nouvelles de l’Archéologie 152(juin): 7–10. https://doi.org/10.4000/nda.4150.
Balée, W., and Cebolla Badie, M. (2009). The meaning of “tree” in two different Tupí-Guaraní languages from two different Neotropical forests. Amazônica 1: 96–135. https://doi.org/10.18542/amazonica.v1i1.140.
Balée, W., Schaan, D. P., Whitaker, J.A., and Holanda, R. (2014) Florestas antrópicas no Acre: Inventário florestal no geoglifo Três Vertentes, Acrelândia. Amazônica - Revista de Antropologia 6(1):142–169.
Barreto Filho, H. T. (2004). Notas para uma história social das áreas de proteção integral no Brasil. In Ricardo, F. (ed.), Terras Indígenas & Unidades de Conservação da natureza: o desafio das sobreposições, Instituto Socioambiental, São Paulo.
Beaudrot, L., Ahumada, J. A., O’Brien, T., Alvarez-Loayza, P., Boekee, K., Campos-Arceiz, A., Eichberg, D., Espinosa, S., Fegraus, E., Fletcher, C., Gajapersad, K., Hallam, C., Hurtado, J., Jansen, P. A., Kumar, A., Larney, E., Lima, M. G. M., Mahony, C., Martin, E. H., McWilliam, A., Mugerwa, B., Ndoundou-Hockemba, M., Razafimahaimodison, J. C., Romero-Saltos, H., Rovero, F., Salvador, J., Santos, F., Sheil, D., Spironello, W. R., Willig, M. R., Winarni, N. L., Zvoleff, A., and Andelman, S. J. (2016). Standardized assessment of biodiversity trends in tropical forest protected areas: The end is not in sight. PLOS Biolog. 14: e1002357. https://doi.org/10.1371/journal.pbio.1002357.
Berlin, B. (1992). Ethnobiological classification: Principles of categorization of plants and and animals in traditional societies, Princeton University Press, Princeton.
Betendorf, J. F. (1910). Chronica da Missão dos Padres da Companhia de Jesus no Estado do Maranhão. Revista do Instituto Historico e Geographico Brazileiro, Tomo LXXII, Parte I, Rio de Janeiro.
Brasil. (2000). Lei n. 9.985, de 18 de julho de 2000. Institui o Sistema Nacional de Conservação e dá outras providências. Diário Oficial da União, Brasília.
Brasil. (2005). Decreto s/n. 2005, 17 de fevereiro. Cria a Estação Ecológica da Terra do Meio, nos municípios de Altamira e São Félix do Xingu, Estado do Pará, e dá outras providências. Presidência da República, Brasília.
Brookfield, H.C. (1984). Intensification revisited. Pacific Viewpoint 25(1):15–44.
Campbell, D. G., Daly, D. C., Prance, G. T., and Ubirajara, N. M. (1986). Quantitative ecological inventory of terra firme and várzea tropical forest on the Rio Xingu, Brazilian Amazon. Brittonia 38: 369–393. https://doi.org/10.2307/2807085.
Carneiro da Cunha, M., and Almeida, M. W. B. (2000). Indigenous people, traditional people, and conservation in the Amazon. Daedalus 129: 315–338.
Carneiro, C., and Pezzuti, J. C. B. (2009). Diagnóstico da caça e da pesca nas Reservas Extrativistas da Terra do Meio. WWF, Belém (unpublished report).
Carvalho Jr. E. (2017). Monitoramento de vertebrados terrestres na Estação Ecológica da Terra do Meio com base no protocolo avançado do Programa Monitora. Technical report, Atibaia.
Clement, C. R., Denevan, W. M., Heckenberger, M. J., Junqueira, A. B., Neves, E. G., Teixeira, W. G., and Woods, W. (2015). The domestication of Amazonia before European conquest. Proceedings of the Royal Society B 282: 1–9.
Eletrobras (2009). AAI –Avaliação Ambiental Integrada Aproveitamentos Hidrelétricos da Bacia Hidrográfica do Rio Xingu Volume I. Eletrobras, São Paulo.
Garcia, L. L. W. G. (2017). Paisagens do médio-baixo Xingu. Arqueologia, Temporalidade e Historicidade. Ph.D. dissertation. Museu de Arqueologia e Etnologia, Universidade de São Paulo, São Paulo.
Grenand, P. (1982). Ainsi parlaient nos ancêtres: Essai d’éthnohistoire “Waỹapi”. L’Office de la Recherche Scientifique et Technique Outre-Mer (Orstom), Paris.
Hecht, S. B. (2013). The scramble for the Amazon and the “lost paradise” of Euclides da Cunha, The University of Chicago Press, Chicago.
Honorato de Oliveira, V. E. (2015). Shatters among Sherds: A study of lithic assemblages of the upper Tapajós River. MA Dissertation. Institute of Archaeology, University College London.
ICMBio (Instituto Chico Mendes de Conservação da Biodiversidade) (2015). Plano de Manejo da Estação Ecológica da Terra do Meio, ICMBio, Brasília.
Ivanauskas, N. M., Rodrigues, R. R., and Nave, A. G. (1997). Aspectos ecológicos de um trecho de floresta de brejo em Itatinga, SP: Florística, fitossociologia e seletividade de espécies. Revista Brasileira de Botânica, São Paulo 20: 139–153. https://doi.org/10.1590/S0100-84041997000200005.
Lei n. 9.985, de 18 de julho de 2000. (2000). Institui o Sistema Nacional de Unidades de Conservação da Natureza e dá outras providências. Diário Oficial da União, Brasília.
Levis, C., Souza, P. F., Schietti, J., Emilio, T., Pinto, J. L. P. V., Clement, C. R., and Costa, F. R. C. (2012). Historical human footprint on modern tree species composition in the Purus-Madeira interfluve, Central Amazonia. PLOS One 7: e48559. https://doi.org/10.1371/journal.pone.0048559.
Levis, C., et al (2017). Persistent effects of pre-Columbian plant domestication on Amazonian forest composition. Science 355: 925–931. https://doi.org/10.1126/science.aal0157.
Levis, C., Flores, B. M., Moreira, P. A., Luize, B. G., Alves, R. P., Franco-Moraes, J., Lins, J., Konings, E., Peña-Claros, M., Bongers, F., Costa, F. R. C., and Clement, C. R. (2018). How people domesticated Amazonian forests. Frontiers in Ecology and Evolution 5: 171. https://doi.org/10.3389/fevo.2017.00171.
Lima, H. P. (2008). História das Caretas: A Tradição Borda-Incisa na Amazônia Central (Doctoral dissertation, Universidade de São Paulo, Museu de Arqueologia e Etnologia, São Paulo, Brasil). Retrieved from http://www.teses.usp.br/teses/disponiveis/71/71131/tde-22092008-125315/pt-br.php
Machado, J. S. (2005). Montículos artificiais na Amazônia Central: Um Estudo de Caso do Sítio Hatahara (Master’s thesis). Universidade de São Paulo, Museu de Arqueologia e Etnologia, São Paulo.
Maxwell, K. R. (1973). Conflicts and conspiracies: Brazil and Portugal, 1750–1808, Cambridge University Press, Cambridge.
McMenamin, M. A. S., and McMenamin, D. L. S. (1996). Hypersea: Life on land. Columbia University Press, New York.
McMichael, C. N. H., Piperno, D. R., Bush, M. B., Silman, M. R., Zimmermann, A., Raczka, M. F., and Lobato, L. C. (2012). Sparse pre-Columbian human habitation in western Amazonia. Science 336: 1429–1431. https://doi.org/10.1126/science.1219982.
McMichael, C. H., Palace, M. W., Bush, M. B., Braswell, B., Hagen, S., Neves, E. G., Silman, M. R., Tamanaha, E. K., and Czarnecki, C. (2014). Predicting pre-Columbian anthropogenic soils in Amazonia. Proceedings of the Royal Society B: Biological Sciences. 281(1777): 2–9. https://doi.org/10.1098/rspb.2013.2475.
Montagnini, F., and Muñiz-Miret, N. (1999). Vegetation and soils of tidal floodplains of the Amazon estuary: A comparison of Várzea and Terra Firme forests in Pará, Brazil. Journal of Tropical Forest Science 11: 420–437.
Moraes, C. P. (2006). Arqueologia na Amazônia Central vista de uma perspectiva da região do Lago do Limão (Master’s thesis, Universidade de São Paulo, Museu de Arqueologia e Etnologia, São Paulo, Brasil). Available: http://www.teses.usp.br/teses/disponiveis/71/71131/tde-15052007-112151/es.php
Nelson, A., and Chomitz, K. M. (2011). Effectiveness of strict vs. multiple use protected areas in reducing tropical Forest fires: A global analysis using matching methods. PLOS One 6(8): e22722. https://doi.org/10.1371/journal.pone.0022722.
Nepstad, D., Schwartzman, S., Bamberger, B., Santilli, M., Ray, D., Schlesinger, P., Lefebvre, P., Alencar, A., Prinz, E., Fiske, G., and Rolla, A. (2006). Inhibition of Amazon deforestation and fire by parks and indigenous lands. Conservation Biology 20: 65–73. https://doi.org/10.1111/j.1523-1739.2006.00351.x.
Neves, E. G. (2000). Levantamento Arqueológico na Área de Confluência dos Rios Negro e Solimões, Estado do Amazonas. Report, FAPESP-MAE, Universidade de São Paulo, São Paulo.
Nimuendaju, C. (1948). Tribes of the lower and middle Xingu. In: Steward, J. (ed.). Handbook of south American Indians, vol. 3, the tropical Forest tribes. Smithsonian Institution, Bureau of American Ethnology, bulletin 143, pp. 209–243. U.S. Government Printing Office, Washington, D.C.
Nimuendaju, C. (1980) [orig. 1944]. Mapa etno-histórico do Brasil e regiões adjacentes. IBGE, Rio de Janeiro.
Nimuendaju, C. (2004). In pursuit of a past Amazon: Archaeological researches in the Brazilian Guyana and in the Amazon region (Rydén, S. and per Stemborg, eds.). Etnologiska studier 45, Gothenburg.
Peres, C. A. (2011). Conservation in sustainable-use tropical forest reserves. Conservation Biology 25: 1124–1129.
Perota, C. (1979). Pesquisas de campo realizadas na bacia do rio Xingu como parte do Programa Nacional de Pesquisas Arqueológicas da Bacia Amazônica (PRONAPABA). Report, CNPq - IPHAN – Universidade Federal do Espírito Santo, Vitória.
Piperno, D. R., McMichael, C., and Bush, M. B. (2015). Amazonia and the Anthropocene: What was the spatial extent and intensity of human landscape modifications in the Amazon basin at the end of prehistory? The Holocene 25: 1588–1597. https://doi.org/10.1177/0959683615588374.
Piperno, D. R., McMichael, C., and Bush, M. B. (2019). Finding forest management in prehistoric Amazonia. Anthropocene. https://doi.org/10.1016/j.ancene.2019.100211.
Pires, J.M. (1976). Aspectos ecológicos da floresta amazônica. Anais II Congresso Brasileiro de Florestas Tropicais. Coleção Mossoroense 65:235–287.
Porter-Bolland, L., Ellis, E. A., Guariguata, M. R., Ruiz-Mallén, I., Negrete-Yankelevich, S., and Reyes-García, V. (2012). Community managed forests and forest protected areas: An assessment of their conservation effectiveness across the tropics. Forest Ecology and Management 268: 6–17. https://doi.org/10.1016/j.foreco.2011.05.034.
Rocha, B. C. (2017). Ipi Ocemumuge: A Regional Archaeology of the Upper Tapajós River. PhD thesis, Institute of Archaeology, University College London.
Rodrigues, A. D., and Cabral, A. S. A. (2012). Tupían. In: Campbell, L., and V. Grondona (eds.), The indigenous languages of South America. Mouto de Gruyter, Berlin.
Roosevelt, A. C., Douglas, J. E., Amaral, A. M., Imazio da Silveira, M., Barbosa, C. P., and Barreto, M. (2009). Early hunter-gatherers in the terra firme rainforest: Stemmed projectile points from the Curuá goldmines. Amazônica 1(2): 442–483. https://doi.org/10.18542/amazonica.v1i2.296.
Santilli, J. (2005). Socioambientalismo e novos direitos. Proteção jurídica à diversidade biológica e cultural, Peirópolis, São Paulo.
Schroth, G., Coutinho, P., Moraes, V. H. F., and Albernaz, A. L. (2003). Rubber agroforests at the Tapajós river, Brazilian Amazon – Environmentally benign land use systems in an old forest frontier region. Agriculture, Ecosystems and Environment 97: 151–165. https://doi.org/10.1016/S0167-8809(03)00116-6.
Schwartzman, S., Alencar, A., Zarin, H., and Souza, A. P. S. (2010). Social movements and large-scale tropical forest protection on the Amazon frontier: Conservation from chaos. The Journal of Environment and Development 19(3): 274–279. https://doi.org/10.1177/1070496510367627.
Schwartzman, S., Villas-Bôas, A., Ono, K. Y., Fonseca, M. G., Doblas, J., Zimmerman, B., Junqueira, P., Jerozolimski, A., Salazar, M., Junqueira, R. P., and Torres, M. (2013). The natural and social history of the indigenous lands and protected areas corridor of the Xingu River basin. Philosophical Transactions of the Royal Society B: Biological Sciences 368: 1–12. https://doi.org/10.1098/rstb.2012.0164.
Shanin, T. (1984). Peasantry as a political factor. In Shanin, T. (ed.), Peasant and peasant societies, Penguinbooks, Middlesex, pp. 238–263.
Shanin, T. (2008) Lições camponesas. In: Paulino, E. T., and J. E. Fabrini (eds.). Campesinato e territórios em disputa. Expressão Popular: 23–47.
Snethlage, E. (2002) [orig. 1910]. A travessia entre o Xingú e o Tapajós. Coleção Documentos da Amazônia, v. 98. Governo do Estado do Amazonas and Secretaria de Cultura, Manaus.
Tamanaha, E.K. (2012). Ocupação policroma no Baixo e Médio Rio Solimões, Estado do Amazonas. MA thesis., Museu de Arqueologia e Etnologia, Universidade de São Paulo, São Paulo). Available: http://www.teses.usp.br/teses/disponiveis/71/71131/tde-05062012-104830/fr.php
ter Steege, H., et al (2013). Hyperdominance in the Amazonian tree flora. Science 342: 1243092. https://doi.org/10.1126/science.1243092.
Terborgh, J. (1999). Requiem for nature, Island Press, Washington, D.C.
Thieme, I. (1993). Karl von den Steinen: Vida e obra. In Coelho, V. P. (ed.), Karl von den Steinen: Um século de antropologia no Xingu, Edusp, São Paulo.
Toney, J.R. (2012). The product of labor: Pottery Technology in the Upper Xingu, southern Amazon, Brazil, A.D. 700–1770. PhD dissertation, Department of Anthropology, University of Florida, Gainesville.
Torres, M. G. (2008). A beiradeira e o grilador: ocupação no oeste do Pará (Master’s thesis, Universidade de São Paulo, Faculdade de Filosofia, Letras e Ciências Humanas, São Paulo. Available: http://www.teses.usp.br/teses/disponiveis/8/8136/tde-27112008-132446/pt-br.php
Villas-Bôas, A., Sanches, R. A., and Scaramuzza, C. A. M. (2003). Estudos preliminares e formulação de uma proposta técnica para a implantação de um mosaico de Unidades de Conservação no Médio Xingu, Instituto Socioambiental, São Paulo.
Villas-Bôas, A., Guerrero, N. R., Junqueira, R. G. P., and Postigo, A. (eds.) (2017). Xingu: história dos produtos da floresta, Instituto Socioambiental, São Paulo.
Viveiros de Castro, E. (1992). From the enemy’s point of view: Humanity and divinity in an Amazonian society. Trans. by Catherine V. Howard. University of Chicago Press, Chicago.
Wagley, C. (2014). [orig. 1953]. Amazon town: A study of human life in the tropics. Oxford University Press, New York.
Acknowledgements
The authors gratefully acknowledge expert assistance provided by Juan David Tovar Durán and María Alejandra Buitrago Aristizabal in determination of many of the trees and lianas in the herbarium. Thanks are due to Juan Doblas for elaboration of the map in Fig. 1. We acknowledge Leandro Giacomin for help with securing access to the herbarium of the Universidade Federal do Oeste do Pará (Ufopa) and for his assistance in finding key individuals to facilitate the research reported herein both in the field and in the herbarium. We thank José Alves Gomes and his family for assistance with logistics in the field; we are further grateful to Jony Martins Oliveira for assistance with tree and liana collections on the inventory sites discussed herein. We thankMarcelo Salazar of Instituto Socioambiental (ISA-Altamira) for advice on technology and logistical matters. Financial and logistical support for the project were offered by the Instituto Socioambiental (ISA – Altamira); we are indebted to Environmental Defense Fund (EDF) for major funding of the fieldwork (proposal number 7601098). The authors thank ICMBio (Chico Mendes Institute of Biodiversity Conservation), IPHAN (Institute of Historical and Artistic Patrimony), Universidade Federal do Oeste do Pará (Ufopa) and Tulane University for administrative endorsement of the project.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Human subjects research reported in this article was approved by the Tulane University Institutional Review Board (reference no. 1101293); in Brazil, plant collections reported herein were approved by the Instituto Chico Mendes de Conservaçāo da Biodiversidade (ICMBio) of the Ministério do Ambiente (MMA) (no. 59656–2); and the archaeological research reported herein was approved by the Instituto de Patrimônio Histórico e Artístico Nacional (Iphan) by Portaria no. 35 of 13 July 2017 for Processo no. 01492.000380/2017–15, as amended by Portaria 0090314–60 of 23 October 2017. The field research reported herein was funded by Environmental Defense Fund (grant no. 76010980).
Conflict of Interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Balée, W., Honorato de Oliveira, V., dos Santos, R. et al. Ancient Transformation, Current Conservation: Traditional Forest Management on the Iriri River, Brazilian Amazonia. Hum Ecol 48, 1–15 (2020). https://doi.org/10.1007/s10745-020-00139-3
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10745-020-00139-3