
Abstract
The current study aims to determine the healing activity of water soluble polysaccharide-rich fraction of a wild mushroom, Termitomyces eurhizus (TEps) against the indomethacin induced gastric ulceration in mice model. Gastric tissue histology, myeloperoxidase (MPO) activity, cyclooxygenases (COX) 1 and 2 expression, prostaglandin E2 (PGE2) synthesis, and modulation of pro/anti inflammatory cytokines expression were studied for this purpose. Histological study shows that TEps (20 mg/kg) effectively healed the gastric ulceration. Based on biochemical results, the healing capacities of TEps could be attributed to reduction of MPO activity and protection of mucosal mucin content. Enhanced synthesis of PGE2 by modulation of COX-1 and COX-2 expression and a prominent shift of cytokines expression from pro (TNF-α, IL-1ß) to anti inflammatory (IL-10) side are also held responsible for ulcer healing. The preliminary study highlights the anti-ulcerogenic property of polysaccharide-rich fraction of Termitomyces eurhizus and opens an alternative cure for NSAID induced gastroduodenal diseases.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Gastric ulcer disease is the common illness, affecting million of humanity yearly [1]. Gastric ulcer develop due to an imbalance between aggressive factors (H. pylori, NSAIDs, gastric acid) and protective factors (mucin, bicarbonate, prostaglandins) that leads to an interruption in the mucosal integrity [2]. Apart from Helicobacter pylori infection, non steroidal anti-inflammatory drug (NSAID) is one of the main causative factor for induction of gastric ulcer. NSAIDs are used worldwide for the treatment of pain, rheumatic arthritis, cardiovascular diseases and more recently, for the prevention of colon cancer and Alzheimer’s disease [3]. But long term use of the NSAIDs ultimately causes gastric ulcer. Twenty-five percent among total gastric ulcer patients are suffering from NSAIDs induced gastric ulcer [4–6]. Most modern antiulcer drugs, which are currently available in the market, show limited efficacy against gastric ulcer and are associated with severe side effects [1, 7]. A drug with multiple mechanism of protective action including antioxidant activity may be highly effective in minimizing tissue injury in human diseases. It has been observed that herbal drugs and formulations which possess potent antioxidant property are effective in healing experimentally induced gastric ulcer [1, 4, 8–10].
Mushroom had been used for traditional food and medicine in China, Japan, and other Asian countries since ancient time. Edible mushrooms are highly nutritious and could be used as medicine for the treatment of cancer, heart ailments, diabetes, inflammation, high blood pressure etc. [11–18].
Termitomyces eurhizus is a wild edible mushroom, which grow at costal and lateritic region of West Bengal in symbiotic association with Termites. It produces fruit bodies just after rainy season (late September to mid October) that earns a superior position as food among the edible mushroom because of its flavour, texture and gastronomic delicacy. The medicinal values of mushrooms are attributed mostly to their content of polysaccharide because of their biological response modification properties [19]. Recent investigation revealed that fungal polysaccharides have immunomodulatory, chemopreventive, hypoglycemic and anti-aging effect [20, 21]. Two major types of polysaccharide are generally present as the structural materials of mushrooms cell wall, among which the water soluble fraction mostly composed of glucans and glycoproteins. Mondal et al. (2004) have isolated two water soluble polysaccharides e.g., PSI and PSII from the hot water extract of fruiting bodies of T. eurhizus and characterized as PSI=(1-3)-D-Glcp and (1-6)-D-Glcp and PSII=(1-6)-D-Glcp [22]. However, medicinal properties of the T. eurhizus’s polysaccharide are not evaluated till date. Since water soluble polysaccharide-rich fraction is easy to isolate and can be taken as food without much purification steps. Hence, in the present study, an attempt has been made to evaluate antiulcerogenic activity of water soluble polysaccharide-rich fraction of T. eurhizus and compare its antiulcer effect with market available most usable drug, omeprazole. Omeprazole and TEps may not act at the same pathway to heal gastric ulcer, but some of the basic antiulcer parameters follow the same pathway. However, signaling molecules may be different. Omeprazole has a dual mechanism of action i.e., it inhibits both H+K+ATPase and gastric mucosa carbonic anhydrase enzymes [23].
Materials and methods
Chemicals
Indomethacin, Omeprazole, 5-bromo-4-chloro-3-indolyl phosphate (BCIP), nitroblue tetrazolium (NBT), tween-20, Bradford reagent, hexadecyl trimethyl ammonium bromide (HTAB), D-Glucose, Folin-Ciocalteau reagent, gallic acid were purchased from Sigma Chemical Co, Mo, St.Louis, USA; methanol from E. Merck, Mumbai, India; bovine serum albumin (BSA), from Merck, Darmstadt, Germany; dimethylformamide (DMF), tetramethyl benzidine (TMB) from Acros, Geel, Belgium; polyclonal rabbit anti-goat COX-1 and polyclonal goat anti-rabbit COX-2 antibodies from Santa Cruz biotechnology, Inc., Santa Cruz, CA; prostaglandin E2 EIA kit from Cayman Chemical. Cytokines TNF-α, IL-1ß and IL-10 ELISA kit from Pierce Biotechnology, Rockford, USA. Other reagents such as 35 % hydrogen peroxide (H2O2) from Lancaster, Morecambe, UK; disodium hydrogen phosphate and sodium dihydrogen phosphate from BDH, Poole, Dorset, UK; Mushroom ß-glucan kit was purchased from Megazyme Int., Wicklow, Ireland. All other chemicals used for this study were of analytical grade.
Animals
Male Swiss albino mice (6–8 weeks, 25 ± 2 g) bred in-house with free access to food and water were used. The mice were kept in 12-h light/dark cycles and housed at 25 ± 1 °C. The animal experiments were conducted in accordance with the guidelines of the Animal Ethical Committee, Institute of Postgraduate Medical Education and Research, Kolkata-20 (Animal Ethical Committee Sanction No:IAEC/SB-3/2008/UCM-64 Dated-15/05/08-2011) and were handled following the International Animal Ethics Committee Guidelines, ensuring minimum animal suffering.
Extraction of Polysaccharide-rich fraction from T. eurhizus basidiocarps
Basidiocarps of T. eurhizus were collected from the local market of South 24 parganas, West Bengal, India. Fresh mushrooms were cleaned and washed thoroughly with distilled water, freeze-dried and grinded to powder form. Powdered basidiocarps of T. eurhizus (100 g) were extracted with 80 % ethanol at room temperature for overnight. The extraction process was repeated for three times. The residual material was separated through linen cloth, dried and subsequently extracted with distilled water in a boiling water bath for 4 h. It was kept overnight at 4 °C and then filtered through linen cloth. The aqueous phase was evaporated and reduced to half of the volume and precipitated with 99 % ethanol (1:4 v/v). It was kept overnight at 4 °C and then centrifuged at 10,000 rpm for 20 min at 4 °C. After centrifugation, the pellets were washed with 70 % (v/v) ethanol and then sequentially washed with ethyl acetate and acetone [20, 21] and then air-dried. This air-dried water soluble crude polysaccharide-rich fraction (TEps) was kept at 4 °C for further use [24] (Appendix).
Physico-chemical characterization of the polysaccharide-rich fraction
The physico-chemical characterization comprises determination of the total carbohydrate, protein and phenol content; estimation of total glucan, ß glucan, α glucan content and FT-IR analysis of the crude polysaccharide-rich fraction.
Determination of polysaccharide content
The total polysaccharide content of the TEps was determined using phenol-sulfuric acid method with D-Glucose as a reference [25]. The reaction mixture consisted of 1 ml extract solution, 0.5 ml of 6 % phenol and 2.5 ml of concentrated sulfuric acid. It was allowed to stand for 30 min at room temperature and absorbance was taken at 490 nm. Standard curve was drawn using 10–100 μg of glucose and results were expressed as g of glucose equivalents/ 100 g of the dry polysaccharide.
Determination of glucan content
Contents of total glucan and α-glucan of TEps were determined using the Mushroom and Yeast ß-glucan Assay Procedure (Megazyme Int.) kit. The enzyme kit contains exo-1, 3-ß-glucanase, ß-glucosidase, amyloglucosidase, invertase, glucose determination reagent and glucose standard solution. To estimate the total glucan content, TEps was hydrolyzed with 37 % HCl for 1 h at room temperature and 2 h at 100 °C. After neutralization, a mixture of exo-ß-(1–3)-ß-glucanase plus ß-glucosidase in Na-acetate buffer (pH 5) was added to break ß1-3 as well as ß1-4 glucans and incubated at 40 °C. Glucose oxidase / peroxidase reagent was added to oxidize glucose which can be visualized spectrophotometrically. The α-glucan content was estimated upon enzymatic hydrolysis with amyloglucosidase plus invertase yielding glucose. Glucose oxidase / peroxidase reagent was added to determine free glucose. The ß glucan content was calculated by subtracting α-glucan from total glucan content according to kit’s manual. All values of glucan content were expressed as g/ 100 g of dry weight of the TEps.
Determination of protein content
Protein concentration of TEps was determined using the method described by Bradford (1976) [26]. 750 μl of sample solution was added with 750 μl of Bradford reagent and incubated at room temperature for 5 min. Absorbance was measured at 595 nm and BSA (5–50 μg) was used to produce standard calibration curve. Total protein content of the extracts was expressed as g of BSA equivalents/ 100 g of the TEps.
Determination of phenol content
The total phenolic compounds present in the TEps were estimated as described by Ziestin and Ben-Zaker (1993) [27]. Briefly, 0.5 ml extract solution was mixed with 2.5 ml distilled water and 125 μl of 1(N) Folin-Ciocalteau reagent. Reaction mixture was incubated for 30 min in dark and absorbance was taken at 725 nm. Gallic acid was used to calculate the standard curve (10–50 μg). Total phenol was expressed as g of gallic acid equivalents/ 100 g of TEps.
FT-IR analysis of TEps
The FT-IR spectra of TEps were recorded on a Perkin Elmer Precirety, Spectrum 100 Model. The crude polysaccharide were ground with potassium bromide powder and then pressed into pellets for FT-IR measurements in the frequency range 400–4,000 cm−1.
Preparation of test samples
The test samples (TEps and Omeprazole) were prepared as aqueous suspension in 2 % gum acacia as the vehicle and administered to the mice orally.
Experimental protocol for ulceration and assessment of healing
The mice were divided into several groups (six mice per group) and each experiment was repeated three times. Except for the normal control group, ulceration in the other mice was induced by indomethacin (18 mg/kg, p. o., single dose), dissolved in distilled water and suspended in 2 % gum acacia as the vehicle [4, 8, 9]. The animals were deprived of food but had free access to tap water for 24 h, before ulcer induction. The normal and untreated control groups received the vehicle only throughout the course of the experiments. The treatment groups received different doses of TEps (1, 10, 20 and 40 mg/kg once daily) and Omeprazole (Omp) (3 mg/kg, once daily) [28] as positive control for different periods, with the first dose started at 6 h after indomethacin administration. On the 1st, 2nd, 3rd, 4th, and 7th day, the mice were sacrificed by cervical dislocation under anesthesia (ketamine, 12 mg/kg). Here, only 3rd day data is incorporated. The stomachs from the normal and treated groups were removed rapidly, opened along the greater curvature, and thoroughly rinsed with normal Saline.
Histology and study of damage score
The fundic portion of stomach was sectioned for histological studies as well as damage score analysis. The tissue samples were fixed in 10 % formalin and embedded in paraffin. The sections (5 μm) were cut using microtome, stained with hematoxylin and eosin and assessed under an Olympus microscope (BX41, Hamburg, Germany). From the histological slides, the damage scores were assessed [29] by grading the gastric injury on a 0–4 scale, based on the severity of hyperemia and hemorrhagic erosions: 0–almost normal mucosa, 0.5–hyperemia, 1–one or two lesions, 2–severe lesions, 3–very severe lesions and 4–mucosa full of lesions (lesions–hemorrhagic erosions, hyperemia-vascular congestions). The sum of the total scores divided by the mean damage score is expressed as the damage score. The experiments were performed by two investigators blinded to the groups and the treatment of animals.
Myeloperoxidase assay
Myeloperoxidase (MPO) activity was determined according to standard procedure [30] with minor modifications. In-brief whole gastric glandular portions of the stomach taken from all groups (100–150 mg) were homogenized in a 50 mM phosphate buffer (pH 6.0) containing 0.5 % HTAB. This was followed by three cycles of freezing and thawing. The homogenate was centrifuged at 12,000 × g for 20 min at 4 °C. The supernatant (50 μl) was collected for MPO assay and added to 80 mM phosphate buffer, pH 5.4, 30 mM TMB and 300 mM H2O2, to make a final reaction volume of 500 μl. After the mixture was incubated at 25 °C for 25 min, the reaction was terminated by adding 500 mM H2SO4 and the change in the absorbance was measured at 450 nm.
Mucin assay
Following a reported method [31], the free mucin in the gastric tissues was estimated. Briefly, the glandular portion of the stomach was separated from the lumen of the stomach, weighed, and transferred immediately to 10 ml of 0.1 % w/v Alcian Blue (AB) solution (in 160 mM sucrose solution buffered with 0.05 mM sodium acetate solution, pH was adjusted to 5.8). After staining for 2 h, the excess dye was removed from the tissue by two successive rinses with 10 ml of 250 mM sucrose solution. The dye, complexed with the gastric wall mucus, was extracted with 10 ml of 500 mM magnesium chloride by intermittent shaking (1 min) at 30-min intervals for 2 h. The blue extract (2 ml) was vigorously shaken with an equal volume of diethyl ether. The resulting emulsion was centrifuged at 3,600 rpm for 10 min, and the absorbance of the aqueous layer was read at 580 nm. The quantity of AB extracted per gram of wet glandular tissue was calculated from a standard curve prepared using various concentrations of AB.
Western blot analysis of COX enzymes
The glandular part of the gastric mucosa was washed with PBS containing protease inhibitors. Then the tissue was minced and homogenized in a lysis buffer (1 ml) containing 200 μl of 5X Tris-EDTA buffer, 200 μl of 5 X NaCl, 200 μl of 5 X SDS, 200 μl of 5 X deoxycholic acid, 200 μl of 5 X Igepal, and 10 μl of protease inhibitor cocktail. The homogenized tissue samples were homogenized and centrifuged at 12,000 × g for 10 min, at 4 °C and the protein concentration of the supernatant was measured by using commercially available Bradford reagent [26]. The proteins were resolved by 10 % non-reducing SDS polyacrylamide gel electrophoresis and transferred to nitrocellulose membrane. The membrane was blocked for 2 h at room temperature in 20 mM Tris-HCl, pH 7.4, 150 mM NaCl, and 0.02 % Tween 20 (TBST) containing 3 % BSA.
Overnight incubation of the membrane at 4 °C in primary antibody (1:500 dilution) of COX-1, COX-2 and ß-actin in TBST buffer containing 3 % BSA was undertaken. The membrane was washed three times with TBST buffer and thereafter incubated with alkaline phosphatase-conjugated secondary antibody. Then the membrane was washed again three times with TBST buffer and finally the bands were visualized using 5-bromo-4-chloro-3-indolyl phosphate / nitro blue tetrazolium substrate solution.
PGE2 assay
Following the harvesting of the stomach, the corpus (full thickness) was excised, weighed (100 mg), and suspended in 10 mM sodium phosphate buffer, pH 7.4 (1 ml). The tissues were finely minced and incubated at 37 °C for 20 min. After centrifugation (9,000 × g), the PGE2 levels in the supernatant were measured by ELISA, using the Prostaglandin E2 EIA kit (Cayman Chemical) instructions.
Estimation of tissue cytokines level
The TNF-α, IL-1ß and IL-10 levels in the tissue homogenate were estimated using commercially available ELISA kits, according to the manufacturer’s protocol.
The glandular part of the gastric mucosa after being washed with PBS containing protease inhibitors, was minced and homogenized in a lysis buffer (10 mM Tris-HCl pH 8.0, 150 mM NaCl, 1 % Triton X-100) containing leupeptin (2 μg/ml) and PMSF (0.4 μM). Then the homogenate was centrifuged at 15,000 × g for 30 min at 4 °C. Then supernatant was collected and cytokines levels were measured. The samples along with the standards were seeded to each well at an appropriate dilution and incubated at room temperature for 90 min. The wells were washed (5 times), diluted polyclonal antibody (100 μl) was added, and the mixture was incubated further for 2 h at room temperature. The wells were washed again and incubated for 2 h after addition of HRP conjugated (100 μl) secondary antibody. After the final wash, TMB (100 μl) was added to each well, the mixture was incubated for 15 min, the reaction was stopped by addition of 0.16 M H2SO4, and the absorbance was measured at 450 nm.
Statistical analysis
Data are expressed as mean±S.D. unless mentioned otherwise. Comparisons were made between different treatments using one way ANOVA followed by an error protecting multiple comparison procedure, namely Tukey-Kraemer post hoc test by Graph Pad InStat (GraphPad Software Inc., San Diego, CA) software for the analysis of significance of all the data.
Results
Extraction yields
Following hot water extraction, the yield of crude polysaccharide-rich fraction from T. eurhizus was 1.51 g/ 100 g of mushroom dry weight.
Physico-chemical characterization
The TEps appeared to be whitish powder which is highly soluble in water. The chemical composition of TEps was summarized in Table 1. Total carbohydrate, protein and phenol content were 36.3 %, 2.83 % and 9.5 %, respectively. Total glucan content was 17.81 g/ 100 g of polysaccharide. The α-glucan and ß-glucan content were 0.79 g and 17.02 g/ 100 g of polysaccharide respectively. In most mushrooms α glucan was found at levels below 1 % whereas ß glucan ranges from 4.71 to 46.2 % on a dry weight basis [20].
FT-IR analysis of TEps
In order to investigate structure of polysaccharides such as monosaccharide types, glucosidic bonds and functional groups FT-IR spectroscopy was done [32]. The FT-IR spectra showed typical carbohydrate pattern (Fig. 1). A strong band near the region of 3,429 cm-1 was characteristic of carbohydrate ring assuming the presence of OH stretching in hydrogen bonds, which was indicative of strong inter and intra-molecular interaction of the polysaccharide chains [33]. Absorption at 2,922 cm−1 indicated CH2 stretching. Peak of amide band at 1,631 cm−1 indicated the presence of some residual protein in the crude polysaccharide mixture. The band between 1,310 and 1,410 cm−1 i.e., 1,384 cm−1 corresponds to OH group of phenolic compound. Absorption region at 1,076 and 1,066 cm−1 was characteristic of the presence of ß-glucan due to O-substituted glucose residue [20].

FT-IR Spectral analysis of Polysaccharide Fraction of T. eurhizus
Histological assessment
Onset of ulceration (i.e., superficial erosion and mild inflammation in the stomach) was observed within 6 h after indomethacin administration, indicating acute ulceration (figure not shown). However, on the 3rd day, marked damage to the glandular portion of the gastric mucosa was noticed in the histological photograph of the stomach sections of the 3rd day-ulcerated group of mice. Hemorrhagic serosa was evident on the 3rd day of ulceration. Previous reports [34, 35] have already highlighted some of the following key players which are responsible for the inflammation viz. neutrophils, eosinophils, monocyte, macrophage and mast cell. Among them, neutrophils and macrophage cells are very important for the study of inflammatory response [36]. In comparison to normal, the ulcerated untreated stomach showed much more neutrophils sloughing in eroted area of the mucosa which loss the superficial epithelium. Treatment with TEps and Omp for 3 days reduced the number of inflammatory cells and mucosal congestion, and increased the number of healthy normal cells in the gastric mucosa, submucosa, serosa, and muscle layers. To verify this data further we have studied myeloperoxidase activity in tissue which is a marker of neutrophil infiltration [4, 8, 9, 37].
The healing effect of TEps was better than Omp. The histological photographs of stomach sections of the 3rd day-groups of normal, ulcerated, and treated mice are shown in Fig. 2.

Histological assessment of acute gastric mucosal injury induced by indomethacin in mice and its healing by TEps and Omp. Ulceration in the mice was induced by indomethacin (18 mg/kg, single dose, p.o.). Treatment was carried out with TEps (20 mg/kg once daily × 3 days, p.o.) and Omp (3 mg/kg once daily × 3 days, p.o.). Representative gastric tissue sections are shown at 10X magnifications. Mucosal and submucosal layers are shown by grey and black arrows, respectively. [a. Control; b. Ulcerated Untreated; c. Ulcerated + TEps Treated; d. Ulcerated + Omeprazole Treated]
Assessment of ulcer healing
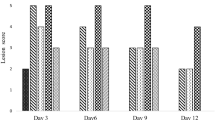
The mice receiving only the vehicle, showed no lesion in the gastric mucosa. Indomethacin (18 mg/kg, p.o., single dose) administration produced acute mucosal lesions in the mice stomach, as evident from damage score analysis. Quantification of the damage scores revealed maximum ulcerative damage on the 3rd day of indomethacin administration. However, the ulcerative damage was reduced on the 7th day (data not shown). Among the chosen doses of TEps, best ulcer healing effect was observed at a dose of 20 mg/kg once daily × 3 days, p.o. (Fig. 3a) at peak ulceration. Compared to the untreated group, the damage scores in the TEps and Omp-treated groups were reduced by 4.61 (p < 0.001) and 2.4 (p < 0.01) fold, respectively.

Ulcer Index (a) and MPO activity (b): Dose-dependent healing capacities of TEps at 3rd day of ulceration. Ulceration in the mice was induced by indomethacin (18 mg/kg, single dose, p.o.). Treatment was carried out with TEps (1, 10, 20, 40 mg/kg, single dose daily up to 3rd days, p. o.) after ulcer induction. The section of mice stomachs were dissected on the 3rd day of ulceration. The values are mean±S. D. **P < 0.01, ***P < 0.001, compared to ulcerated untreated mice
Regulation of mucosal MPO activity
The mucosal myeloperoxidase (MPO) activity of the indomethacin-administered mice, increased immediately, reaching the peak value on the 3rd day (Fig. 3b). The results were consistent with the damage score data. Treatment with TEps (20 mg/kg once daily × 3 days, p.o.) and Omp reduced the MPO activity by 68.41 % (p < 0.001) and 61.49 % (p < 0.001), respectively, compared to that of the untreated group (Fig. 3b).
Modulation of COX expression
The Western blots of COX-1 and COX-2 expressions in the gastric mucosa of the sham-treated / control, ulcerated untreated and drug (TEps or Omp)-treated mice are shown in Fig. 4a. Ulceration significantly depleted the expression of gastric COX-1 compared to the normal group. However, COX-2 expression was not altered by indomethacin. Treatment with TEps in ulcerated group elevated COX-1 and COX-2 expressions significantly, compared to the ulcerated-untreated group. In contrast, the effect of Omp in ulcerated group was less prominent than TEps.

Effect of TEps on mucosal COX-1 and COX-2 expressions (a) and PGE2 synthesis (b) in indomethacin (18 mg/kg, single dose, p.o.)—induced ulcerated mice. The values are mean±S. D. **P < 0.01, ***P < 0.001, compared to ulcerated untreated mice
Regulation of PGE2 synthesis
Compared to the normal control, the mucosal PGE2 level was markedly suppressed (2.66 fold) (p < 0.001) in the untreated-ulcerated mice. Treatment with either TEps or Omp for consecutive 3 days up-regulated the mucosal PGE2 level by 2.30 (p < 0.001) and 1.7 (p < 0.01) fold, respectively, compared to the untreated group (Fig. 4b).
Mucin assay
Compared to the normal control, the mucin content was markedly suppressed (1.63 fold) (p < 0.01) in the untreated-ulcerated mice. Treatment with either TEps or Omp for consecutive three days up-regulated the mucin content by 39.89 % (p < 0.01) and 19.1 % (p < 0.05) respectively, compared to the untreated group (Fig. 5).

Effect of TEps (20 mg/kg, single dose daily × 3 days, p. o.) on mucosal mucin synthesis in indomethacin (18 mg/kg, single dose, p.o.)—induced ulcerated mice. The values are mean±S. D. *P < 0.05, **P < 0.01 compared to ulcerated untreated mice
Effect on cytokines
Indomethacin administration modulated the balance of pro/anti-inflammatory cytokines levels. ELISA study depicted that indomethacin treatment induced TNF-α (2.08 fold, p < 0.001), IL-1ß (6.40 fold, p < 0.001), IL-10 (1.08 fold, p < 0.05) levels in compared to normal control. However, TEps at its dose 20 mg/kg, significantly reduced the pro inflammatory cytokines (TNF-α, IL-1ß) levels (2.21, p < 0.001 and 1.53, p < 0.01 fold respectively) (Fig. 6a–b) and induced anti-inflammatory (IL-10) levels (1.81, p < 0.001 fold) (Fig. 7) compared to the ulcerated-untreated mice.

Effect of TEps (20 mg/kg, single dose daily × 3 days, p. o.) on different cytokines levels in indomethacin (18 mg/kg, single dose, p.o.).—induced ulcerated mice. a tissue TNF-α level; b tissue IL-1ß level. The values are mean±S. D. **P < 0.01, ***P < 0.001, compared to ulcerated untreated mice

Effect of TEps (20 mg/kg, single dose daily × 3 days, p. o.) on antiinflammatory cytokine (IL-10) in indomethacin (18 mg/kg, single dose, p.o.)—induced ulcerated mice. The values are mean±S. D. ***P < 0.001, compared to ulcerated untreated mice
Discussion
In recent days, gastric ulcer is a serious global problem. Gastric ulcers have been described as an imbalance of the luminal acid secretions and the mucosal defenses. Stress, smoking, nutritional deficiencies, H. pylori infection and ingestion of NSAIDs are the major factors of gastric ulcer formation [4, 5, 8, 9, 38].
The conception of gastric ulcer management is changing fast. Gastric ulcer therapy faces a major drawback in modern medicine due to the unpredictable side effects of the long-term use of commercially available drugs [39]. Hence, the search is still on to find drugs, which will serve as a powerful therapeutic agent to heal gastric ulceration [4, 8, 9, 40].
Mushrooms have been part of the human diet for thousands of years because of their high nutritional value and medicinal properties. Some of the current reports already have informed that the wild and edible mushrooms harbor many bioactive molecules that exhibit interesting biological activities, including providing remedies for gastric ulcer and chronic gastritis [1, 39, 41]. In our lab we evaluated the healing efficacy of a wild mushroom, Termitomyces eurhizus against the indomethacin-induced gastric ulceration in mice, and assessed their role in mucosal defense system. Our present study highlighted that the water soluble polysaccharide-rich fraction of the T. eurhizus showed potent healing effect on indomethacin-induced gastric ulceration.
Our histological results showed marked gastric mucosal damage in mice, on the 3rd day after indomethacin administration. The partial auto healing observed in the ulcerated untreated mice. TEps showed impressive mucosal healing. Under an optimized three-day treatment regime, TEps (20 mg/kg) and Omp (3 mg/kg) produced similar ulcer healing (Figs. 2 and 3a).
To correlate the histological and damage score data we have assessed the MPO activity. The MPO activity is known to increase under ulcerated conditions and reduced during the healing process [42]. It is often used as a risk marker and diagnostic tool for assessing severity of gastric ulcer [43]. Treatment with TEps sufficiently restored the normal gastric mucosal integrity, while reducing the MPO activity (Fig. 3b).
The NSAIDs exert both their therapeutic and toxic effects mainly by decreasing the levels of circulating PGE at the gastric mucosa via inhibition of the COX isozymes. The reduced level of PGs is known to cause gastric ulceration and also exacerbate preexisting gastric ulcers in rodents and humans [44, 45]. PGs stimulate mucus and bicarbonate secretion as well as mucosal blood flow and induce angiogenesis [46].
All these factors contribute to accelerated ulcer healing. Our western blots revealed reduced expressions of COX-1 and COX-2 at peak ulceration (Fig. 4a), associated with reduced synthesis of mucosal PGE (Fig. 4b). Treatment with TEps increased the expressions of both the enzymes. Omp showed slightly less effect on these enzymes. The test samples augented the mucosal PGE that correlated well with their respective abilities to regulate the expressions of the COX isoforms.
Depletion of gastric mucosal mucin level also contributes to the NSAID-mediated gastropathy. Maintenance of mucus production may provide partial but significant protection against reactive oxygen metabolites. Our results revealed that stomach ulceration reduced the gastric mucin content. This might reduce the ability of the mucosal membrane to protect the mucosa from physical damage and back diffusion of hydrogen ions [47], and hinder epithelial recovery. Treatment with TEps and Omp significantly accelerated ulcer healing by increasing the mucin content of the gastric mucosa (Fig. 5).
In mucosal defense cytokines also play a central role in the regulation of the mucosal immune system and are, therefore, extremely important. The role of cytokines in GI inflammation and injury-healing has recently been reviewed in detail [48]. Inflammatory cytokines are extremely important in mucosal defense. One of the most prominent modes of medication of indomethacin-induced gastropathy is the increased expression of the pro-inflammatory cytokines [49, 50], which also correlates with the extent of ulceration. After trauma, the Th1/Th2 imbalance with Th2 predominance is reflected by an increase of anti-inflammatory cytokines such as IL-10 [51]. In view of this, the immune response due to ulceration and its modulation by the TEps was monitored. This enabled us to associate the inflammatory response with a better prognosis. Indomethacin administration raised the levels of pro-inflammatory Th1 cytokines (TNF-α and IL-1ß) and reduced the anti-inflammatory cytokine, IL-10, as reported earlier [52]. Given that IL-10 is stimulated by PGE2, we also observed reduction of IL-10 on indomethacin administration, as reported earlier [53]. All these are likely to promote oxidative stress and result in the formation of ulceration [54].
The enhancement of the levels of soluble inflammatory modulators by indomethacin reflected a pro-inflammatory trigger. Treatment with TEps reversed the imbalance by lowering the Th1 cytokines drastically (Fig. 6a–b), and restoring the level of IL-10 to normalcy (Fig. 7). This is consistent with their respective effects in healing gastric ulcer as well as altering the other biochemical parameters. The upregulation of the anti-inflammatory cytokines by the TEps is likely to inhibit the stimulatory effect of indomethacin on the release of the pro-inflammatory cytokines in tissue. TEps was also found to augent the PGE level in the indomethacin-treated mice. The increased PGE might stimulate IL-10 release, which, in turn, controls PGE and Th1 cytokines via a negative feedback.
However, the role of cytokine imbalance in gastropathy has not been adequately emphasized. Our results showed that the balance of the pro- and anti-inflammatory cytokines could play a significant role in the NSAID-induced gastric mucosal injury. Thus all the above studies suggested that TEps healed the gastric lesion by upregulating the mucin, PGE as well as cyclooxygenase and the balance of Th1/Th2 in favour of Th2.
Overall, our results clearly revealed the healing ability of TEps against indomethacin-induced stomach ulceration. The results are consistent with the anti-inflammatory property of TEps. Based on the available evidence, the therapeutic effect of this test sample could be related to their mucin-protecting, COX and PGE-enhancing properties.
References
Omar, N.A.M., Abdullah, N., Kuppusamy, U.R., Abdulla, M.A., Sabaratnam, V.: Nutritional Composition,Antioxidant Activities, and Antiulcer Potential of Lentinus squarrosulus (Mont.) Mycelia Extract. Evid-Based Complement Alternat Med Article ID 539356. 8 pages (2011)
Debabrata, M., John, A.: Peptic ulcers and their complications. Surgery 24(3), 110–114 (2006)
Sung, J.J., Leung, W.K., Go, M.Y., To, K.F., Cheng, A.S., Ng, E.K., Chan, F.K.: Cyclooxygenase-2 expression in Helicobacter pylori-associated premalignant and malignant gastric lesions. Am. J. Pathol. 157(3), 729–735 (2000)
Chatterjee, A., Chattopadhyay, S., Bandyopadhyay, S.K.: Biphasic Effect of Phyllanthus emblica L. Extract on NSAID-Induced Ulcer: An Antioxidative Trail Weaved with Immunomodulatory Effect. Evid-Based Complement Alternat Med (2011), 7 pages. Article ID 146808
Chatterjee, A., Chatterjee, S., Bandyopadhyay, S.K.: H. pylori-induced gastric ulcer: pathophysiology and herbal remedy. Int. J. Biol. Med. Res. 31, 1461–1465 (2012)
Dhikav, V., Singh, S., Pande, S., Chawla, A., Anand, K.S.: Non-steroidal drug-induced gastrointestinal toxicity: mechanisms and management. Indian J. Clin. Med. 4, 315–322 (2003)
Goel, R.K., Sairam, K.: Anti-ulcer drugs from indigenous sources with emphasis on Musa sapientum. Tamrabhasma. Asparagus racemosus and Zingiber officinale. Indian J. Pharmacol. 34, 100–110 (2002)
Chatterjee, A., Chatterjee, S., Das, S., Saha, A., Chattopadhyay, S., Bandyopadhyay, S.K.: Ellagic acid facilitates indomethacin-induced gastric ulcer healing via COX-2 up-regulation. Acta. Biochim. Biophys. Sin. 44, 565–576 (2012)
Chatterjee, A., Chatterjee, S., Biswas, A., Bhattacharya,S., Chattopadhyay, S., Bandyopadhyay, S.K.: Gallic Acid enriched fraction of Phyllanthus emblica, potentiates NSAID-induced gastric ulcer healing via eNOS dependent pathway. Evid-Based Complement Alternat Med (2012), 13 pages. Article ID 487380
Lehne, R.A., Saunders, W.B.: Antacids, Pharmacology for Nursing Care. Philadelphia, Pa, USA. 3, 781–783 (1998)
Acharya, K., Acharya, S.N., Thomas, J.E.: Medicinal properties of mushroom. Advances in medicinal plant research Signpost, Kerala (India), 215–236 (2007)
Chang, R.: Functional properties of edible mushroom. Nutr. Rev. 54, 91–93 (1996)
Chatterjee, S., Biswas, G., Basu, S.K., Acharya, K.: Antineoplastic effect of mushrooms: a review. Aust. J. Crop. Sci. 5, 904–911 (2011)
Biswas, G., Chatterjee, S., Sarkar, S., Acharya, K.: Chemopreventive activity of the ethanolic extract of Astraeus hygrometricus (Pers.) Morg. On Ehrlich’s Ascites Carcinoma Cell. Dig. J. Nanomater Bios. 7, 185–191 (2012)
Chatterjee, S., Biswas, G., Chandra, S., Saha, G.K., Acharya, K.: Apoptogenic effects of Tricholoma giganteum on Ehrlich’s ascites carcinoma cell. Bioproc. Biosystems Eng. 36, 101–107 (2013)
Chatterjee, S., Dey, A., Datta, R., Dey, S., Acharya, K.: Hepatoprotective effect of the Ethanolic Extract of Calocybe indica on Mice with CCl4 Hepatic Intoxication. Int. J. Pharmtech. Res. 3, 2162–2168 (2011)
Acharya, K., Chatterjee, S., Biswas, G., Chatterjee, A., Saha, G.K.: Hepatoprotective effect of a wild edible mushroom on carbon tetrachloride-induced hepatotoxicity in mice. Int. J. Pharm. Pharm. Sci. 4, 285–288 (2012)
Biswas, G., Rana, S., Sarkar, S., Acharya, K.: Cardioprotective activity of ethanolic extract of Astraeus hygrometricus (Pers.) Morg. Pharmacologyonline 2, 808–817 (2011)
Wasser, S.P.: Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. Appl. Microbiol. Biotechnol. 60, 258–274 (2002)
Kozarski, M., Klaus, A., Niksic, M., Jakovljevic, D., Helsper, J.P.F.G., Griensven, L.J.L.D.V.: Antioxidative immunomodulating activities of polysaccharide extracts of the medicinal mushrooms.Agaricus bisporus, Agaricus brasiliensis, Ganoderma lucidum Phellinus linteus. Food Chem. 129, 1667–1675 (2011)
Luo, A., He, X., Zhou, S., Fan, Y., Luo, A., Chun, Z.: Purification, composition analysis antioxidant activity of the polysaccharides from Dendrobium nobile Lindl. Carbohydr. Polym. 79, 1014–1019 (2010)
Mondal, S., Chakraborty, I., Pramanik, M., Rout, D., Islam, S.S.: Structural studies of water-soluble polysaccharides of an edible mushroom, Termitomyces eurhizus. A reinvestigation. Carbohydr. Res. 339, 1135–1140 (2004)
Puscus, I., Coltau, M., Baican, M., Domuta, G.: Omeprazole has a dual mechanism of action: it inhibits both H + K + ATPase and gastric mucosa carbonic anhydrase enzyme in humans (in vitro and in vivo experiments). J. Pharmacol. Exp. Therpeut. 290, 530–534 (1999)
Cui, Y., Kim, D.S., Park, K.C.: Antioxidant effect of Inonotus obliquus. J. Ethnopharmacol. 96, 79–85 (2005)
Dubois, M., Gilles, A., Hamitton, J.K., Reders, P.A., Smith, F.: Colorimetric method for determination of sugars related substances. Anal. Chem. 28, 350–356 (1956)
Bradford, M.M.: A refined sensitive method for the quantitation of microgram quantities of protein using the principle of protein-dye binding. Anal. Biochem. 72, 248–255 (1976)
Ziestin, N., Ben-Zaker, R.: Peroxidase activity presence of phenolic substance in peduncles of rose flowers. Plant Physiol. Biochem. 31, 333–339 (1993)
Banerjee, D., Bauri, A.K., Guha, R.K., Bandyopadhyay, S.K., Chattopadhyay, S.: Healing properties of malabaricone B and malabaricone C, against indomethacin-induced gastric ulceration and mechanism of action. Eur. J. Pharmacol. 578, 300–312 (2008)
Dokmeci, D., Akpolat, M., Aydogdu, N., Doganay, L., Turan, F.N.: L-carnitine inhibits ethanol-induced gastric mucosal injury in rats. Pharmacol. Rep. 57, 481–488 (2005)
Suzuki, K., Ota, H., Sasagawa, S.: Assay method for myeloperoxidase in human polymorphonuclear leukocytes. Anal. Biochem. 132, 345–352 (1983)
Tariq, M., Moutaery, A.A.: Menadione protects gastric mucosa against ethanol-induced ulcers. Exp. Toxicol. Pathol. 56, 393–399 (2005)
Yang, L., Zhang, L.M.: Chemical structural chain conformational characterization of some bioactive polysaccharides isolated from natural sources. Carbohydr. Pol. 76, 349–361 (2009)
Mathlouthi, M., Koenig, J.L.: Vibrational spectra of carbohydrates. Adv. Carbohydr. Chem. Biochem. 44, 789 (1986)
Serhan, C.N., Brain, S.D., Buckley, C.D., Gilroy, D.W., Haslett, C., O’Neill, L.A.J., Perretti, M., Rossi, A.G., Wallace, J.L.: Resolution of inflammation: state of the art, definitions and terms. FASEB J. 21, 325–332 (2007)
Frasch, S.C., Fernandez-Boyanapalli, R.F., Berry, K.A.Z., Murphy, M., Leslie, C.C., Nick, J.A., Henson, P.M., Bratton, D.L.: Neutrophils regulate tissue neutrophilia in inflammation via the oxidant-modified lipid lysophosphatidylserine. J. Biol. Chem. 288, 4583–4593 (2013)
Adhikary, B., Yadav, S.K., Bandyopadhyay, S.K., Chattopadhyay, S.: Epigallocatechin gallate accelerates healing of indomethacin-induced stomach ulcer in mice. Pharmacol. Rep. 63, 527–536 (2011)
La Casa, C., Villegas, I., Alarcón de la Lastra, C., Motilva, V., Martín Calero, M.J.: Evidence for protective and antioxidant properties of rutin, a natural flavone, against ethanol induced gastric lesions. J. Ethnopharmacol. 71, 45–53 (2000)
Belaiche, J., Burette, A.D., Vos, M., Louis, E., Huybrechts, M., Deltenre, M.: Study group of NSAID-GI complications, observational survey of NSAID related upper gastrointestinal adverse events in Belgium. Acta Gastroenterol. Belgica 65, 65–73 (2002)
Rony, K.A., Mathew, J., Neenu, P.P., Janardhanan, K.K.: Ganoderma lucidum (Fr.) P. Krast occurring in south India attenuates gastric ulceration in rats. Indian J. Natl. Prod. Res. 2, 19–27 (2011)
Zhao, W., Zhu, F., Shen, W., Fu, A., Zheng, L., Yan, Z., Zhao, L., Fu, G.: Protective effects of DIDS against ethanol-induced gastric mucosal injury in rats. Acta. Biochim. Biophys. Sin. 41, 301–308 (2009)
Gao, Y., Zhou, S., Wen, J., Huang, M., Xu, A.: Mechanism of the antiulcerogenic effect of Ganoderma lucidum polysaccharides on indomethacin-induced lesions in the rat. Life Sci. 72, 731–745 (2002)
Souza, M.H.L.P., Lemos, H.P., Oliveira, R.B., Cunha, F.Q.: Gastric damage and granulocyte infiltration induced by indomethacin in tumour necrosis factor receptor 1 (TNF-R1) or inducible nitric oxide synthase (iNOS) deficient mice. Gut 53, 791–796 (2004)
Krawisz, J.E., Sharon, P., Stenson, W.F.: Quantitative assay for acute intestinal inflammation based on myeloperoxidase activity. Gastroenterol. 87, 1344–1350 (1984)
Halter, F., Tarnawski, A.S., Schmassmann, A., Peskar, B.M.: Cyclooxygenase 2 -implications on maintenance of gastric mucosal integrity and ulcer healing: controversial issues and perspectives. Gut 49, 443–453 (2001)
Rainsford, K.D.: The effects of aspirin and other non-steroid anti-inflammatory/analgesic drugs on gastrointestinal mucus glycoprotein biosynthesis in vivo: relationship to ulcerogenic actions. Biochem. Pharmacol. 27, 877–885 (1978)
Wallace, J.L.: Nonsteroidal anti-inflammatory drugs and gastroenteropathy: the second hundred years. Gastroenterol. 112, 1000–1016 (1997)
Jones, M.K., Wang, H., Peskar, B.M., Levin, E., Itani, R.M., Sarfeh, I.J., Tarnawski, A.S.: Inhibition of angiogenesis by nonsteroidal anti-inflammatory drugs: insight intomechanisms and implications for cancer growth and ulcer healing. Nat. Med. 5, 1418–1423 (1999)
Playford, R.J., Ghosh, S.: Cytokines and growth factor modulators in intestinal inflammation and repair. J. Pathol. 250, 417–425 (2005)
Yoshikawa, T., Naito, Y., Kishi, A., Tomii, T., Kaneko, T., Iinuma, S., Ichikawa, H., Yasuda, M., Takahashi, S., Kondo, M.: Role of active oxygen, lipid peroxidation, antioxidants in the pathogenesis of gastric mucosal injury induced by indomethacin in rats. Gut 34, 732–737 (1993)
Brzozowski, T., Konturek, P.C., Konturek, S.J., Sliwowski, Z., Pajdo, R., Drozdowicz, D., Ptak, A., Hahn, E.G.: Classic NSAID and selective cyclooxygenase COX-1 and COX-2 inhibitors in healing of chronic gastric ulcers. Microsc. Res. Tech. 43, 343–353 (2001)
Modolell, M., Corraliza, I.M., Link, F., Soler, G., Eichmann, K.: Reciprocal regulation of the nitric oxide synthase/arginase balance in mouse bone marrow derived macrophages by TH1 and TH2 cytokines. Eur. J. Immunol. 25, 1101–1104 (1995)
Slomiany, B.L., Piotrowski, J., Slomiany, A.: Role of endothelin-1 and constitutive nitric oxide synthase in gastric mucosal resistance to indomethacin injury: effect of antiulcer agents. Scand. J. Gastroenterol. 34, 459–464 (1999)
Rhind, S.G., Gannon, G.A., Shephard, R.J., Shek, P.N.: Indomethacin modulates circulating cytokine responses to strenuous exercise in humans. Cytokine 19, 153–158 (2002)
Chatterjee, S., Premachandran, S., Bagewadikar, R.S., Bhattacharya, S., Chattopadhyay, S., Poduval, T.B.: Arginine metabolic pathways determine its therapeutic benefit in experimental heat stroke: role of Th1/ Th2 cytokine balance. Nitric Oxide 15, 408–416 (2006)
Acknowledgement
Authors like to acknowledge Director, I.P.G.M.E&R, Kol-20 and Director, KPCMC&H, Kol-32 for their kind support and help. Ananya Chatterjee is thankful to Board of Research in Nuclear Science (BRNS), Department of Atomic Energy (DAE), GOI India for providing fellowship. The work was financially supported by BRNS, DAE, GOI.
Author information
Authors and Affiliations
Corresponding author
Additional information
Highlights
1. Healing activity of water soluble polysaccharide-rich fraction of Termitomyces eurhizus (TEps) against the indomethacin induced gastric ulceration in mice model is proposed.2. Histological study shows that TEps (20 mg/kg) effectively healed the gastric ulceration.3. Reduction of MPO activity, protection of mucosal mucin content, enhanced synthesis of PGE2 by modulation of COX-1 and COX-2 expression and balance between pro inflammatory (TNF-α, IL-1ß) and anti inflammatory cytokines (IL-10) are responsible for ulcer healing.4. The preliminary study uncovers the anti-ulcerogenic property of TEps.
Appendix
Appendix
TEps, Polysaccharide-rich fraction of Termitomyces eurhizus; NSAID, Non Steroidal Anti Inflammatory Drug; PG, Prostaglandin; ELISA, Enzyme Linked Immunosorbent Assay; UI, Ulcer Index; TNF-a , Tumour Necrosis Factor , alpha ; IL , Interleukin; Omp, Omeprazole; g, Gram.
Rights and permissions
About this article
Cite this article
Chatterjee, A., Khatua, S., Chatterjee, S. et al. Polysaccharide-rich fraction of Termitomyces eurhizus accelerate healing of indomethacin induced gastric ulcer in mice. Glycoconj J 30, 759–768 (2013). https://doi.org/10.1007/s10719-013-9479-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10719-013-9479-5