
Abstract
This study investigated the ability of exogenous melatonin (MT) to alleviate drought stress in Brassica napus L. (rapeseed) seedlings. Seedling traits under control condition, drought stress, and drought stress with exogenous MT were evaluated. The results indicated that 0.05 mmol/L exogenous MT had the greatest protective effect against simulated drought stress. Exogenous MT alleviated the seedling growth inhibition under drought stress and significantly increased the leaf area and fresh and dry weights of roots and shoots compared to stress conditions lacking MT. In addition, although the hydrogen peroxide (H2O2) content increased under drought stress, it was decreased by exogenous MT. Moreover, antioxidant enzyme activities were increased in response to drought stress, and the activities of catalase, ascorbate peroxidase, and peroxidase were significantly enhanced by exogenous MT. The results also showed that solute accumulation under stress was enhanced with exogenous MT through increases in the contents of soluble sugars and proteins. These results suggest that exogenous MT can alleviate the negative effects of drought stress and improve the growth of seedlings. The findings indicate that MT possesses antioxidative, osmotic activity-adjusting, and growth-inducing properties, thus making it beneficial for drought acclimatization.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Drought is a serious abiotic determinant of crop productivity worldwide (Basu et al. 2016). When plants are subjected to drought stress, the growth process is blocked, reducing seedling stem and root lengths and reducing shoot dry, root dry, and total dry weights (Zhang et al. 2014; Brunner et al. 2015). In addition, the relative water content and leaf area are decreased (Rymaszewski et al. 2017; Zong et al. 2017). Drought causes even more serious effects, such as wilting, local necrosis, or whole-plant death (Neumann 2008; Cominelli et al. 2013). Under drought, plants also develop multiple strategies to respond to the stress, including numerous alterations such as the up- or down-regulation of specific genes, a transient increase in abscisic acid (ABA) levels (Parvathi and Karaba 2015), accumulation of compatible solutes and protective enzymes (Hatzig et al. 2014), and enhanced levels of antioxidants and inhibition of energy-consuming pathways (Salekdeh et al. 2009; Huseynova 2012; Wang et al. 2016b). In these strategies, antioxidant capacity and osmotic adjustment are important indices of drought tolerance (Hatzig et al. 2014; Blum 2016).
Plant growth regulators are widely used to regulate growth and enhance resilience (Liu et al. 2011; Liu et al. 2016; Khan et al. 2017); therefore, exploring potential growth regulators and their mechanisms is of great importance to improving crop drought tolerance. Melatonin (MT, N-acetyl-5-methoxy tryptamine), an indole hormone widely present in plants and animals (Barratt et al. 1977; Dubbels et al. 1995; Reiter et al. 2011; Shi et al. 2016a), has many functions in plants, including the regulation of plant growth and development. Among other functions (Foyer et al. 1994), MT can promote lateral root growth (Wen et al. 2016), alter fluorescence (Byeon and Back 2014; Shi et al. 2016b) and delay leaf senescence (Byeon et al. 2012). In addition, MT can enhance plant tolerance to drought, salt stress, heavy metals, high temperature, and cold and other stresses (Arnao and Hernandez-Ruiz 2014; Nawaz et al. 2015; Shi et al. 2016a). Under water stress, MT can increase the germination rate of cucumber (Zhang et al. 2013), promote the growth of the root system, reduce damage to grape cuttings (Meng et al. 2014), and delay apple leaf senescence (Wang et al. 2013).
Rapeseed is one of the most important oil crops (Yin et al. 2009), and it has a large water requirement. Indeed, the water requirement during the entire growth period far exceeds than that of corn, wheat, and other crops (Rad and Zandi 2014). If water shortage occurs at the seedling stage, not only will it inhibit growth of leaves and the root system, but it will also affect flower bud differentiation and the flower angle number, ultimately leading to reduced production (Zhang et al. 2014; Bhardwaj et al. 2015). Although there are reports on drought tolerant effects of MT, there have been no studies on oil crops to date. Therefore, a study of the effect of MT on plant physiological and biochemical characteristics under drought condition has very important theoretical and practical significance for enhancing drought tolerance of rapeseed.
Materials and methods
Tested materials and reagents
Rapeseed variety ZS11 was provided by the Oil Crops Research Institute of the Chinese Academy of Agricultural Sciences, Wuhan, China. The MT and polyethylene glycol 6000 (PEG6000) used in the study were purchased from Shanghai Sinopharm Chemical Reagent Co., Ltd. Catalase (CAT), peroxidase (POD), proline (Pro), ascorbate peroxidase (APX), and other kits were obtained from Nanjing Jiancheng Bioengineering Institute, China.
Experimental methods
Preparation of the PEG6000 solution
Modified Hoagland’s nutrient solution (Dun et al. 2016; Murashige and Skoog 1962) was used as the basic medium to prepare a 4% PEG solution.
Preparation of MT solution
MT was added to 4% PEG solution to prepare 0, 25, 50, 75 and 100 µmol/L MT solutions.
Experimental treatment
Healthy and uniform seeds were carefully selected and sterilized in 3% NaOCl solution for 10 min. The seeds were then rinsed with distilled water, sown on fine gauze, and placed in a 24 °C culture room for 7 days under a 16/8 h light and dark cycle. Seven days later, the seeds were transferred to a hydroponic box containing Hoagland’s culture medium. At the two true leaf stage, seedlings with consistent growth were selected for the following treatments: control (CK1), 4% PEG (CK2, 40 g/L PEG), 4% PEG + 25 µmol/L MT (5.81 mg/L MT), 4% PEG + 50 µmol/L MT (11.61 mg/L MT), 4% PEG + 75 µmol/L MT (17.4 mg/L MT), and 4% PEG + 100 µmol/L MT (23.23 mg/L MT). Each treatment was repeated three times, and each replicate included eight seedlings.
Measurements
Assessment of physiological indicators
After 7 days of treatment, the root length, shoot length (height from cotyledon to growing point), and root and shoot fresh weights of each seedling were measured. Each seedling was then dried at 105 °C for 30 min and dried further at 80 °C to obtain a constant weight; the dry weight was then measured.
The third fresh leaf of each seedling was harvested and stored at − 80 °C for measurement of biochemical indicators, including concentrations of POD, APX, CAT, hydrogen peroxide (H2O2), proline, water-soluble proteins (WSP), and water-soluble sugars (WSG). All leaf biochemical indicators were assessed using Nanjing Jiancheng kits according to the manuals.
Data processing
The Microsoft Excel software was used to process the data. The SPSS 20.0 software was used to perform single factor analysis of variance at the level of 0.05, and the Origin 8.0 software was used to generate graphs.
Results
Effects of different concentrations of MT on physiological indicators
As shown in Table 1, compared with rapeseed seedlings (CK1) grown under normal growth conditions, various physiological indicators (CK2) decreased significantly under 4% PEG stress. The indicators, leaf area, root length, shoot and root fresh weights, and dry weight were most affected. The leaf area decreased to 74%, the shoot fresh weight decreased to 61%, and the root fresh weight decreased to 60.7%. The significant decrease of these indicators indicated tremendous damage to the seedlings. Compared with the various physiological indices of rapeseed seedlings under CK2 (4% PEG) drought stress, root application of different concentrations (25 µmol/L, 75 and 100 µmol/L) of MT significantly improved various physiological indices. Although the observed improvement was similar at these concentrations, 50 µmol/L MT showed the greatest effect; that is, the rates of improvement in leaf area (89%), shoot fresh weight (39%), root fresh weight (19%), shoot dry weight (20%), and root dry weight (16%) were all more obvious at 50 µmol/L compared with the other concentrations.
Effects of different concentrations of MT on biochemical indicators
Effect of exogenous MT on the activity of antioxidant enzymes
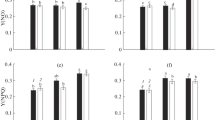
The results indicated that 4% PEG increased POD activity by 14% compared with the control CK1, and this increase in POD activity was more significant after the application of MT (Table 1 and Fig. 1). The highest POD activity was found in rapeseed leaves after the 50 µmol/L MT treatment, 64% greater than CK1. The POD activity of rapeseed leaves treated with 100 µmol/L MT was the second highest, at an increase of 50%, followed by leaves treated with 25 and 75 µmol/L MT, with increases of 47 and 29%, respectively, compared with CK1.

Effect of different concentrations of melatonin on POD activity under drought stress. Asterisk, double asterisk, and triple asterisk Significant at 0.05, 0.01, and 0.001 levels, respectively. Values are means of three biological replications, and bars indicate SEM
Compared with CK1, CAT activity decreased dramatically by 44% under 4% PEG stress (Table 2 and Fig. 2). However, CAT activity in leaves treated with 50 μmol/L MT was slightly higher than in treatments of several other concentrations. CAT activity in rapeseed leaves treated with 50 µmol/L MT decreased by only 24%, that in leaves treated with 25 µmol/L MT decreased by 31%, and that in leaves treated with 75 or 100 µmol/L MT decreased by 41%.

Effect of different concentrations of melatonin on CAT activity under drought stress. Asterisk, double asterisk, and triple asterisk Significant at 0.05, 0.01, and 0.001 levels, respectively. Values are means of three biological replications, and bars indicate SEM
Under PEG stress, activity of APX increased significantly in rapeseed seedlings, with an increase of 51% compared with CK, and the application of different concentrations of MT further enhanced APX activity (Table 2 and Fig. 3). The increase in APX was greatest when seedlings were treated with 50 µmol/L MT, with a 125% increase over that of CK1 followed by 25 µmol/L MT treatment, with an increase of 100%, and 75 µmol/L and 100 µmol/L MT, with respective increases of 90 and 77%.

Effect of different concentrations of melatonin on APX activity under drought stress. Asterisk, double asterisk, and triple asterisk significant at 0.05, 0.01, and 0.001 levels, respectively. Values are means of three biological replications, and bars indicate SEM
Effects of exogenous MT on the regulation of osmolyte concentrations
Compared with CK1, under PEG stress, WSG in rapeseed seedlings exhibited an upward trend, with an increase of 13% (Table 2 and Fig. 4). The increase in WSG was even greater after the application of different concentrations of MT, with the highest after treatment with 50 µmol/L MT, an increase of 42% compared with CK1, followed by 25, 75 and 100 µmol/L MT, which increased WSG by 31, 30, and 25%, respectively.

Effect of different concentrations of melatonin on WSG activity under drought stress. Asterisk, double asterisk, and triple asterisk significant at 0.05, 0.01, and 0.001 levels, respectively. Values are means of three biological replications, and bars indicate SEM
Compared with CK1, WSP in rapeseed showed a downward trend under 4% PEG stress, with a decrease of 10%, though WSP displayed an upward trend after application of different concentrations of MT (Table 2 and Fig. 5). WSP in rapeseed leaves treated with 50 μmol/L MT increased 53% compared with CK1, and the application of 25 μmol/L MT increased WSP by 48%, whereas that of 75 and 100 μmol/L MT increased WSP by 47 and 39%, respectively.

Effect of different concentrations of melatonin on WSP activity under drought stress. Asterisk, double asterisk, and triple asterisk significant at 0.05, 0.01, and 0.001 levels, respectively. Values are means of three biological replications, and bars indicate SEM
Under 4% PEG stress, the proline content increased by approximately 3.8-fold, and this increase was even greater after the application of different concentrations of MT (Table 2 and Fig. 6). The proline content in rapeseed leaves treated with 50 μmol/L MT was the highest, showing an increase of 12.1-fold compared with control CK1 and an increase of approximately 1.7-fold compared with 4% PEG stress. The proline content after the application of 75 µmol/L MT increased 10.1-fold compared with CK1, and that after the application of 25 and 100 μmol/L MT increased 8.7- and 8.1-fold, respectively, compared with CK. Treatment with 50 µmol/L MT had a more obvious effect with regard to enhancing the Pro content compared with the other concentrations.

Effect of different concentrations of melatonin on Pro activity under drought stress. Asterisk, double asterisk, and triple asterisk Significant at 0.05, 0.01, and 0.001 levels, respectively. Values are means of three biological replications, and bars indicate SEM
Effects of exogenous MT on H2O2 levels
H2O2 is an active oxygen molecule that causes oxidative stress. As shown in Table 2 and Fig. 7, the H2O2 concentration increased dramatically by 266% under 4% PEG stress compared that with under normal growth conditions. However, the H2O2 content decreased in all cases compared with the level under 4% PEG stress with the application of different concentrations of MT. The decrease in H2O2 content was the largest after the application of 50 μmol/L MT, a decrease of 58% compared with 4% PEG stress. After the application of 25, 75, and 100 μmol/L MT, the respective decreases in H2O2 content were 49, 47, and 46% compared with levels under 4% PEG stress.

Effect of different concentrations of melatonin on H2O2 activity under drought stress. Asterisk, double asterisk, and triple asterisk significant at 0.05, 0.01, and 0.001 levels, respectively. Values are means of three biological replications, and bars indicate SEM
Discussion
This study used PEG 6000 to simulate drought stress in rapeseed seedlings, and different concentrations of exogenous MT were applied to examine the changes in physiological and biochemical indicators. The leaf area, root and shoot fresh, and dry weights at the seedling stage are important indicators of growth and development that will directly affect the entire growth and development stages of rapeseed, and the results showed that drought stress severely inhibited the growth of rapeseed seedlings, as specifically demonstrated by these indicators. These indicators were significantly increased with exogenous MT. Although root length was not significantly increased or was even smaller than that under PEG stress without MT, the root fresh weight and dry weight were significantly increased, suggesting that exogenous application of MT can increase the number of roots, perhaps by promoting the growth of adventitious roots or lateral roots. A previous study (Zhang et al. 2014) showed that exogenous MT could effectively relieve the inhibition of cucumber seed germination caused by PEG stress, promote rooting, enhance root vitality, and improve the root/shoot ratio (Zhang et al. 2013). Even under normal condition, exogenous MT stimulates root growth in etiolated seedlings of Brassica juncea (Chen et al. 2009) and adventitious root regeneration in shoot tip explants of sweet cherry (Sarropoulou et al. 2012). In this study, after exogenous application of MT, leaf area and shoot fresh and dry weights were significantly increased. A previous study in soybean also showed that MT increased plant growth under drought stress (Wei et al. 2015). In addition, transcriptome analysis in Arabidopsis revealed that auxin-responsive genes were both up- and down-regulated in response to MT and that MT up-regulates the transcript levels of many defense-related factors, including stress receptors, kinases, and transcription factors (Weeda et al. 2014). In both our and previous studies, root application of MT could increase biomass accumulation in plants and relieve growth inhibition caused by drought stress.
When plants are exposed to drought, reactive oxygen species (ROS) such as H2O2, superoxide, singlet oxygen, and hydroxyl radical are produced naturally by a number of metabolic pathways (Becana et al. 2000). These free radicals disturb normal metabolism by oxidizing membrane lipids and denaturing proteins, photosynthetic pigments and nucleic acids (Lin and Ho 2000). To maintain the balance between ROS production and removal, plants have developed scavenging systems against ROS, including antioxidant enzymes such as APX, POD, and CAT, to eliminate excess ROS (Reddy et al. 2004; Ge et al. 2006). POD, CAT, and APX are important antioxidant enzymes in plants that scavenge hydroxyl radicals and H2O2 and thereby reduce damage to the cell membrane, mitigate membrane lipid peroxidation, and stabilize membrane permeability. In the present study, we measured POD, CAT, and APX activities and observed an upward trend for POD and APX in rapeseed seedlings after PEG stress, suggesting that these antioxidant enzymes respond to drought stress to reduce stress-induced membrane lipid peroxidation and stabilize membrane permeability. Related research showed that the activities of POD, APX, and other antioxidant enzymes all exhibit upward trends under drought stress (Salekdeh et al. 2009; Huseynova 2012; Wang et al. 2016a). However, in the present study, we observed downward trend for CAT under drought stress, and we speculate that this result may be related to the duration of the stress for rapeseed or the variety of rapeseed utilized. Zhang et al. reported that superoxide dismutase (SOD) activity was decreased and that CAT activity was first enhanced, but then declined as the duration of water stress increased (Salekdeh et al. 2009). After the root application of MT under drought stress, the activities of POD, CAT, and APX in rapeseed seedlings were all higher than those under drought stress, indicating that MT can enhance plant antioxidant enzyme activity and antioxidant capacity under drought stress. Studies by Rodriguez et al. showed that MT could directly or indirectly regulate the antioxidant enzyme system and increase enzyme activity in plants (Rodriguez et al. 2004). A study result by Zhao et al. showed that 0.1 mmol MT could improve the activities of SOD and CAT in Rhodiola rosea (Zhao et al. 2011), and the results of Wang et al. revealed significantly elevated levels of ascorbic acid and other antioxidants in apple seedlings treated with MT (Wang et al. 2012). Similar results were also reported in cucumber seedlings under high temperature stress (Xu et al. 2010). In this study, root application of MT significantly enhanced the activities of POD, CAT, and APX, which was helpful for scavenging ROS. The present study also measured H2O2 as representative of ROS, and the concentration of H2O2 also decreased in rapeseed seedlings treated exogenously with MT, an important endogenous free radical scavenger. Indeed, MT may directly scavenge H2O2 and help in maintaining intracellular H2O2 at steady-state levels. This homeostasis might be due to the inhibition of H2O2 accumulation and enhanced CAT and POD activities caused by endogenous MT supplementation. Related studies showed that the ROS levels in plant seedlings treated with MT under salt and drought stresses were significantly lower than those of plant seedlings without MT treatment (Li et al. 2012; Zhang et al. 2013).
In this study, proline and WSG levels increased under PEG stress, and these levels further increased in stressed rapeseed seedlings when MT was applied to roots. The WSG indicator was slightly decreased under PEG stress compared with under normal growth conditions, and this may be related to the duration of drought stress or the extent of stress suffered by the seedlings. A related study also reached a similar conclusion, whereby soluble proteins in rapeseed seedlings exhibited a downward trend when stressed using a high concentration of PEG (Zhang 2012). WSP in rapeseed seedlings treated with root-applied MT under drought stress showed a significant upward trend. Proline, WSG, and WSP are important osmotic regulators in plants. To maintain the water potential difference between the inside and outside of the cell for water absorption even at low external water potentials, the levels of osmotic regulatory substances in plants will increase under drought stress (Subbarao et al. 2000), thus maintaining the turgor required for cell growth and allowing growth during mild water shortage condition (Basu et al. 2016). Studies showed that the osmotic regulation capacity affects the degree of rapeseed response to drought (Wright et al. 1995) and its yield under drought stress (Wright et al. 1997). In addition, other studies also indicated that proline and WSG play a role in both osmotic protection and ROS scavenging, thus protecting the cell membrane (Trinchant et al. 2004; Verbruggen and Hermans 2008; Signorelli 2016).
Conclusion
The results of this study showed that exogenous MT has antioxidant activity, and can increase antioxidant enzyme activities to reduce the accumulation of free radicals, H2O2 and other substances that are harmful to the cell membrane. It can also increase osmolyte content to improve osmotic regulation capacity and enhance the response to water stress to alleviate damage caused by drought. Therefore, exogenous MT could promote root and leaf growth to increase rapeseed seedling biomass accumulation under drought stress.
Author contribution statement
Xiling Zou and Chunsheng Li conceived of the study and participated in its design. Jingjing Li carried out most of the work and drafted the manuscript. Liu Zeng was involved in designing and planning the work, and in interpreting the results. Yong Cheng, Guangyuan Lu, and Guiping Fu prepared the seeds and helped with the data analysis. Qinghai Ma and Qingyun Liu participated the assessment of physiological indicators. Yongguo Zhao helped with the manuscript writing. All authors have read and approved the final manuscript.
References
Arnao MB, Hernandez-Ruiz J (2014) Melatonin: plant growth regulator and/or biostimulator during stress? Trends Plant Sci 19:789–797
Barratt GF, Nadakavukaren MJ, Frehn JL (1977) Effect of melatonin implants on gonadal weights and pineal gland fine structure of the golden hamster. Tissue Cell 9:335–345
Basu S, Ramegowda V, Kumar A, Pereira A (2016) Plant adaptation to drought stress. F1000Res 5:1–10
Becana M, Dalton DA, Moran JF, Iturbe-Ormaetxe I, Matamoros MA, Rubio MC (2000) Reactive oxygen species and antioxidants in legume nodules. Physiol Plant 109:372–381
Bhardwaj AR, Joshi G, Kukreja B, Malik V, Arora P, Pandey R, Shukla RN, Bankar KG, Katiyaragarwal S, Goel S (2015) Global insights into high temperature and drought stress regulated genes by RNA-Seq in economically important oilseed crop Brassica juncea. BMC Plant Biol 15:9
Blum A (2016) Osmotic adjustment is a prime drought stress adaptive engine in support of plant production. Plant Cell Environ 40:4–10
Brunner I, Herzog C, Dawes MA, Brunner I, Herzog C, Dawes MA, Arend M, Sperisen C (2015) How tree roots respond to drought. Front Plant Sci 6:547
Byeon Y, Back K (2014) An increase in melatonin in transgenic rice causes pleiotropic phenotypes, including enhanced seedling growth, delayed flowering, and low grain yield. J Pineal Res 56:408–414
Byeon Y, Park S, Kim YS, Park DH, Lee S, Back K (2012) Light-regulated melatonin biosynthesis in rice during the senescence process in detached leaves. J Pineal Res 53:107–111
Chen Q, Qi WB, Reiter RJ, Wei W, Wang BM (2009) Exogenously applied melatonin stimulates root growth and raises endogenous indoleacetic acid in roots of etiolated seedlings of Brassica juncea. J Plant Physiol 166:324–328
Cominelli E, Conti L, Tonelli C, Galbiati M (2013) Challenges and perspectives to improve crop drought and salinity tolerance. N Biotechnol 30:355–361
Dubbels R, Reiter RJ, Klenke E, Goebel A, Schnakenberg E, Ehlers C, Schiwara HW, Schloot W (1995) Melatonin in edible plants identified by radioimmunoassay and by high performance liquid chromatography-mass spectrometry. J Pineal Res 18:28–31
Dun X, Tao Z, Wang J, Wang X, Liu G, Wang H (2016) Comparative transcriptome analysis of primary roots of Brassica napus seedlings with extremely different primary root lengths using RNA sequencing. Front Plant Sci 7:1238
Foyer CH, Descouvieres P, Kunert KJ (1994) Protection against oxygen radicals: an important mechanism studied in transgenic plants. Plant Cell Environ 17:507–523
Ge TD, Sui FG, Bai LP, Lu Y, Zhou G (2006) Effects of water stress on the protective enzyme activities and lipid peroxidation in roots and leaves of summer maize. Agric Sci China 5:291–298
Hatzig S, Zaharia LI, Abrams S, Hohmann M, Legoahec L, Bouchereau A, Nesi N, Snowdon RJ (2014) Early osmotic adjustment responses in drought-resistant and drought-sensitive oilseed rape. J Integr Plant Biol 56:797–809
Huseynova IM (2012) Photosynthetic characteristics and enzymatic antioxidant capacity of leaves from wheat cultivars exposed to drought. Biochim Biophys Acta 1817:1516–1523
Khan A, Anwar Y, Hasan M, Iqbal A, Ali M, Alharby H, Hakeem K, Hasanuzzaman M (2017) Attenuation of drought stress in brassica seedlings with exogenous application of Ca2 + and H2O2. Plants 6:1–13
Li C, Wang P, Wei Z, Liang D, Liu CH, Yin LH, Jia DF, Fu MY, Ma FW (2012) The mitigation effects of exogenous melatonin on salinity-induced stress in Malus hupehensis. J Pineal Res 53:298–306
Lin MY, Ho LT (2000) Melatonin suppresses iron-induced neurodegeneration in rat brain. Free Radical Biol Med 28:904
Liu D, Pei ZF, Naeem MS, Ming DF, Liu HB, Khan F, Zhou WJ (2011) 5-aminolevulinic acid activates antioxidative defence system and seedling growth in Brassica napus L. under water-deficit stress. J Agron Crop Sci 197:284–295
Liu D, Hu LY, Ali B, Yang AG, Wan GL, Xu L, Zhou WJ (2016) Influence of 5-aminolevulinic acid on photosynthetically related parameters and gene expression in Brassica napus L. under drought stress. Soil Sci Plant Nutr 62:254–262
Meng JF, Xu TF, Wang ZZ, Fang Y, Xi Z, Zhang Z (2014) The ameliorative effects of exogenous melatonin on grape cuttings under water-deficient stress: antioxidant metabolites, leaf anatomy, and chloroplast morphology. J Pineal Res 57:200–212
Murashige T, Skoog F (1962) A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol Plant 15:473–497
Nawaz MA, Huang Y, Bie Z, Waqar A, Reiter RJ, Niu ML, Saba H (2015) Melatonin: current status and future perspectives in plant science. Front Plant Sci 6:1230
Neumann PM (2008) Coping mechanisms for crop plants in drought-prone environments. Ann Bot 101:901–907
Parvathi MS, Karaba NN (2015) Emerging tools, concepts and ideas to track the modulator genes underlying plant drought adaptive traits: an overview. Plant Signal Behav 11:1
Rad AHS, Zandi P (2014) Agronomic evaluation of rapeseed varieties (Brassica napus L.) in response to late-season water deficit stress. Agric Conspectus Scientificus 79:157–165
Reddy AR, Chaitanya KV, Jutur PP, Sumithra K (2004) Differential antioxidative responses to water stress among five mulberry (Morus alba L.) cultivars. Environ Exp Bot 52:33–42
Reiter RJ, Coto-Montes A, Boga JA, Fuentes-Broto L, Rosales-Corral S, Tan DX (2011) Melatonin: new applications in clinical and veterinary medicine, plant physiology and industry. Neuro Endocrinol Lett 32:575–587
Rodriguez C, Mayo JC, Sainz RM, Antolín I, Herrera F, Martín V, Reiter RJ (2004) Regulation of antioxidant enzymes: a significant role for melatonin. J Pineal Res 36:1–9
Rymaszewski W, Vile D, Bédiée A, Dauzat, M, Luchaire N, Kamrowska D, Granier C, Hennig J (2017) Stress-response gene expression reflects morpho-physiological responses to water deficit. Plant Physiol
Salekdeh GH, Reynolds M, Bennett J, Boyer J (2009) Conceptual framework for drought phenotyping during molecular breeding. Trends Plant Sci 14:488–496
Sarropoulou VN, Therios IN, Dimassi-Theriou KN (2012) Melatonin promotes adventitious root regeneration in in vitro shoot tip explants of the commercial sweet cherry rootstocks CAB-6P (Prunus cerasus L.), Gisela 6 (P. cerasus × P. canescens), and MxM 60 (P. avium × P. mahaleb). J Pineal Res 52:38–46
Shi H, Chen K, Wei Y, He C (2016a) Fundamental issues of melatonin-mediated stress signaling in plants. Front Plant Sci 7:1124
Shi H, Wei Y, Wang Q, Reiter RJ, He C (2016b) Melatonin mediates the stabilization of DELLA proteins to repress the floral transition in Arabidopsis. J Pineal Res 60:373–379
Signorelli S (2016) The fermentation analogy: a point of view for understanding the intriguing role of proline accumulation in stressed plants. Front Plant Sci 7:1339
Subbarao GV, Chauhan YS, Johansen C (2000) Patterns of osmotic adjustment in pigeonpea—its importance as a mechanism of drought resistance. Eur J Agron 12:239–249
Trinchant JC, Boscari A, Spennato G, Van DSG, Le RD (2004) Proline betaine accumulation and metabolism in alfalfa plants under sodium chloride stress. Exploring its compartmentalization in nodules. Plant Physiol 135:1583–1594
Verbruggen N, Hermans C (2008) Proline accumulation in plants: a review. Amino Acids 35:753–759
Wang P, Yin L, Liang D, Li C, Ma F, Yue Z (2012) Delayed senescence of apple leaves by exogenous melatonin treatment: toward regulating the ascorbate-glutathione cycle. J Pineal Res 53:11–20
Wang P, Sun X, Li C, Wei Z, Liang D, Ma F (2013) Long-term exogenous application of melatonin delays drought-induced leaf senescence in apple. J Pineal Res 54:292–302
Wang H, Yang L, Li Y, Hou J, Huang J, Liang W (2016a) Involvement of ABA- and H2O2-dependent cytosolic glucose-6-phosphate dehydrogenase in maintaining redox homeostasis in soybean roots under drought stress. Plant Physiol Biochem 107:126–136
Wang R, Gao M, Ji S, Wang S, Meng Y, Zhou Z (2016b) Carbon allocation, osmotic adjustment, antioxidant capacity and growth in cotton under long-term soil drought during flowering and boll-forming period. Plant Physiol Biochem 107:137–146
Weeda S, Zhang N, Zhao X, Ndip G, Guo YD, Buck GA, Fu CG, Ren S (2014) Arabidopsis transcriptome analysis reveals key roles of melatonin in plant defense systems. PLoS One 9:e93462
Wei W, Li QT, Chu YN, Reiter RJ, Yu XM, Zhu DH, Zhang WK, Ma B, Lin Q, Zhang JS (2015) Melatonin enhances plant growth and abiotic stress tolerance in soybean plants. J Exp Bot 66:695–707
Wen D, Gong B, Sun SS, Liu SQ, Wang XF, Wei M, Yang FJ, Li Y, Shi QH (2016) Promoting roles of melatonin in adventitious root development of Solanum lycopersicum L. by regulating auxin and nitric oxide signaling. Front Plant Sci 7:718
Wright PR, Morgan JM, Jessop RS, Cass A (1995) Comparative adaptation of canola (brassica napus) and indian mustard (b. juncea) to soil water deficits: yield and yield components. Field Crops Research 42:1–13
Wright PR, Morgan JM, Jessop RS (1997) Turgor maintenance by osmoregulation in Brassica napus and B. juncea under field conditions. Ann Bot 80:313–319
Xu XD, Sun Y, Sun B, Zhang J, Guo XQ (2010) Effects of exogenous melatonin on active oxygen metabolism of cucumber seedlings under high temperature stress. Yingyong Shengtai Xuebao (Chin J Appl Ecol) 21:1295–1300
Yin Y, Wang HZ, Liu X (2009) Analysis and strategy for 2009 rapeseed industry development in China. Chin J Oil Crop Sci 2:028
Zhang T (2012) Effects of PEG6000-simulated drought stress on physiological and biochemical characteristics of rape seedlings. J Anhui Agric Sci 20:003
Zhang N, Zhao B, Zhang HJ, Weeda S, Yang C, Yang ZC, Ren S, Guo YD (2013) Melatonin promotes water-stress tolerance, lateral root formation, and seed germination in cucumber (Cucumis sativus L.). J Pineal Res 54:15–23
Zhang X, Lu G, Long W, Zou X, Li F, Nishio T (2014) Recent progress in drought and salt tolerance studies in Brassica crops. Breed Sci 64:60–73
Zhao Y, Qi LW, Wang WM, Saxena PK, Liu C (2011) Melatonin improves the survival of cryopreserved callus of Rhodiola crenulata. J Pineal Res 50:83–88
Zong H, Liu S, Xing R, Chen X, Li P (2017) Protective effect of chitosan on photosynthesis and antioxidative defense system in edible rape (Brassica rapa L.) in the presence of cadmium. Ecotoxicol Environ Saf 138:271
Acknowledgements
This study was supported by the National Science Foundation of China (31371663), Agricultural Science and Technology Innovation Program of CAAS, Canola Key Industrial Innovation Team of Xiaogan, and the Hubei Agricultural Science and Technology Innovation Center.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Communicated by W Zhou.
Rights and permissions
About this article

Cite this article
Li, J., Zeng, L., Cheng, Y. et al. Exogenous melatonin alleviates damage from drought stress in Brassica napus L. (rapeseed) seedlings. Acta Physiol Plant 40, 43 (2018). https://doi.org/10.1007/s11738-017-2601-8
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11738-017-2601-8