
Abstract
We made a first descriptive study of weedy sunflowers infesting sunflower crop fields in one Spanish and three French regions. Overall, weedy sunflowers affected around 15% of sunflower fields. Infested fields were most often dispersed over the study areas without evident geographical clustering. In France, five weedy populations were surveyed more intensively. They were composed of a large diversity of morphotypes showing a combination of typical wild and domesticated traits in proportions that differed between populations. Yield losses reached 50% in heavily infested patches. Our results suggest that weedy sunflowers may have arisen through the hybridization of cultivated and wild sunflower, potentially during the seed production process. Such crop-wild hybrids would have been introduced recurrently into fields through the seed lots, where they evolved to locally invasive weedy populations.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In the course of crop evolution, the development of agriculture created a new environment, the agro-ecosystem, constituted by agricultural fields, cultivated or not, and their margins. Some crop-related plants have adapted to this environment in an undesired way, leading to agricultural weeds (Arnold 2004). These weeds can be defined as plants that have the capacity to compete with the crop and to lead to more or less severe yield losses (Basu et al. 2004). Crop-related weeds have evolved through different kinds of process (Londo and Schaal 2007): the adaptation of wild populations to the agro-ecosystem (e.g. wild rice taxa in Asia, Ellstrand 2003), crop-wild hybridization (Arnold 2004; Boudry et al. 1993), reversion of the cultivated plants to wild habits. Among these processes, crop-wild and crop-weed hybridization is recognized as being very important in generating more noxious weeds (Arnold 2004; Campbell et al. 2006). The study of crop-related weeds can help to prevent further weed evolution and to design appropriate management methods. Crop-related weeds may also constitute interesting genetic resources. Moreover, they may be the focus of basic research as examples of the recent evolution and adaptation of plants species to new environments (Kane and Rieseberg 2008).
Sunflower (Helianthus annuus L.) was domesticated in North America at least 4,000 years ago (Harter et al. 2004). It was introduced into Europe, probably through Spain, in the sixteenth century, first as an ornamental plant (Putt 1997). Breeding of oil-producing varieties started in Russia in the nineteenth century, and from there, sunflower expanded as an oilseed crop in other European countries and all over the world (Putt 1997). Main traits distinguishing wild Helianthus annuus L. from present cultivated varieties are: strong branching, presence of anthocyanin pigmentation in various organs, reduced size of head and achenes, seed dormancy, seed shattering, and sporophytic self-incompatibility (Burke et al. 2002; Gandhi et al. 2005).
Although not a native of Europe, H. annuus is sometimes observed without having been sown by farmers. First, as for every crop, seeds lost at harvest can germinate and give rise to volunteer plants. Such sunflower plants naturally grow in cultivated areas and display phenotypic traits similar to those of the varieties, with the exception of branching for some of them (Reagon and Snow 2006). Second, weedy plants, showing typical wild phenotypic traits (e.g. Fig. 1), have been described for decades in Italy (Vischi et al. 2006), Spain (Faure et al. 2002) and Central Europe (Holec et al. 2005). They were officially confirmed in France in 2004 (Muller et al. 2006) although some farmers have been confronted with them since as early as 1990. In France and Spain, they mainly occur within sunflower fields (Fig. 2) and are thus potentially problematic for sunflower cultivation. Few quantitative data are available on these European weedy populations.

Some distinctive traits of weedy sunflowers. a Strongly branched plant without apical dominance; b disk pigmentation; c stem pigmentation; d seed shattering. Weedy sunflowers generally combine some of these traits with cultivated-like traits

Patch of weedy sunflowers within a sunflower cultivated field
The objectives of the present studies were to
-
Describe the distribution of weedy sunflower populations in four geographical areas: Andalusia, and three French regions
-
Assess the morphological diversity within and among given weedy populations and their impact on sunflower cultivation.
The results could constitute a baseline for studying the process of weed evolution as well as for identifying useful genetic resources.
Materials and methods
Survey procedure
In every region we followed predefined roads and observed all sunflower fields along these roads. For each field, we scored the infestation level by weedy sunflowers on the following scale: 0 (no weedy sunflower), 1 (less than 10 plants), 2 (a few patches) and 3 (heavily infested). We were careful not to confuse weedy sunflowers with other kinds of branched sunflowers such as volunteers (i.e. F2 plants issued from seeds lost from a previous harvest) or branched plants of the variety itself (a phenomenon that sometimes happens due to climatic conditions). The criteria used to adequately identify infestation by weedy sunflowers were the occurrence of plants growing between the rows of the crop and showing one of the following wild traits: anthocyanins in stem and/or disk, strong branching, small size of the disks and reduced apical dominance. Not all weedy plants will show these traits, but at least some are present in some of the plants.
In Spain, from 20th to 30th July 2005, we travelled through 650 km of roads around Cordoba (225 km) and in the agricultural area between Cordoba and Seville (425 km, Campiña, Fig. 3). In this region, weedy sunflower also grow outside sunflower fields (Fig. 4); therefore, we also recorded weedy sunflower populations observed with no direct connection to a sunflower field (i.e. excluding infested field margins), when these consisted of more than about 20 plants. These populations are referred to as “escaped weedy populations” in the following.

Map showing the surveyed area of Andalusia (shadowed zone) and the location of the main sites observed in 2005. ◊, Heavily infested sunflower fields (levels 2 and 3); ▲, Main escaped weedy sunflower populations (i.e. observed with no direct connection with an infested field at the time of our visit. See text for details). Some points may correspond to two sites close by. Infested fields are often associated with weedy sunflowers located in ditches or roadsides

Weedy sunflowers growing outside cultivated fields (Spain)
In France, we surveyed three regions (Fig. 5a). In the Lauragais region, we followed the same roads over an area of approximately 1,500 km2 during summers 2005, 2006 and 2007. Only the results of 2007 (23–24th July) will be shown for comparisons with the other French regions. In summer 2005, the survey was included in a wider expedition allowing us to locate heavily infested fields all over the region (Fig. 5b). In the “Gers-Lot et Garonne” region, the survey was conducted from the 17th to the 30th July 2007 and covered an area of approximately 5,000 km2; in the “Poitou-Charentes” region, observations were made on the 25th July and 1st August 2007 over an area of around 1,500 km2. The percentage of the area cultivated in sunflower varies between regions and year but is consistently lower in the “Gers-Lot et Garonne” region. The standard intercropping duration is 2–3 years. In France, latitude and longitude of all fields were measured using global positioning system. In Spain, only the coordinates of infested fields were measured.

a Map of France showing the three surveyed regions (shadowed zones). b Maps of region Lauragais showing the surveyed roads (interrupted lines), the heavily infested fields located during the first expedition in 2005 (◊), and the weedy populations surveyed in 2006 (▲). These weedy populations are called by the name of the nearest town
Morphological diversity and agronomic impact
In summer 2006, we surveyed five sunflower fields in the Lauragais region. These fields were selected at the beginning of July, as they showed a heavy infestation with weedy sunflower (level 3). In four of these fields, an experiment was conducted to estimate the impact of weeds on yield. Weeds were manually destroyed on six rows of cultivated sunflower (weeded—30 m2) and the density of weeds in an adjacent area of the same size was estimated (infested). At the end of the season, crop heads were manually harvested on the weeded and the infested plots, on 24 m2 only to avoid border effect. The seed production, seed weight and oil content of the crop were determined.
On 18 and 25th July, during the flowering period of the weeds, 30 branched weeds were randomly chosen on each of the five fields. The following traits were scored: male-sterility and anthocyanin pigmentation of petiole, disk and stem. One head per plant was bagged before the opening of the first flower. At the end of August, bagged heads were collected, and from each of the surveyed plants, at least one open-pollinated head was collected. The number of achenes produced by the bagged head was determined in the laboratory. A plant was scored as self-incompatible if less than five achenes had been produced, and as self-compatible in the other case.
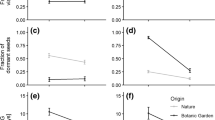
In spring 2006, six achenes (single-seeded fruits, which will be referred to as seeds in the following) from open-pollinated heads per maternal plant, were placed on filter paper and soaked in tap water. After 3 days at 25°C, the number of germinated seeds was counted. If no seed had germinated in a maternal family, we removed the seed coat and allowed two more days to germinate. We scored the need to remove the seed coat to obtain germination of at least one seed per maternal plant. We considered this trait as indicative of pericarp-imposed seed dormancy.
All together, six traits had been scored. For male-sterility, we computed the frequency of male-sterile plants per population. For each of the other traits and each population, the frequency of the wild type (i.e. anthocyanins in each of three organs, self-incompatibility and seed dormancy) was computed. The significance of differences in frequency across the set of populations was tested with a χ2 test on contingency tables. The correlation between traits within each population was investigated with exact test on contingency table. All tests were performed with SAS software (SAS Institute, Cary, NC, USA).
Results
Field surveys in Spain and France
On Fig. 6, we compared the frequencies of the different levels of infestation in Spain, and in the three French regions. The frequency of infested sunflower fields varied between 27% in Gers-Lot et Garonne and 13% in Andalusia. In France, the highest infestation rates (level 3) were observed in the Lauragais region: in these heavily infested fields, the density of weeds can reach 15 plants/m2 over some patches. The frequencies observed in Andalusia differed greatly between the two sub regions surveyed, Cordoba and Campiña as shown in Table 1. In the Cordoba region, escaped weedy populations were observed more frequently compared to the Campiña region, although the distance covered was shorter.

Rates of infestation estimated in one Spanish region in 2005 and in three French regions in 2007 (in percentages). The total number of fields observed was respectively 300, 196,195, and 100
In none of the region was any geographical clustering of infested fields observed, and heavily infested fields were dispersed all over the areas surveyed (Figs. 3, 5). In the “Gers-Lot et Garonne” region, where the area covered was much larger, infested fields were however concentrated on an area of approximately 800 km2 around Lectoure (data not shown). On a smaller scale, the absence of clustering was more striking: indeed, many heavily infested fields were located beside completely uninfested fields. In some cases however, the field adjoining a heavily infested fields, was also sparsely infested (data not shown).
Impact on yield
High densities of weedy sunflowers greatly decreased crop yield (Table 2): Losses can reach more than 50%. These losses are due to a reduction in achene number and achene weight. The seed oil content was unaffected (data not shown). Visually, weedy sunflowers greatly overtopped the crop (Fig. 2).
Morphological traits
The weedy forms were morphologically clearly different from the volunteers arising from the segregation of hybrid-F1 varieties. Our formal observations confirmed that they exhibit traits typical of wild forms of H. annuus: pigmentation of disk, stems and petioles, self-incompatibility, and seed dormancy (Fig. 7). A trait more typical of a cultivated variety, although sometimes observed in wild populations, male-sterility, was also observed at a varying rate in the weedy populations. For five out of six traits, the frequency varied significantly between populations. These frequency variations were correlated between traits, namely Baziège was the population with the lowest frequency of wild-like traits for all traits, whereas Villefranche was the population with most wild phenotypes. This tendency was also observed for less quantifiable traits such as seed shattering, or amount of branching.

Frequency of phenotypic traits in the different weedy populations. Significance of frequency differences between populations: * P < 0.05; ** P < 0.01; ns not significant
Within populations, significant correlations between traits were only observed between the scoring of anthocyanins in different organs (stem, petioles and disk). No other correlation was detected, confirming the visual observation that all kinds of combination of traits were present in the weeds. Weedy populations are characterized by a high morphological diversity with plants combining in different proportions domesticated and wild traits, from typical F2 plants to typical wild-like phenotypes.
The flowering periods of cultivated and weedy sunflowers overlap, the first weeds starting flowering during the flowering of the crop. But the flowering period of the weeds extends almost until harvest time.
Discussion
Our study demonstrated formally the occurrence of weedy forms of sunflower in France and Spain. These plants show a combination of typically wild and domesticated traits, they grow in the sunflower fields, and can lead to a yield loss of more than 50% when they reach a high density. They infested about 15% of fields in the surveyed areas. Up to 4% of fields were heavily infested in the Lauragais region, which has been the focus of the most detailed studies. In such fields, harvest is difficult and in some cases, farmers have given up sunflower cultivation. Our data provide only a rough estimation of weedy sunflower infestation: uncertainties result from varying size of the fields, and varying weed density within infested fields. More accurate observations on a reduced territory, as well as interviews with farmers, would allow us to describe more precisely the distribution and density of weedy sunflowers and to infer their economic impact at a regional scale. Very occasionally, weedy sunflowers have also been observed in summer crops such as sorghum. Moreover, in Spain, we saw many “escaped” weedy sunflower populations, developing outside cultivated fields, e.g. in ditches or roadsides.
The situations of the naturalized sunflower outside its area of origin are quite diverse and show that this species has adapted to a variety of environments (Cantamutto et al. 2008). In Italy, it infests sunflower and other crops such as maize, tobacco, alfalfa, but also spontaneously grows along roads and in ditches (Vischi et al. 2006). In the Czech Republic, it is mainly observed in non arable land, but it has recently been pointed out to be a weed in sunflower fields (Holec et al. 2005). In Argentina, large populations of Helianthus annuus L. and Helianthus petiolaris Nutt. occur on wide areas in roadsides, ditches and on disturbed soils, but relatively rarely in sunflower fields (Ureta et al. 2008). Even if no quantitative data are available, these weeds cause problems in all these cases, among which yield impact, concern on coexistence with GMO crops and risks of contamination of seed production stand out.
In Spain and France, the occurrence of these weedy sunflowers within sunflower fields, and the frequency of typical wild traits suggest that they may have originated via seed lot contamination with wild seeds or crop-wild hybrids. This hypothesis has already been mentioned for Spain and Italy (Vischi et al. 2006). This origin has been demonstrated for other species, for instance weed beets (Boudry et al. 1993). An alternative hypothesis could be the spontaneous evolution of wild-type traits in volunteers. However this evolution has rarely been evidenced (although see Londo and Schaal 2007) and does not appear to be very probable considering the diversity of phenotypes observed and the frequency of some traits such as anthocyanin pigmentation within weedy populations. Weedy sunflowers may have evolved many times independently even within a given region as suggested by the absence of any geographical clustering of heavily infested fields (Fig. 5b). Pigmented and branched ornamental annual sunflowers which are sometimes cultivated in gardens may also have contributed to this evolution. Only the ongoing genetic analysis will allow the confirmation and the clarification of this scenario. By contrast, in Argentina where it is mainly observed in unmanaged areas, weedy H. annuus has supposedly evolved from wild sunflower cultivated as forage before 1949 and established after the end of cultivation (Cantamutto et al. 2008).
Within the five weedy populations surveyed in detail, the low association detected between wild-type traits and the high diversity of phenotypes combining wild and domesticated traits suggests that crop-weed hybridization is frequent and recurrent. This is highly probable considering the occurrence of both forms in the same fields and the overlapping of flowering. The rate of spontaneous crop-weed and weed-crop hybridization has been investigated in different conditions for weedy sunflower in Argentina (Ureta et al. 2008). Finally, escaped populations observed in Spain show that such weedy forms can also leave the cultivated field and adapt to non arable places. Such evolution probably depends on the level of exploitation and artificialization of the agro-ecosystem. Around Cordoba, the landscape is more heterogeneous and more places are left unmanaged than in the Campiña region where farming is very intensive.
Compared to volunteers, the wild-type traits make these weedy forms highly invasive at a field level. Indeed, seed shattering and branching allow the enrichment of the seed bank with weeds or crop-weed hybrids, and the quick spread of the infestation within the fields: this rapid dynamic has been described by many farmers who have seen their fields become heavily infested in two or three generations. Also, dormant seeds can persist for years in the seed bank. Indeed, weedy sunflowers have been observed in some fields after more than seven years of absence (due to the cultivation of winter crops and use of appropriate herbicides in the meanwhile). These observations suggest that weed management should be conducted at the initial stage of the infestation, when the local population density are still low (Humston et al. 2005); when a threshold has been crossed, it becomes quite impossible to eliminate the weeds. By contrast with the within-field dynamics, the dispersion seems to be much slower between fields and from fields to uncultivated places. It indeed involves two phenomena that are less probable than those involved in within-field invasion: First seed transfer between fields is less frequent than seed shattering on the field. Second, pollen from the weeds to cultivated plants in a neighbouring field competes with pollen of the variety itself; moreover, even if it succeeds in fecundating a crop ovule, this will produce a seed on a head that does not shatter and has a high probability to be harvested. These events are however not impossible, as shown by some cases in the geographical survey.
These results raise questions that can only be addressed through detailed local surveys and molecular analysis. We plan now (1) to confirm the origin of these weedy populations using nuclear and cytoplasmic molecular markers and (2) to investigate the local dynamics and crop-weed genetic exchange through detailed multiyear surveys and sampling. These studies could help to predict the response of weedy populations to different kinds of weed management, the impact of crop-weed or weed-crop gene flow and to prevent new weed evolution, dispersion or seed stock contamination.
Finally, the processes involved in weed evolution make these plants interesting genetic resources. Indeed, in addition to the evolution of weediness itself, they involve an adaptation to new climate and local environmental conditions and to various biotic and abiotic stresses (Cantamutto et al. 2008).
References
Arnold ML (2004) Natural hybridization and the evolution of domesticated, pest and disease organisms. Mol Ecol 13:997–1007. doi:10.1111/j.1365-294X.2004.02145.x
Basu C, Halfhill MD, Mueller TC, Stewart CN Jr (2004) Weed genomics: new tools to understand weed biology. Trends Plant Sci 9:391–398. doi:10.1016/j.tplants.2004.06.003
Boudry P, Mörchen M, Saumitou-Laprade P, Vernet P, Van Dijk H (1993) The origin and evolution of weed beets: consequences for the breeding and release of herbicide-resistant transgenic sugar beets. Theor Appl Genet 87:471–478. doi:10.1007/BF00215093
Burke JM, Tang S, Knapp SJ, Rieseberg LH (2002) Genetic analysis of sunflower domestication. Genetics 161:1257–1267
Campbell LG, Snow AA, Ridley CE (2006) Weed evolution after crop gene introgression: greater survival and fecundity of hybrids in a new environment. Ecol Lett 9:1198–1209. doi:10.1111/j.1461-0248.2006.00974.x
Cantamutto M, Poverene M, Peinemann N (2008) Multi-scale analysis of two annual Helianthus species naturalization in Argentina. Agric Ecosyst Environ 123:69–74. doi:10.1016/j.agee.2007.04.005
Ellstrand NC (2003) Dangerous liaisons? When cultivated plants mate with their wild relatives. John Hopkins University Press, Baltimore
Faure N, Serieys H, Bervillé A (2002) Potential gene flow from cultivated sunflower to volunteer, wild Helianthus species in Europe. Agric Ecosyst Environ 89:183–190. doi:10.1016/S0167-8809(01)00338-3
Gandhi SD, Heesacker AF, Freeman CA et al (2005) The self-incompatibility locus (S) and quantitative trait loci for self-pollination and seed dormancy in sunflower. Theor Appl Genet 111:619–629. doi:10.1007/s00122-005-1934-7
Harter A, Gardner KA, Falush D et al (2004) Origin of extant domesticated sunflowers in eastern North. Am Nat 430:201–205
Holec J, Soukup J, Cerovska M, Novakova K (2005) Common sunflower (Helianthus annuus var. annuus) – potential threat to coexistence of sunflower crops in Central Europe. In: Proceedings 2th internal conference on co-existence between GM and non-GM based agricultural supply chains, Montpellier, France, 14–15 Nov 2005, pp 271–272
Humston R, Mortensen DA, Bjornstad ON (2005) Anthropogenic forcing on the spatial dynamics of an agricultural weed: the case of the common sunflower. J Appl Ecol 42:863–872. doi:10.1111/j.1365-2664.2005.01066.x
Kane NC, Rieseberg LH (2008) Genetics and evolution of weedy Helianthus annuus populations: adaptation of an agricultural weed. Mol Ecol 17:384–394. doi:10.1111/j.1365-294X.2007.03467.x
Londo JP, Schaal BA (2007) Origins and population genetics of weedy rice in the USA. Mol Ecol 16:4523–4535. doi:10.1111/j.1365-294X.2007.03489.x
Muller M-H, Arlie G, Bervillé A et al (2006) Le compartiment spontané du tournesol Helianthus annuus en Europe: prospections et premières caractérisations génétiques. Actes du Colloque BRG 6:335–353
Putt ED (1997) Early history of sunflower. In: Schneiter AA (ed) Sunflower technology and production. ASA, CSSA, SSSA, Madison, pp 1–19
Reagon M, Snow AA (2006) Cultivated Helianthus annuus (Asteraceae) volunteers as a genetic “bridge” to weedy sunflower populations in North America. Am J Bot 93:127–133. doi:10.3732/ajb.93.1.127
Ureta MS, Carrera AD, Cantamutto MA, Poverene MM (2008) Gene flow among wild and cultivated sunflower Helianthus annuus in Argentina. Agric Ecosyst Environ 123:343–349. doi:10.1016/j.agee.2007.07.006
Vischi M, Cagiotti ME, Cenci CA, Seiler GJ, Olivieri AM (2006) Dispersal of wild sunflower by seed and persistant basal stalks in some areas of central Italy. Helia 45:89–94
Acknowledgements
We thank Muriel Tavaud for careful reading of the manuscript. We thank the Bureau des Ressources Génétiques for funding this project.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Muller, MH., Délieux, F., Fernández-Martínez, J.M. et al. Occurrence, distribution and distinctive morphological traits of weedy Helianthus annuus L. populations in Spain and France. Genet Resour Crop Evol 56, 869–877 (2009). https://doi.org/10.1007/s10722-009-9409-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10722-009-9409-3