
Abstract
Cold stress disturbs numerous physiological and biochemical processes, restricting plant growth and productivity. In the current study, the protective role of exogenous serotonin in alleviating cold stress was investigated in rapeseed (Brassica napus L.) seedlings. The rapeseed seedlings were pretreated with different concentrations (0, 0.01, 0.02, 0.03, and 0.04 g L− 1) of serotonin solution and then were exposed to cold stress (4, 2, 0, and − 2 °C). The results indicated that the pretreatment of serotonin significantly increased the survival rate. Mainly, 0.03 g L− 1 of serotonin increased the survival rate by 75% compared to control conditions. The physiological and biochemical indexes and the expression of cold tolerance-related genes were analyzed in the seedlings pretreated with 0.03 g L− 1 of serotonin. The contents of proline (PRO), soluble sugar (SS), and soluble protein (SP); and the activities of antioxidant defense such as catalase (CAT), peroxidase (POD), and superoxide dismutase (SOD) were significantly increased by exogenous serotonin under cold stress. Supplemented serotonin significantly increased the expression of SOD, COR6.6, COR15, and CBFs genes under cold stress.Overall, our results indicate that the optimal concentration (0.03 g L− 1) of exogenous serotonin maintained the osmotic potential balance in cells under cold stress by increasing the content of osmotic regulatory substances (SS, SP, and PRO), improving the scavenging ability of reactive oxygen species (ROS), increasing the antioxidant enzyme activities (CAT, POD, and SOD) and the transcriptional level of cold stress-related genes, helping rapeseed seedlings to cope with the cold stress. Therefore, serotonin-induced regulatory interactions between physiological and biochemical processes and the elevated expression of stress-associated genes may be a beneficial technique for cold stress tolerance in plants.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Climate change gives rise to several abiotic stresses, including extreme temperatures, heavy metals, drought, and soil salinity are major adverse environmental conditions that plants often encounter (Raza Ali et al. 2019, 2020a). Among them, cold stress or low temperature [(chilling (0–15°C) and freezing temperature (< 0°C)] meaningfully act as a significant threat to agricultural production (Agarwal et al. 2006; Chinnusamy et al. 2006). Under the freezing stress, extracellular icicles induce cell water to convent to extracellular space, causing cellular dehydration, mechanical damage, and even death of plants (Pearce 2001; McCully et al. 2004; Li et al. 2018a). Low-temperature stress disturbs the equilibrium between the production and scavenging of reactive oxygen species (ROS) in plants (Miller et al. 2008; Hasanuzzaman et al. 2020). Whereas the accumulation of ROS impaired large molecules such as proteins, lipids, DNA, and cell membrane, even leading to the permanent damage of cells (Miller et al. 2008; Hasanuzzaman et al. 2020). Cold stress not only reduces plant photosynthesis, cell membrane fluidity, and basal metabolism but also inhibits plant growth and development (Pearce 2001; Ding et al. 2019).
Over the past few years, several plant growth regulators were widely used to increase stress tolerance in plants (Li et al. 2018b; Strydhorst et al. 2018). Serotonin has antioxidative and growth-inducing properties, therefore demonstrating useful for stress adaptation and plant physiology. Complex molecular connections of serotonin and stress-responsive genes have proposed its antagonistic nature (Kaur et al. 2015). Serotonin was detected in plants’ vegetative and reproductive tissues (Mukherjee et al. 2014); the precursor substance of melatonin is widely found as an indole-tryptamine in plants and animals (Radhakrishnan and Lee 2013; Shi et al. 2016). Serotonin not only functions as neurotransmitters in animals but has also been reported to regulate many processes of growth and development in plants (Mukherjee et al. 2014; Park and Back 2012; Pelagio-Flores et al. 2011). Csaba and Katalin (1982) found that serotonin promoted the growth of barley roots and cotyledons. Apart from the beneficial effects on plant growth and development, serotonin also serves as a defensive agent against several abiotic stresses such as drought, salinity, heavy metals, and high temperature (Arnao and Hernández-Ruiz 2014; Nawaz et al. 2016; Shi et al. 2016). For instance, exogenous application of melatonin improves the drought resistance of rapeseed by increasing scavenging of ROS (Tian et al. 2019). Under the stress conditions, it can regulate the cellular ROS levels, thus preventing oxidative damage. Exogenous serotonin decreased the content of hydrogen peroxide (H2O2) and malondialdehyde (MDA) (Arnao and Hernández-Ruiz 2014; Nawaz et al. 2016; Shi et al. 2016). Moreover, exogenous serotonin alleviated seedling growth inhibition under salt stress and significantly increased fresh and dry weights of sunflower (Helianthus annuus L.) roots and shoots. It also promoted the root growth and hypocotyl elongation of sunflower seedlings under salt stress (Mukherjee et al. 2014).
Rapeseed (Brassica napus L.) is the largest oil crop in China and often experiences low-temperature stress in winter (Raza 2020b). Rapeseed yield parameters have been changed owing to the augmented exposure to several abiotic stresses, mainly cold stress, reducing biomass production and seed yield (Raza 2020b). Therefore, to understand the protective role of serotonin on rapeseed under cold stress, this experiment studied the effect of different concentrations of exogenous serotonin on Guanyouza303 variety under cold stress. The physiological and biochemical indexes and expression levels of cold tolerance-related genes were investigated in the seedlings pretreated with serotonin. That is a meaningful approach for understanding the mechanism of serotonin improving cold tolerance in rapeseed and the application of serotonin in the production.
Materials and methods
Plant materials and growth conditions
The rapeseed seeds of GuanYouza303 [(GuanYouza303 is a hybrid and relatively cold-tolerant rapeseed variety based on our previous experiment (Yan et al. 2018)] were obtained from the Oil Crops Research Institute, Chinese Academy of Agricultural Sciences, Wuhan, China. The seedling was grown followed by the procedure of Yan et al. (2018).
Chemicals
The list of all the chemicals and their sources used in this study (Table 1).
The treatments
About 10 seeds were sown in a 10 cm pot containing a mixture of vermiculite and nutrient soil (1:1). When the seedlings were grown to two leaves, they were thinned to 6–8 seedlings per pot to make sure the seedlings were similar. A few days later, the uniform seedlings with three leaves were selected to make sure that each pot had four seedlings. Before exposed to cold stress, seedlings were pretreated with 0.01, 0.02, 0.03, and 0.04 g L− 1 of serotonin solutions. Each pot with four seedlings was sprayed with 10 mL (2.5 mL plant− 1) of the above-mentioned serotonin solutions, and the seedlings of the control were sprayed with 10 mL of water. Each treatment set was repeated three times (4 pots per replication) and randomly arranged. One day after the pretreatment, all the seedlings were treated with cold stress following the method of Yan et al. (2019). Briefly, the seedlings were placed under 4 °C for 24 h for cold acclimation. Cold stress was imposed as a temperature of 2 °C for 1 h and then reduced gradually to 0 °C, and − 2 °C for 1 h, respectively. After each temperature treatment, green leaves were harvested and immediately frozen in liquid nitrogen and then stored at − 80 °C for the next experiments.
Physiological analysis
After the treatment of cold stress, seedlings were incubated at 4 °C for 24 h, and then the survival rate was measured. The concentrations of soluble sugar (SS), soluble protein (SP), and proline (Pro) were determined using commercial kits according to the manufacturer’s instructions (Solarbio, China). The content of MDA was determined by the thiobarbituric acid reaction (Hodges et al. 1999). The concentrations of superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD) were determined using commercial kits according to the manufacturer’s instructions (Solarbio, China). All the biochemical indices were performed using Multiskan FC (Thermo Fisher Scientific, USA) microplate reader.
The qRT-PCR analysis
To analyze the expression of stress-related marker genes, total RNA was extracted using a HiPure plant RNA Mini Kit (Megan, China) and treated with DNase I (Ambion, Austin, TX, USA) to purify the RNA samples. The cDNA was subsequently synthesized using RevertAid First-Strand Synthesis Kit (Thermo, USA) according to the manufacturer’s instructions.
The quantitative real time-polymerase chain reaction (qRT-PCR) reaction was performed employing ChamQTMSYBR® color qPCR Master Mix (High ROX Premixed) (Vazyme, Nanjing, China). The sequences of all primers were listed in Table 2, and actin of B. napus was used as the internal reference gene to analyze the marker genes’ transcript levels. The conditions for PCR were as follows: 30 s at 95 °C, followed by 40 cycles of 95 °C for 10 s, and 60 °C for 30 s. Three technical replicates of each sample were analyzed, and the data was processed using the 2−ΔΔCT method. All the performance of qRT-PCR was conducted using a StepOnePlus Real-Time PCR System (Applied Biosystems, USA).
Statistical data analysis
All the experiments were performed following a completely randomized design, and three biological replicates were included. All the data were subjected to analysis of variance (ANOVA), and the mean differences were compared by a Duncan’s multiple range test (DMRT) test using SPSS Statistics 25.0 software (SPSS Inc., Chicago, IL). Graphs were plotted using GraphPad Prism 7.0. Differences at P < 0.05 were considered significant.
Results
Effects of serotonin on the survival rate of rapeseed seedlings under cold stress
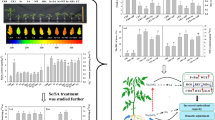
When the temperature dropped from 22 to − 2 °C, the freezing phenomenon occurred in rapeseed seedlings under stress. Upon recovery growth to room temperature, the stem and leaves began to lose water, leading to wilting. Necrosis spots gradually appeared, and some seedlings even died (Fig. 1a). The seedlings pretreated with serotonin showed a higher survival rate and less damage compared to the control seedlings. Therefore, the pretreatment with serotonin could significantly alleviate the wilting in leaves and increase rapeseed seedlings’ survival rate under cold stress. The survival rate of the seedlings pretreated with different serotonin concentrations was statistically analyzed (Fig. 1b). Compared with that of the control (12.5%), the seedlings’ survival rate with the pretreatment of 0.01, 0.02, and 0.03 g L− 1 of serotonin was significantly increased by 29.2%, 62.5%, and 75.5%, respectively. The results showed that with the increase of serotonin concentration in some range (0.01–0.03 g L− 1), the survival rate of the seedlings under the cold stress was increased at the beginning and then was decreased. In the present study, the optimal concentration of serotonin was 0.03 g L− 1, which had the highest survival rate (75.5%) under low-temperature.

The morphology and survival rate of rapeseed pretreated with serotonin under cold stress. a Morphology of the seedlings pretreated with different serotonin concentrations under cold stress. Red arrows indicate necrosis spot; b the survival rate of rapeseed pretreated with serotonin under cold stress. Error bars represent SD (n = 3). Different letters indicate significant differences (P < 0.05)
Effects of serotonin on physiological and biochemical indexes under cold stress
Effects of exogenous serotonin on the level of osmolytes under cold stress
The osmolytes’ content showed similar change trends in the seedlings pretreated with 0.03 g L− 1 serotonin and water (CK) when the temperature dropped from 22 to − 2 °C. The proline content was significantly increased under 0 and − 2 °C in the seedlings pretreated with 0.03 g L− 1 serotonin and water (CK) (Fig. 2a). Under cold stress, the soluble sugar content was gradually decreased when the temperature slowly dropped to − 2 °C (Fig. 2b). The content of soluble protein was reduced when exposed to 4 °C. Subsequently, it was slightly increased when the temperature was dropped to 2 °C, and it decreased as the temperature continued to decline (Fig. 2c). Exogenous serotonin increased the content of soluble sugar, soluble protein, and proline by 37–65%, 1–16%, and 7–38%, respectively, compared to that of the control. The content of soluble sugar was significantly increased by exogenous serotonin under 22 °C, while the content of soluble protein and proline was significantly increased with the serotonin treatment under cold stress.

Effect of serotonin (0.03 g L− 1) on the osmotic adjustment substances in rapeseed seedlings under cold stress. a The content of proline, b the content of soluble sugar, c the soluble protein content under 4, 2, 0, and − 2 °C compared to the control. Error bars represent SD. (n = 3) Different letters indicate significant differences (P < 0.05)
Effects of exogenous serotonin on the level of MDA content under cold stress
The level of MDA content is a negative symbol indicating the degree of membrane structural integrity (Janero 1990), and it is was investigated in rapeseed seedlings under cold stress. Under cold stress, the MDA content increased first and then decreased to rise again in both control and serotonin pretreated seedlings (Fig. 3). Mainly, 4 °C and 0 °C temperature were two main inflection points for MDA content under cold stress. Compared with the control, the MDA content in the seedlings pretreated with serotonin was decreased by 13–30% (Fig. 3). The pretreatment of serotonin significantly reduced MDA content under 22, 4, 2, 0, and − 2 °C.

Effect of serotonin (0.03 g L− 1) on malondialdehyde content changes in rapeseed seedlings under 4, 2, 0, and − 2 °C with control. Error bars represent SD (n = 3). Different letters indicate significant differences (P < 0.05)
Effects of exogenous serotonin on the antioxidative enzyme activities under cold stress
Under cold stress, ROS accumulation is increased in plants compared with that under normal growth conditions. Antioxidant enzymes, such as POD, CAT, and SOD, regulate the balance of ROS under stress conditions. In this study, the activities of CAT, POD, and SOD in leaves were detected. With the temperature dropped, CAT activity was decreased at first and then was increased (Fig. 4a). Simultaneously, POD and SOD activity was augmented at first and then was reduced in both the seedlings of the control and that pretreated with serotonin (Fig. 4b, c). The pretreatment of serotonin increased POD, CAT, and SOD activities by 10–31%, 4–42%, 9–27%, respectively. Under normal conditions, the activities of POD and CAT were significantly increased by exogenous serotonin. Under − 2 °C, CAT, POD, and SOD activities were increased dramatically by exogenous serotonin. The highest activity of POD and SOD were recorded in the seedlings pretreated with serotonin under 2 °C.

Effect of serotonin (0.03 g L− 1) on antioxidant enzymes’ activity in rapeseed seedlings under cold stress. a The activity of CAT, b the activity of POD, and c the activity of SOD under 4, 2, 0, and − 2 °C with control. Error bars represent SD (n = 3). Different letters indicate significant differences (P < 0.05)
Effects of exogenous serotonin on the expression of cold‐responsive genes under cold stress
In this study, exogenous serotonin improved the activity of SOD under cold stress. Therefore, the expression of the Cu/Zn-SOD genes was assayed by qRT-PCR. The results indicated that exogenous serotonin increased the expression of Cu/Zn-SOD-1-1 by 0.9–2.2 folds in the seedlings under cold stress (Fig. 5a). When the temperature dropped to − 2 °C, exogenous serotonin increased the expression of Cu/Zn-SOD-1-2 by 36.5 folds in the seedlings (Fig. 5b).

The effects of serotonin on the relative expression of SOD-related genes in rapeseed under cold stress. a Relative expression of Cu/Zn-SOD-1-1; b relative expression of Cu/Zn-SOD-1-2. Error bars represent SD (n = 3)
The expression of two cold-responsive marker genes (COR15 and COR6.6) was measured in this study. Exogenous serotonin increased the expression levels of COR6.6 by 218.2, 426.3, 312.9 and 231.2 folds compared to that in control under 4, 2, 0, and − 2 °C (Fig. 6a). Likewise, the expression level of COR15 in the seedlings pretreated with serotonin was 1137.4-1571.4 folds higher compared to that in control (Fig. 6b).

The effects of serotonin on the relative expression of COR genes in rapeseed under cold stress. a Relative expression of COR6.6; b relative expression of COR15. Error bars represent SD. (n = 3)
Previously, it has been reported that the promoter region of COR6.6 and COR15 genes contain CRT/DRE (C-repeat/Responsive Element), which is regulated by the CBF gene (Shengty, 2014). To determine whether CBF genes’ expressions were regulated by exogenous serotonin, the expression of CBF1, CBF2, and CBF4 were quantitatively analyzed in this study. After the cold treatment, exogenous serotonin increased the expression of CBF1-1, CBF1-2, CBF1-3, CBF1-4 by 195.5, 461.0, 87.0, 23.2 folds under 4 °C, 14.3, 63.0, 13.4, 17.8 folds under 2 °C, 82.2, 302.8, 2519.4, 732.4 folds under 0 °C, and 114.8, 281.0, 2019.3, 317.1 folds under − 2 °C, respectively (Fig. 7a, b, c, d). Similarly, under cold stress, exogenous serotonin increased the expression of the CBF2-1 gene by 203.9, 51.2, 488.8, 362.3 -folds under 4, 2, 0, and − 2 °C, and the CBF2-2 gene by 22.4, 14.2, 739.8, 2698.1 folds under 4, 2, 0, and − 2 °C (Fig. 7e, f). Moreover, it also increased the expression of CBF4-1 by 14.5, 1.9, 146.8, 188.5 folds under 4, 2, 0, and − 2 °C (Fig. 7g). Compared to the control, exogenous serotonin increased the expression of CBF4-2 by 538.7 fold under 0 °C, while the expression of CBF4-2 was increased 273.3 fold under − 2 °C, respectively (Fig. 7h). Thus, serotonin significantly increased the expression level of CBF1, CBF2, and CBF4 in rapeseed under cold stress.

The effects of serotonin on the relative expression of CBF genes in rapeseed under cold stress. a–d relative expression of CBF1 (CBF1-1, CBF1-2, CBF1-3, CBF1-4); e, f relative expression of CBF2 (CBF2-1, CBF2-2); g, h relative expression of CBF4 (CBF4-1, CBF4-2). Error bars represent SD (n = 3)
Discussion
In this study, rapeseed seedlings suffered from freezing stress when the temperature dropped to − 2 °C. Freezing stress has many deleterious effects on plants, such as disrupting the plasma membrane’s stability and flexibility, inducing intracellular water into the intercellular space, and forming ice crystals (Pearce 2001; McCully et al. 2004; Li et al. 2018a). Under freezing stress, plant cells not only lead to excessive dehydration of cells and tissue necrosis but also causes lytic damage to rapeseed cells in the process of ice crystal melting, which directly leads to wilting and death in plants (Becketal 1984). In the current study, when the seedlings recovered to grow at room temperature, the leaves gradually turned yellow and withered. This study showed that serotonin alleviates the severe wilting in stems and leaves caused by cold stress, and the concentration of 0.01 g L− 1 nearly avoided the wilting compared to the control. Results indicated that exogenous application of serotonin might play an important role in maintaining the stability of cell membranes and enhancing cold tolerance in rapeseed. Applied serotonin showed an increase in rapeseed survival rate with a 0.01–0.03 g L− 1 serotonin, and a significant decrease was observed with a low-temperature of 0.04 g L− 1 serotonin. Under low concentration, the effect was less and not prominent, whereas, under higher concentration, it would have an inhibitory effect, which was consistent with previous studies’ results (Bajwa et al. 2014; Tian et al. 2019). Therefore, it is essential to choose an appropriate concentration in the application of serotonin.
Additionally, cold stress also changed the protein structure and activity in the cells, decreased enzymes’ activity, and disturbed normal metabolic activities such as photosynthesis and respiration, affecting the normal growth and development in a plant and leading to wilting (Kodama et al. 1995). Altering the osmotic balance is proposed to be efficient to maintain the integrity and stability of the cell membrane for plant adaption under stress conditions. With the decreasing temperature, the content of SP increased at first and then decreased, which was consistent with the results of previous studies (Yan et al. 2019). Upon exposure to cold stress, the proline content increased and decreased with the decrease in the temperature. When the temperature dropped to − 2 °C, the proline content sharply increased, which may be due to different mechanisms in response to chilling and freezing stress (Wang et al. 2013). Besides, studies have also suggested that elevated proline content manifests plant cell damage (Jia et al. 2016). In this study, it was found that serotonin significantly increased the content of osmotic regulating substances (SS, SP, and proline), which was essential to alleviate the damage of cold stress in rapeseed. Tian et al. (2019) found that melatonin increases SS, SP, and proline content in drought-stressed rapeseed seedlings.
MDA is a common product of lipid peroxidation, and the content of MDA reveals the level of oxidative injury in plants. GuanYouza303, the variety used in this study, is a moderate cold-tolerant one (Yan et al. 2018). The MDA content was increased at 4 °C and then was decreased with the gradual decrease in the temperature. Notably, MDA content was raised under 0 °C and decreased again when the temperature continued to reduce. The changing trend of the MDA content in Guanyouza303 suggested that the level of MDA was closely related to the extent of cold stress and act as a negative symbol of cold tolerance (Zhang et al. 2016; Li et al. 2018c). In this study, the pretreatment of serotonin reduced MDA content by 6–15% under cold stress. Tian et al. (2019) found that serotonin reduced MDA content and alleviated the drought stress-induced damage in rapeseed seedlings. However, there were no differences in the MDA content’s changing trend between the seedlings with the pretreatment and the control.
Cold stress leads to an accumulation of ROS in plants, and the protective enzyme system plays an important role in ROS elimination. The activities of CAT, POD, and SOD in the protective system are commonly regarded as indicators of cold tolerance (Yan et al. 2018, 2019). In this study, CAT activity decreased gradually with the decrease in temperature and increased under − 2 °C. POD and SOD activities increased at first and then reduced; nonetheless, both reached a peak under 2 °C. The results indicated that POD and SOD activities were induced by chilling stress, while CAT activity was sensitive to freezing stress. Tian et al. (2019) found that CAT activity decreased gradually in drought stressed-rapeseed plants, while POD and SOD activities increased at first and then reduced under stress conditions compared to control plants. Yan et al. (2019) found that the CAT activity of C18 under freezing stress was higher than that under chilling stress, but that of SOD had no differences. In addition, this study also found that exogenous serotonin increased CAT activities, POD, and SOD under cold stress. Serotonin was considered free radical scavengers and antioxidants, removing ROS (Soumya 2018). The previous findings have shown that serotonin can alleviate ROS damage to plants under abiotic stress by increasing the activity of antioxidant enzymes (Soumya 2018; Tian et al. 2019). Interestingly, it has been found that serotonin can directly regulate the transcription level of antioxidant enzymes and improve tolerance of abiotic stress in the plant (Sharma et al. 2020; Mervat and Ahmed 2020).
Cu/Zn-SOD accounts for the largest proportion of SOD enzymes, and it plays a key role in scavenging ROS. Cu/Zn-SOD is closely related to cold, salt, and other abiotic stress tolerance in plants (Song et al. 2006). In the present study, qRT-PCR results showed that exogenous serotonin significantly increased Cu/Zn-SOD genes’ expression under cold stress, especially when the temperature was below 0 °C. Indicating that serotonin could increase the transcription level of SOD related genes, which explained the rise in the activity of SOD enzymes. Cen et al. (2020) also found that the metabolite of serotonin (melatonin) could increase the expression of Cu/Zn-SOD enzyme-related gene under salt stress (Cen et al. 2020). Several researches indicated that melatonin increased antioxidant enzymes’ activity and transcriptional level of antioxidant enzyme-encoding genes under drought, salt, and low-temperature stress (Yu et al. 2018; Sharma et al. 2020).
The signal pathway of an inducer of CBF expression (ICE), C-repeat binding factor(CBF), and cold-responsive (COR) genes plays a vital role under cold stress in plants (Yan et al. 2019). Therefore, the expression of some genes in this pathway was analyzed in this study. The results found that the pretreatment of serotonin significantly increased the expression levels of COR6.6, COR15, CBF1, CBF2, and CBF4 genes in rapeseed compared with CK under cold stress. It was indicated that serotonin was involved in the transcription network of rapeseed under cold stress by regulating COR and CBF genes’ expression. Shi et al. (2015) also found that the metabolite of serotonin (melatonin) can improve tolerance to salt damage, drought, and low-temperature stress by regulating the expression of AtCBF1, AtCBF2, AtCBF3, and COR15a in Arabidopsis (Shi et al. 2015). Moreover, the expression of genes related to the ABA-dependent signaling pathway under cold stress was also analyzed. The results showed that serotonin did not affect gene expression related to the ABA-dependent signaling pathway under cold stress. This indicated that serotonin did not increase cold tolerance through the ABA-dependent pathway in rapeseed.
Conclusions
In conclusion, exogenous serotonin improved cold tolerance by maintaining the osmotic potential balance in cells under cold stress by increasing osmolytes’ content (SS, SP, and proline). On the other hand, serotonin also advances ROS scavenging by increasing the activity of antioxidant enzymes (CAT, POD, and SOD) and the transcriptional level of stress-related genes in rapeseed. Besides, serotonin also participated in the signaling network of cold tolerance by promoting the expression of CBFs and CORs. Therefore, future research should focus on how serotonin regulated CBFs and CORs genes and how to apply serotonin in rapeseed production under adverse stress.
References
Agarwal PK, Agarwal P, Reddy M, Sopory SK (2006) Role of DREB transcription factors in abiotic and biotic stress tolerance in plants. Plant Cell Rep 25:1263–1274
Arnao MB, Hernández-Ruiz J (2014) Melatonin: plant growth regulator and/or biostimulator during stress? Trends Plant Sci 19:789–797
Bajwa VS, Shukla MR, Sherif SM, Murch SJ, Saxena PK (2014) Role of melatonin in alleviating cold stress in Arabidopsis thaliana. J Pineal Res 56:238–245
Cen HF, Wang TT, Liu HY, Tian DY, Zhang YW (2020) Melatonin application improves salt tolerance of alfalfa (Medicago sativa L.) by enhancing antioxidant capacity. Plants 9:220
Chinnusamy V, Zhu J, Zhu JK (2006) Gene regulation during cold acclimation in plants. Physiol Plant 126:52–61
Csaba G, Katalin Pál (1982) Effects of insulin, triiodothyronine, and serotonin on plant seed development. Protoplasma 110:20–22
Ding Y, Shi Y, Yang S (2019) Advances and challenges in uncovering cold tolerance regulatory mechanisms in plants. New Phytol 222:690–1704
Hasanuzzaman M, Bhuyan MB, Zulfiqar F, Raza A, Mohsin SM, Mahmud JA, Fujita M, Fotopoulos V (2020) Reactive oxygen species and antioxidant defense in plants under abiotic stress: revisiting the crucial role of a universal defense regulator. Antioxidants 9:681
Hodges DM, DeLong JM, Forney CF, Prange RK (1999) Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 207:604–611
Janero D (1990) Malondialdehyde and thiobarbituric acidreactivity as diagnostic indices of lipid peroxidation and peroxidative tissue injury. Free Radical Biol Med 9:515–540
Jia H, Hao L, Guo X, Liu S, Yan Y, Guo X (2016) A Raf-like MAPKKK gene, GhRaf19, negatively regulates tolerance to drought and salt and positively regulates resistance to cold stress by modulating reactive oxygen species in cotton. Plant Sci 252:267–281
Kaur H, Mukherjee S, Baluska F, Bhatla SC (2015) Regulatory roles of serotonin and melatonin in abiotic stress tolerance in plants. Plant Signal Behav 10:e1049788
Kodama H, Horiguchi GL, Nishiuchi T, Nishimura M, Iba K (1995) Fatty acid desaturation during chilling acclimation is one of the factors involved in conferring low-temperature tolerance to young tobacco leaves. Plant Physiol 107:1177–1185
Li D, Zhu Z, Sun DW (2018a) Effects of freezing on cell structure of fresh cellular food materials: A review. Trends Food Sci Technol 75:46–55
Li J, Zeng L, Cheng Y, Lu G, Fu G, Ma H, Li C (2018b) Exogenous melatonin alleviates damage from drought stress in Brassica napus L. (rapeseed) seedlings. Acta Physiol Plant 40:43
Li X, Wei JP, Scott ER, Liu JW, Guo S, Li Y, Zhang L, Han WY (2018c) Exogenous melatonin alleviates cold stress by promoting antioxidant defense and redox homeostasis in Camellia sinensis L. Molecules 23:165
Mervat SS, Ahmed B (2020) Alleviation of drought stress by melatonin foliar treatment on twoflax varieties under sandy soil. Physiol Mol Biol Plants 26:907–919
McCully ME, Canny MJ, Huang CX (2004) The management of extracellular ice by frosted, acclimated herbaceous petioles. Ann Bot 94:665–674
Miller G, Shulaev V, Mittler R (2008) Reactive oxygen signaling and abiotic stress. Physiol Plant 133:481–489
Mukherjee S, David A, Yadav S, Baluška F, Bhatla SC (2014) Salt stress-induced seedling growth inhibition coincides with differential distribution of serotonin and melatonin in sunflower seedling roots and cotyledons. Physiol Plant 152:714–728
Nawaz MA, Huang Y, Bie Z, Ahmed W, Reiter RJ, Niu M, Hameed S (2016) Melatonin: current status and future perspectives in plant science. Front Plant Sci 6:1230
Park S, Back K (2012) Melatonin promotes seminal root elongation and root growth in transgenic rice after germination. J Pineal Res 53:385–389
Pearce RS (2001) Plant freezing and damage. Ann Bot 87:417–424
Pelagio-Flores R, Ortíz-Castro R, Méndez-Bravo A, Macías-Rodríguez L, López-Bucio J (2011) Serotonin, a tryptophan-derived signal conserved in plants and animals, regulates root system architecture probably acting as a natural auxin inhibitor in Arabidopsis thaliana. Plant Cell Physiol 52:490–508
Radhakrishnan R, Lee IJ (2013) Ameliorative effects of spermine against osmotic stress through antioxidants and abscisic acid changes in soybean pods and seeds. Acta Physiol Plant 35:263–269
Raza A (2020) Eco-physiological and biochemical responses of rapeseed (Brassica napus L.) to abiotic Stresses: consequences and mitigation strategies. J Plant Growth Regul. https://doi.org/10.1007/s00344-020-10231-z
Raza A, Razzaq A, Mehmood SS, Zou X, Zhang X, Lv Y, Xu J (2019) Impact of climate change on crops adaptation and strategies to tackle its outcome: a review. Plants 8:34
Raza A, Ashraf F, Zou X, Zhang X, Tosif H (2020) Plant adaptation and tolerance to environmental stresses: mechanisms and perspectives. In: Plant ecophysiology and adaptation under climate change: mechanisms and perspectives I. Springer, Singapore, pp 117–145
Sharma A, Wang JF, Xu DB, Tao SC, Chong SL, Yan DL, Li Z, Yuan HW, Zheng BS (2020) Melatonin regulates the functional components of photosynthesis, antioxidant system, gene expression, and metabolic pathways to induce drought resistance in grafted Carya cathayensis plants. Sci Total Environ 713
Shi HT, Qian YQ, Tan DX, Russel JR, He CZ (2015) Melatonin induces the transcripts of CBF/DREB1s and their involvement in both abiotic and biotic stresses in Arabidopsis. J Pineal Res 59:334–342
Shi H, Chen K, Wei Y, He C (2016) Fundamental issues of melatonin-mediated stress signaling in plants. Front Plant Sci 7:1124
Strydhorst S, Hall L, Perrott L (2018) Plant growth regulators: What agronomists need to know. Crops Soils 51:22–26
Song FN, Yang CP, Liu XM, Li GB (2006) Effect of salt stress on activity of superoxide dismutase (SOD) in Ulmus Pumila L. J Forest Res 017:13–16
Soumya M (2018) Novel perspectives on the molecular crosstalk mechanisms of serotonin and melatonin in plants. Plant Physiol Biochem 132:33–45
Tian S, Li JQ, Zhang XK, Fu GP et al (2019) Effect of exogenous 5-hydroxytryptamine on rapessed (Brassica napus L.) seedling under drought stress. Chin J Oil Crops 41:192–198 (in Chinese)
Wang Y, Luo Z, Du R, Liu Y, Ying T, Mao L (2013) Effect of nitric oxide on antioxidative response and proline metabolism in banana during cold storage. J Agric Food Chem 61:8880–8887
Xin Z, Browse J (2000) Cold comfort farm: the acclimation of plants to freezing temperatures. Plant Cell Environ 23:893–902
Yan L, Cai J, Gao L, Huang B, Ma H, Liu Q, Dai X, Zhang X, Chen Y, Zou X (2018) Identification method and selection of cold tolerance in rapeseed (Brassica napus L.). Chinese J Oil Crop Sci 40:074–083 (in Chinese)
Yan L, Tariq S, Chen Y, Lü Y, Zhang X, Zou X (2019) Physiological and molecular responses to cold stress in rapeseed (Brassica napus L.). J Integr Agric 18:2742–2752
Yu Y, Lv Y, Shi YN, Li T, Chen YC, Zhao DK, Zhao ZW (2018) The role of phyto-melatonin and related metabolites in response to stress. Molecules 23:1887
Zhang N, Zhao B, Zhang HJ, Weeda S, Yang C, Yang ZC, Ren S, Guo YD (2013) Melatonin promotes water-stress tolerance, lateral root formation, and seed germination in cucumber (Cucumis sativus L.). J Pineal Res 54:15–23
Zhang Y, Yu H, Yang X, Li Q, Ling J, Wang H, Gu X, Huang S, Jiang W (2016) CsWRKY46, a WRKY transcription factor from cucumber, confers cold resistance in transgenic-plant by regulating a set of cold-stress responsive genes in an ABA-dependent manner. Plant Physiol Biochem 108:478–487
Funding
This work was supported by the National Key Research and Development Program (2017YFD0101700), Agricultural Science and Technology Innovation Program of CAAS, and the Hubei Agricultural Science and Technology Innovation Center.
Author information
Authors and Affiliations
Contributions
HH and LY performed the experiment and collected the data; HH, LY, and AR wrote and revised the manuscript; YZ helped in preparing the initial draft; AR, YL, and YC helped in the relevant literature; LZ and XD provided the reagents and materials; XZ designed and supervised the study.
Corresponding author
Ethics declarations
Conflict of interest
All authors declare that they have no conflict of interest.
Additional information
Communicated by: Luca Sebastiani.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
He, H., Lei, Y., Yi, Z. et al. Study on the mechanism of exogenous serotonin improving cold tolerance of rapeseed (Brassica napus L.) seedlings. Plant Growth Regul 94, 161–170 (2021). https://doi.org/10.1007/s10725-021-00700-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10725-021-00700-0