
Abstract
Monounsaturated fatty acids (MUFA)-rich and n-6 polyunsaturated fatty acid (n-6 PUFA)-rich vegetable oils are increasingly used as fish oil replacers for aquafeed formulation. The present study investigated the fatty acid metabolism in juvenile European sea bass (Dicentrarchus labrax, 38.4 g) fed diets containing fish oil (FO, as the control treatment) or two different vegetable oils (the MUFA-rich canola/rapeseed oil, CO; and the n-6 PUFA-rich cottonseed oil, CSO) tested individually or as a 50/50 blend (CO/CSO). The whole-body fatty acid balance method was used to deduce the apparent in vivo fatty acid metabolism. No effect on growth performance and feed utilization was recorded. However, it should be noted that the fish meal content of the experimental diets was relatively high, and thus the requirement for n-3 long-chain polyunsaturated fatty acid (n-3 LC-PUFA) may have likely been fulfilled even if dietary fish oil was fully replaced by vegetable oils. Overall, relatively little apparent in vivo fatty acid bioconversion was recorded, whilst the apparent in vivo β-oxidation of dietary fatty acid was largely affected by the dietary lipid source, with higher rate of β-oxidation for those fatty acids which were provided in dietary surplus. The deposition of 20:5n-3 and 22:6n-3, as % of the dietary intake, was greatest for the fish fed on the CSO diet. It has been shown that European sea bass seems to be able to efficiently use n-6 PUFA for energy substrate, and this may help in minimizing the β-oxidation of the health benefiting n-3 LC-PUFA and thus increase their deposition into fish tissues.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Lipids are the main energy source for cultured finfish species, and traditionally, fish oil has been the principal lipid source used in aquafeed formulation. However, environmental and economic concerns over the use of fish oil are driving the aquafeed industry to increasing the substitution of this commodity with alternative, more economically advantageous and environmentally sustainable oil sources (Turchini et al. 2009). Different vegetable oils are characterized by a wide range of fatty acid compositions, but their common denominator is that they are all devoid of the n-3 long-chain polyunsaturated fatty acids (n-3 LC-PUFA), which, on the other hand, are abundant in and characteristic of fish oil (Sargent et al. 2002).
The fillet fatty acid composition of cultured fish fed vegetable oil-based diets is normally characterized by significantly decreased levels of n-3 LC-PUFA, compared to fish fed a fish oil-based diet. This situation is even more exacerbated in carnivorous marine finfish because, unlike the majority of freshwater species, marine finfish have been generally reported to have a limited ability to biosynthesize LC-PUFA from C18 PUFA precursors due to apparent limited fatty acid elongase and desaturase enzymatic activities (Mourente and Dick 2002; Mourente et al. 2005).
Vegetable oils can be classified into four major groups (Turchini et al. 2011): (1) those rich in saturated fatty acids (SFA; i.e. palm and coconut oil), (2) those rich in monounsaturated fatty acids (MUFA; such as rapeseed/canola and olive oil), (3) those rich in PUFA of the omega-6 series (n-6 PUFA; primarily linoleic acid, LA, 18:2n-6; such as soybean and cottonseed oil), and (4) those rich in PUFA of the omega-3 series (n-3 PUFA; primarily α-linolenic acid, ALA, 18:3n-3; such as linseed/flaxseed and camelina oil). Within the oils available for utilization in aquafeed formulation, those rich in MUFA or n-6 PUFA are readily available, in growing supply, favourably priced and they provide easily digestible energy for fish growth the most abundant; however, specific and direct comparisons of these two types of oils are scarce.
Most of the readily available vegetable oils have been tested across a variety of commercially relevant cultured species, and a considerable amount of knowledge has been generated related to the resultant modification of their final fatty acid make-up (Rosenlund et al. 2010) and their lipid and fatty acid metabolism (Tocher 2003; Torstensen and Tocher 2010). It has been shown that in European sea bass (Dicentrarchus labrax), one of the most important marine carnivorous species cultivated in Europe; it is possible to substitute levels up to 50–60 % of dietary fish oil with a range of vegetable oils, without negatively affecting growth performance and overall flesh quality (Izquierdo et al. 2003; Montero et al. 2005). In terms of n-3 LC-PUFA biosynthetic capabilities, Mourente et al. (2005) suggested that the bioconversion rate of radiolabeled α-linolenic acid (14C-ALA) to eicosapentaenoic acid (20:5n-3) and docosahexaenoic acid (22:6n-3) was present but quantitatively very limited in hepatocytes of European sea bass fed diet containing partial substitution of dietary fish oil with blends of rapeseed, linseed and palm oil. Accordingly, in European sea bass, hepatocytes in vitro production of the products of all the enzymes involved in the elongation and desaturation of C18 PUFA have been recorded (Mourente and Dick 2002), and similarly, in vivo production of n-6 LC-PUFA intermediates was noted in European sea bass fed a soybean oil-based diet (Izquierdo et al. 2003). More recently, the enzyme Δ-6 desaturase has been cloned and fully characterized in European sea bass (Santigosa et al. 2011).
Thus, it is evident that European sea bass possesses the so-called typical ‘marine fish pattern’ in the metabolism of 18:3n-3 to 20:5n-3, and therefore, it requires relatively high dietary levels of n-3 LC-PUFA (i.e. fish oil) in its diet (Mourente and Dick 2002; Mourente et al. 2005). Nevertheless, there is limited information on how dietary fatty acids, and in particular dietary MUFA and n-6 PUFA, which are commonly provided by vegetable oils, may affect the overall fatty acid metabolism, including fatty acid β-oxidation for energy production, fatty acid bioconversion and their deposition in the fillet of European sea bass.
The aim of the present study was to investigate the fatty acid metabolism in European sea bass fed 100 % fish oil replaced diets containing different level of MUFA and n-6 PUFA, through the monitoring of the metabolic fate of individual dietary fatty acids. To achieve these objectives, a feeding trial was conducted in which fish were fed on specifically formulated experimental diets containing alternative lipid sources with distinct fatty acid profiles. Specifically, two plant-derived oils were selected for their ‘extreme’ fatty acid composition: canola/rapeseed (CO), rich in MUFA, and cottonseed (CSO), rich in n-6 PUFA; and the experimental diets were composed of a control fish oil-based diet (FO) and three fish oil-deprived diets in which all the added lipid source were comprised of CO, CSO or a 50/50 blend of these two oils (CO/CSO).
Materials and methods
Animal, husbandry and experimental diets
Juvenile European sea bass, Dicentrarchus labrax, were stocked into a marine flow-through system consisting of 12 cylindro-conical tanks (500 L), each stocked with 20 fish (38.4 ± 0.2 g), and the 4 experimental treatments were tested in triplicate tanks/groups randomly allocated. The experimental rearing system was supplied with continuously aerated sea water (40 g L−1) at a flow rate of approximately 2 L min−1. Each tank was supplied with individual fluorescent light set-up to provide 12:12-h light/dark (0700–1900 hours) photoperiod. Fish were fed manually to apparent satiation twice daily (0900 and 1800 hours) for 162 days. The experiment was conducted at 27.6 ± 0.6 °C, dissolved oxygen and pH were measured every 5 days, and they averaged to 7.2 ± 0.4 mg L−1 and 7.8 ± 0.2 mg L−1, respectively.
Four iso-nitrogenous and iso-energetic fish meal-based experimental diets were formulated with 20 % lipid originating either from fish oil (Engraulis encrasicolus oil) (FO; as control diet), from CSO, CO or a blend of CSO and CO (CO/CSO; 1:1, w:w) (Table 1). Chromic oxide (Cr2O3) was included in the diets (1 %) as an internal marker for digestibility estimations. All diets contained 510 mg/g of fish meal as the main protein source, thus all fish oil-deprived diets contained some n-3 LC-PUFA derived from the residual lipid fraction of fish meal. The diets were prepared by thoroughly mixing all dry ingredients with a mixer and adding the wet ingredients to form a dough. The dough was screw-pressed though a 3 mm die and the strands of feed were air dried at room temperature to approximately 890 g kg−1 dry matter and stored at −20 °C throughout the duration of the feeding trial.
Sample collection
At the beginning of the experiment, an initial sample of 20 fish from the common pool of fish was culled for whole-body proximate and fatty acid analyses. Fish were starved for 48 h before sampling. All fish in each group were anaesthetized (2-phenoxyethanol at 0.5 mL L−1) (Sigma-Aldrich AS, MO, USA) and then weighed individually, after removal of excess surface moisture, to the nearest 0.01 g. At the termination of the experiment, all fish were individually weighed. Ten fish from each tank were carefully captured with a net, killed with a blow to the head (cerebral percussion) and stored at −20 °C for subsequent analyses. All fish handling procedures complied with Turkish guidelines for animal care (No. 28141) set by the Ministry of Food, Agriculture and Livestock.
Proximate and fatty acid analysis
Before analyses, samples of fish whole bodies were individually finely chopped and accurately homogenized. Proximate composition and fatty acid analysis of fish whole bodies, diets and faeces were performed using standard methods previously described (Turchini et al. 2003; Eroldoğan et al. 2004). Briefly, the total lipid content was analysed by chloroform/methanol extraction (2 : 1 v/v) (Folch et al. 1957). Fatty acid methyl esters (FAME) were prepared according to Metcalfe and Schmitz (1961) and analysed as described previously by Czesny and Dabrowski (1998), with only minor modifications. Prior to transmethylation, a known amount of nonadecanoate (19:0), proportional to the amount of total lipids detected (8 mg per 50 mg of lipids), was added to the samples as the internal standard for the subsequent quantification of fatty acids. FAME were then separated by gas chromatography (Agilent 6890 N, Santa Clara, USA), equipped with a flame ionization detector and fitted with a DB 23 (Agilent 6890 N, Santa Clara, USA) capillary column (60 m, 0.25 mm i.d. and 0.25 μm, Cat No: 112-88A7); the oven temperature programme was 190 °C for 35 min then increasing at 30 °C per min up to 220 °C where it was maintained for 5 min. Carrier gas was hydrogen (2 mL min−1 and split ratio was 30:1). Fatty acids were then identified by comparing their retention times to that of a standard mix of fatty acids (Sigma, Germany) and quantified by comparing their peak area with that of the internal standard.
Fatty acid digestibility and metabolism assessment
Faecal samples were collected during the last 22 days of the feeding trial. Prior to collection, a specific faecal collection columns were fitted to each of the experimental tanks by using the Guelph system (Cho et al. 1982), which was modified as described by Tibbetts et al. (2006). Faecal samples were collected on a daily basis. After the final meal (1800 hours), tanks were siphoned to remove the uneaten feed and the faecal collection columns set on each tank were covered with ice in a plastic bag to minimize possible bacterial growth. Faeces were then collected overnight up to the next morning (0700 hours) and were stored at −20 °C until analyses. Approximately, 12–13 h elapsed between the last meal and the faecal collection. The diets and freeze-dried faecal samples (35 g wet faecal each tank) were analysed for chromic oxide (Cr2O3) content according to Furukawa and Tsukahara (1966). The apparent digestibility coefficients of fatty acids were calculated from standard formula (Maynard et al. 1979). The computation of the whole-body fatty acid balance was carried out as initially described by Turchini et al. (2007) and with further developments as described in Turchini et al. (2008) and Turchini and Francis (2009) to assess the fish fatty acid metabolism. The results of the method have been reported as apparent in vivo enzyme activity (nmol g−1 day−1) and, for key fatty acids, their utilization (β-oxidation, bioconversion and accumulation) was also expressed and reported as percentage of their specific net intake.
Statistical analyses
Data were expressed as mean ± SEM. (5 fish pooled per tank, 3 tanks per treatment; n = 3; N = 12). Data were analysed by one-way analysis of variance (ANOVA) at a significance level of 0.05 %, after confirmation of normality and homogeneity of variance. Where significant differences were detected, data were subjected to a Student–Newman–Keuls post hoc test for identifying homogeneous subsets. All statistical analyses were performed using SPSS 11.5 (SPSS Inc., Chicago, IL, USA).
Results
The four experimental diets were iso-nitrogenous (varying from 458 to 469 mg g−1), iso-lipidic (varying from 192 to 204 mg g−1) and iso-energetic (varying from 19.5 to 20.0 kJ g−1) (Table 1). Mortality was not affected by dietary treatments, and the maximum mortality recorded was 1 fish per tank. At the end of the feeding trial, final gain was more than 2.6-fold higher than the initial weigh, and growth performance and feed efficiency parameters were unaffected by dietary treatments (Table 1).
The main difference in the experimental diets composition was their fatty acid profile, which varied significantly amongst the four diets (Table 2). Briefly, CO contained 428.6 mg/g lipid of 18:1n-9, CSO contained 407.2 mg/g lipid of 18:2n-6 and CO/CSO contained 307.3 and 288.9 mg/g lipid of 18:1n-9 and 18:2n-6, respectively. The three vegetable oil-based diets contained very limited amounts of n-3 LC-PUFA, varying from 14.0 to 19.5 mg/g lipid, whilst FO contained 136.2 mg/g lipid of these fatty acids. The fish whole-body fatty acid composition was markedly modified by the dietary treatment, with the main differences observed between diets being mirrored in the fish body (Table 2). In general, the three more abundant fatty acids found in European sea bass whole bodies, independently from the dietary treatments, were 16:0, 18:1n-9 and 18:2n-6. The major differences in the fatty acid composition of the fish whole body were as follows: (1) significantly higher 20:5n-3 (37.3 mg/g lipid), 22:6n-3 (83.7 mg/g lipid) and total n-3 LC-PUFA (140.2 mg/g lipid) in FO fed fish; (2) significantly higher 18:1n-9 (277.2 mg/g lipid) and total MUFA (353.1 mg/g lipid) in CO fed fish; and (3) significantly higher 18:2n-6 (228.0 mg/g lipid) and total n-6 PUFA (244.0 mg/g lipid) in CSO fed fish. The fatty acid composition of whole body of fish fed CO/CSO was somewhat midway between that recorded for fish fed CO and fish fed CSO, with relatively abundant 18:1n-9 (232.4 mg/g lipid) and 18:2n-6 (181.4 mg/g lipid).
The mean apparent fatty acids digestibility for each of the experiment diets are reported in Table 3. Individual fatty acids all recorded relatively high levels of digestibility values, despite a few statistical significant differences. The digestibility of saturated fatty acids tend to decrease with the increase of the carbon chain length (14:0 > 16:0 > 18:0 > 20:0). Irrespective of dietary treatment, the degree of unsaturation of individual fatty acids resulted in the digestibility of PUFA > MUFA > SFA. The lowest digestibility in 18:3n-3 was recorded in CSO fed fish, whilst the same fatty acid recorded no significant differences in its digestibility across other test diets. Digestibility of 18:2n-6 in fish fed the FO diet was significantly lower in comparison with the other test diets. Total PUFA had the highest digestibility for fish fed the FO and CO/CSO diets and the lowest for those fed the CO diet. n-3 PUFA were highly digested in FO fed fish, whilst the highest n-6 PUFA digestion was found to be in CO/CSO fed fish.
The fatty acid content of fish whole body (expressed as mg of fatty acid per fish, at the beginning and the end of the trial) and the total net intake of fatty acids (fatty acid content of the diet × feed intake × fatty acid digestibility; total mg of fatty acid absorbed per fish during the feeding trial) were computed. These data were then used for the computation of the individual fatty acids appearance or disappearance (mg of fatty acid appeared or disappeared per fish over the entire experimental period), which were greatly affected by the dietary treatment (Table 4). In general, with the exception of few instances such as 22:1n-11, 22:2n-6, 22:2n-6 and 20:4n-3 in CSO fed fish and 22:5n-3 in CO/CSO fed fish, all other fatty acids recorded a disappearance. Across all treatments, the fatty acids recording the highest disappearance were 16:0, 18:1n-9 and 18:2n-6, which were the most abundantly provided with the diets. In FO fed fish, remarkably higher disappearances of 20:5n-3 and 22:6n-3 (1,019.1 and 1,321.4 mg per fish, respectively) were recorded, compared to other treatments. Fish on CO treatment recorded the highest disappearance of 18:1n-9 (9,332.2 mg per fish), CSO fed fish recorded the highest disappearance of 18:2n-6 (8,942.7 mg per fish) and CO/CSO fed fish recorded disappearances of these two latter fatty acids (6,198.1 mg of 18:1n-9 and 5,922.5 mg of 18:2n-6 per fish) which were intermediate between that of CO and CSO (Table 4).
The apparent in vivo fatty acid β-oxidation, bioconversion (desaturation and/or elongation) expressed as nmol g−1 day−1 for major fatty acid classes are reported in Table 5. Overall, no apparent Δ-5 and Δ-9 desaturase activity were recorded, whilst Δ-6 desaturase activity was recorded only on 18:2n-6 in fish fed CO diet. Apparent in vivo elongation of all fatty acids (SFA, MUFA, n-6 PUFA and n-3 PUFA) was recorded in fish fed the CSO diet, whilst only n-6 PUFA apparent in vivo elongation was recorded in fish fed the CO/CSO diet. The total fatty acid apparent in vivo fatty acid β-oxidation was not affected by dietary treatments, with recorded values ranging from 5,903.3 to 6,212.7 nmol g−1 day−1. The highest apparent in vivo β-oxidation of n-3 PUFA was recorded in fish fed the FO diet, whilst the lowest value was observed in fish fed the CSO diet. FO, as with CO, recorded the highest apparent in vivo β-oxidation of SFA and MUFA, whilst the highest apparent in vivo β-oxidation of n-6 PUFA was recorded in fish fed the CSO diet (Table 5).
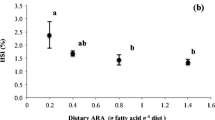
The level of apparent in vivo elongation, β-oxidation and deposition on some specific fatty acids, expressed as percentage of the specific fatty acid net intake, is reported in Table 6. A small percentage of dietary 18:2n-6 and 20:4n-6 was bioconverted to longer or more unsaturated fatty acids (0.09 and 0.82 % for fish fed the CSO diet, respectively), whilst larger percentages for 16:0, 18:1n-9, 18:2n-6 and 18:3n-3 were β-oxidized in the same group. Despite CO diet contained the highest amount of 18:3n-3, fish fed this diet recorded significantly higher apparent in vivo deposition of 18:3n-3 as percentage of net intake. Irrespective of the dietary treatment, the β-oxidation of 18:3n-3 was substantially high, ranging from 95.5 to 98.1 %. Significantly, higher deposition of 20:4n-6 was recorded in fish fed the CO (28.1 %) and CSO (30.5 %) diets, compared to the other two treatments (Table 6). The deposition of 20:5n-3 was significantly higher in fish fed the CSO diet (9.9 %), whilst 22:6n-3 was deposited at significantly higher rates in fish fed CSO (27.4 %) and CO (17.4 %) diets, compared to the other treatments.
Discussion
In this study, diets with markedly different fatty acid composition did not affect growth performance, survival and other feed utilization parameters (i.e. feed intake and food conversion ratio) in European sea bass, even compared to the fish fed a diet containing 100 % fish oil. In the previous study, Izquierdo et al. (2003) reported that replacement of 60 % fish oil with soybean oil, linseed oil, rapeseed oil and a blend of them did not negatively impact fish performance, when high lipid content (25 %) diets were tested. Accordingly, the lipid content of the experimental diets in the present study was relatively high (20 %), and notably higher in comparison with other experimental diets (11 %) previously used for testing potential alternative oils (sunflower and soybean oils) in European sea bass (Yıldız and Şener 1997). Other previous studies on the same species reported growth retardation in European sea bass when dietary fish oil was replaced with different vegetable oils (Izquierdo et al. 2003; Yıldız and Şener 2004; Mourente et al. 2005; Mourente and Bell 2006). Importantly, in the present study, diets contained a relatively large amount of fish meal (510 g kg−1), and thus also all experimental diets devoid of fish oil contained some n-3 LC-PUFA (varying from 0.28 to 0.38 % of dry diet in CSO and CO, respectively), which could help explain the lack of any major detrimental effect on fish performances. The known quantitative essential fatty acid (n-3 LC-PUFA) requirements for juvenile marine species vary from 0.3 to 2.5 % of dry diet (NRC 2011), and for European sea bass, the suggested minimal requirement of n-3 LC-PUFA is 1 % of dry diet (Coutteau et al. 1996). However, this value was recorded for weaning, first feeding, juveniles, and was also equivalent to the lowest n-3 LC-PUFA inclusion level tested in the latter study. Thus, the results of current study suggest that for larger juveniles (40–100 g), a dietary content of n-3 LC-PUFA as low as 0.28 % of dry diet can be sufficient to guarantee that minimal requirements are fulfilled.
The fatty acid composition of the whole body of European sea bass fed the four experimental diets was mirroring that of the diets. This observation is in full agreement with previous results on a variety of freshwater and some marine species, including European sea bass (Izquierdo et al. 2003; Bell et al. 2003; Mourente et al. 2005; Francis et al. 2007a). Despite the abundant provision of 18:2n-6 in all three diets in which fish oil was replaced, the final content of 20:4n-6 was lower than that recorded in fish on the FO diet, suggesting that the possible in vivo fatty acid bioconversion, if any, had only minimal to effect on final tissue fatty acid composition. Izquierdo et al. (2003) showed that the inclusion of dietary vegetable oils for gilthead sea bream and European sea bass led to a diet dependent reduction of 20:5n-3 and 20:4n-6 in muscle, whereas 22:6n-3 was deposited in fish tissue. Similar trends could be reported in the current study when observing the whole-body fatty acid composition of fish fed the three diets in which fish oil was replaced compared to the control group fed the FO diet. However, observing the results achieved by the whole-body fatty acid balance method, it was possible to note that the deposition of 20:4n-6 and 22:6n-3 was significantly higher in the three treatments in which fish oil was replaced compared to FO fed fish, whilst 20:5n-3 deposition was improved only in fish fed the CSO diet. Overall, the remarkable higher deposition rates recorded for LC-PUFA compared to other fatty acids are also in agreement with previous results on European sea bass (Mourente and Bell 2006) and other species, such as Atlantic salmon (Salmo salar) (Bell et al. 2001, 2003; Mourente and Bell 2006) and brown trout (Arslan et al. 2012). This apparent selective deposition might be due to the physiological importance of these fatty acids (Sargent et al. 2002; Bell and Sargent 2003). Interestingly, when observing the recorded whole-body fatty acid composition, some of values recorded for CO/CSO were not in between the values reported for CO and CSO treatments, as one could expect. For example, whilst the values of 18:2n-6 and 18:3n-3 recorded in CO/CSO whole bodies were in between the values observed for fish on CO and CSO treatments, the content of 20:4n-6, 20:5n-3 and 22:6n-3 were in all instances lower for CO/CSO compared to the other two treatments (CO and CSO). A possible explanation for these results will be attempted below when discussing fatty acid β-oxidation and deposition.
The individual fatty acid digestibility of the experimental diets increased with increasing degree of unsaturation and decreased with chain length as reported for various fish species (Morais et al. 2005; Turchini et al. 2005; Francis et al. 2007b). At this point, it seems important to remember that oleic acid-rich oils, such as canola or olive oil, are apparently less digested than n-6 PUFA-rich diets (Brown and Hart 2010), similar to that observed in the present study. Olsen et al. (1998) reported that the digestive lipase specificity towards PUFA partly explains better digestion of PUFA than MUFA or SFA. The fatty acids present in the highest concentrations in the experimental diets were mainly 18:1n-9 (CO, MUFA-rich oil diet) and 18:2n-6 (CSO, n-6 PUFA-rich oil diet), which have individual melting points of 13.0 and −5 °C, respectively. Thus, the high melting points of 18:1n-9 compare to 18:2n-6 can explain the decreased overall apparent fatty acid absorption of LC-PUFA (n-3 LC-PUFA and n-6 LC-PUFA) in fish fed the CO diet compared to other treatments.
Previous studies on the fatty acid bioconversion capabilities of fish have been primarily done on salmonid species, and most commonly using ex vivo methods providing a series of important and fundamental information (Bell et al. 1997; Tocher et al. 2002; Stubhaug et al. 2005; Zheng et al. 2005; Mourente et al. 2005). The intrinsic advantages and/or limitation of ex vivo and in vivo methods have been critically reviewed by Brown (2005), and accordingly, more recently, the whole-body fatty acid balance method has been increasingly used in animal nutrition studies as a valuable additional analytical tool capable of providing a general overview of the apparent in vivo fatty acid deposition/retention, elongation, desaturation and β-oxidation (Cunnane and Anderson 1997; Turchini et al. 2007). This method offers a series of advantages, as it is simply implemented; it can quite accurately describe the overall apparent in vivo enzymatic activities and has been proved useful to provide innovative insights into in vivo fatty acid metabolism (Thanuthong et al. 2011; Cleveland et al. 2012), but has also some limitations. Specifically, one of the most commonly reported drawbacks of this method is the relatively large variability recorded (Senadheera et al. 2012). Secondly, and importantly, the results obtained by this method cannot be directly compared to results obtained by in vitro methodologies and should always be reported and considered as ‘apparent in vivo enzymes activities’ (Turchini et al. 2007), to avoid possible confusion in the interpretation of results.
To the best of the authors’ knowledge, this is the first instance in which this method has been implemented in European sea bass. The principal observation reported in the present study is that European sea bass did not elongate and/or desaturate dietary 18:3n-3 and 18:2n-6, and this is consistent with previous studies on the same species. In fact, Mourente et al. (2005) and Mourente and Dick (2002) demonstrated that the European sea bass presented a ‘marine fish pattern’ in the metabolism of 18:3n-3 to 20:5n-3 and 22:6n-3. Nevertheless, a Δ-6 desaturase activity on n-6 PUFA, though quantitatively very limited, was recorded in the present study, and this is consistent with previous indication that European sea bass posses this metabolic capability (Mourente and Dick 2002; Izquierdo et al. 2003; Mourente et al. 2005; Turchini et al. 2009; Santigosa et al. 2011). On the other hand, it seems that fatty acid elongase activity is more prominent in European sea bass, and consistently with previous studies on this species (Mourente et al. 2005), a very low direct elongation towards ‘dead end’ products of 18:3n-3 and 18:2n-6 also occurred in fish fed the three diets deprived of fish oil in the current study. Importantly, the relatively high content of fish meal in the present experimental diets, and thus the presence of some n-3 LC-PUFA derived from the residual fat of the fish meal fraction, may have limited the fatty acid bioconversion. Nevertheless, it seems likely that the LC-PUFA biosynthesis in European sea bass is theoretically possible, but extremely limited and in practice nil, as previously suggested (Mourente and Dick 2002; Mourente et al. 2005; González-Rovira et al. 2009; Turchini et al. 2009; Santigosa et al. 2011). In agreement with these observations recently, Alhazzaa et al. (2011) also found a limited efficiency for LC-PUFA biosynthesis in Asian sea bass (Lates calcarifer; an euryhaline carnivore which can be considered similar to European sea bass) receiving vegetable oils containing diets. As the life cycle of European and Asian sea basses involves different saline environments (i.e. freshwater to coastal waters), their digestive systems appear to be adapted to utilize nutrients over a wide range of salinities that will have other critical roles in other physiological aspects (Eroldoğan et al. 2004; Harpaz et al. 2005; Alhazzaa et al. 2011). In general, LC-PUFA biosynthesis in some freshwater fish is greater than marine fish due to high concentration of 18:3n-3 and limited 22:6n-3 in their natural diet (Sargent et al. 2002). However, and interestingly, Alhazzaa et al. (2011) found that salinity did not appear to influence the PUFA bioconversion activity in juvenile Asian sea bass.
In general, in the present study, it was shown that the fatty acid bioconversion activities in European sea bass were very limited and mainly related to the SFA, MUFA and n-6 PUFA, and no apparent in vivo fatty acid Δ-5 nor Δ-6 desaturase activity on n-3 PUFA was recorded. This suggests that any differences noted in the n-3 LC-PUFA content of fish tissues should be related to the actual deposition efficiency and/or β-oxidation of n-3 LC-PUFA (Turchini and Francis 2009; Turchini et al. 2009). Lower β-oxidation of n-3 LC-PUFA, and therefore higher deposition into fish tissues, was recorded in Murray cod (Maccullochella peelii peelii) fed with abundant MUFA and to a lesser extent SFA (Turchini et al. 2011). Conversely, in the present study on European sea bass, it seemed that higher dietary content of 18:2n-6 (CSO treatment) resulted in lower β-oxidation and higher deposition/retention of n-3 LC-PUFA. This observation clearly warrants further investigations, as it seems that the different species can have different preferential β-oxidation for the two different fatty acid classes (MUFA and n-6 PUFA, respectively) over n-3 LC-PUFA.
In salmonids, a marginal increases in the β-oxidation capacity in fish fed high n-3 LC-PUFA compared to those fed n-3 LC-PUFA deficient diets have been reported (Stubhaug et al. 2005, 2006, 2007). Moreover, it was suggested that this was primarily due to an increased uptake of fatty acid into the cells or into mitochondria, rather than a direct stimulation of the actual β-oxidation system (Turchini et al. 2003; Torstensen and Stubhaug 2004). Previous studies on Atlantic salmon and rainbow trout (Oncorhynchus mykiss) have indicated that 18:3n-3 and 18:2n-6, as well as MUFA (namely 18:1n-9 and 22:1n-11), are readily β-oxidized when present in high concentrations in the diets (Torstensen et al. 2004; Stubhaug et al. 2007; Turchini and Francis 2009). Accordingly, in the present study, the apparent in vivo β-oxidation of dietary 18:1n-9 and 18:2n-6 was highest in fish receiving the MUFA-rich CO diet and the n-6 PUFA-rich CSO diet, respectively, clearly demonstrated that dietary fatty acids are β-oxidized proportionally to their availability.
As mentioned above with regard to the whole-body fatty acid composition, similarly, some of the apparent in vivo β-oxidation and deposition values recorded for CO/CSO were not in between the values recorded for CO and CSO treatments. Specifically, the deposition values recorded for 20:4n-6, 20:5n-3 and 22:6n-3 were in all instances lower for CO/CSO compared to the other two treatments (CO and CSO). Thus, it seems plausible to speculate that the provision of a single fatty acid in large abundant surplus, such as 18:1n-9 in CO or 18:2n-6 in CSO, may be more efficient in sparing LC-PUFA from β-oxidation, and thus improve their deposition, rather than the provision of a diet containing a more balanced blend of fatty acids. This explanation, though only speculative at this stage, would be in full agreement with the previous observations that dietary fatty acids are preferentially and abundantly β-oxidized when provided in surplus (Stubhaug et al. 2005, 2006, 2007).
In conclusion, this study provided additional evidences to confirm the hypothesis that even though European sea bass may possess all the enzymatic machinery to biosynthesize LC-PUFA, it is very inefficient, and thus an appropriate amount of LC-PUFA has to be provided with the diet. Thus, the possible optimization of European sea bass diets should focus on strategies towards improving the efficiency of LC-PUFA retention and deposition. Accordingly, a novel finding of this study was that European sea bass seems to be able to efficiently use n-6 PUFA for energy substrate, and this can help in minimizing the β-oxidation of n-3 LC-PUFA, and thus increasing their deposition into body compartments. However, the provision of high amount of dietary n-6 PUFA can also be considered as having a negative effect on the overall nutritional characteristic of European sea bass fillets for human consumption, and thus other strategies towards sparing dietary n-3 LC-PUFA from oxidation, warrant further investigations.
References
Alhazzaa R, Bridle AR, Nichols PD, Carter CG (2011) Replacing dietary fish oil with echium oil enriched barramundi with C18 PUFA rather than long-chain PUFA. Aquaculture 312:162–171
Arslan M, Sirkecioglu AN, Bayir A, Arslan H, Aras M (2012) The influence of substitution of dietary fish oil with different vegetable oils on performance and fatty acid composition of brown trout, Salmo trutta. Turkish J Fish Aqua Scien 12:575–583
Bell JG, Sargent JR (2003) Arachidonic acid in aquaculture feeds: current status and future opportunities. Aquaculture 128:491–499
Bell JG, Tocher DR, Farndale BM, Cox DI, McKinney RW, Sargent JR (1997) The effect of dietary lipid on polyunsaturated fatty acid metabolism in Atlantic salmon (Salmo salar) undergoing parr-smolt transformation. Lipids 32:515–525
Bell JG, McEvoy J, Tocher DR, McGhee F, Campbell PJ, Sargent JR (2001) Replacement of fish oil with rapeseed oil in diets of Atlantic salmon (Salmo salar) affects tissue lipid compositions and hepatocyte fatty acid metabolism. The Journal of nutrition 131:1535–1543
Bell JG, Tocher DR, Henderson RJ, Dick JR, Crampton VO (2003) Altered fatty acid compositions in Atlantic salmon (Salmo salar) fed diets containing linseed and rapeseed oils can be partially restored by a subsequent fish oil finishing diet. The Journal of nutrition 133:2793–2801
Brown JE (2005) A critical review of methods used to estimate linoleic acid Δ6-desaturation ex vivo and in vivo. Eur J Lipid Sci Technol 107(2):119–134
Brown PB, Hart SD (2010) Soybean oil and other n-6 polyunsaturated fatty acid-rich vegetable oil. In: Turchini GM, Ng WK, Tocher DR (eds) Fish oil replacement and alternative lipid sources in aquaculture feeds. CRC Press, Boca Raton, pp 133–160
Cho CY, Slinger SJ, Bayley HS (1982) Bioenergetics of salmonid fishes: energy intake, expenditure and productivity. Comp Biochem Physiol Part B 73:25–41
Cleveland BJ, Francis DS, Turchini GM (2012) Echium oil provides no benefit over linseed oil for (n-3) long-chain PUFA biosynthesis in rainbow trout. The Journal of Nutrition 142:1449–1455
Coutteau P, Van Stappen G, Sorgeloos P (1996) A standard experimental diet for the study of fatty acid requirements of weaning and first ongrowing stages of marine fish: a comparison of extruded and extruded/coated diets. Arch Anim Nutr 49:49–59
Cunnane SC, Anderson MJ (1997) The majority of dietary linoleate in growing rats is B-oxidized or stored in visceral fat. The Journal of nutrition 127(1):146–152
Czesny S, Dabrowski K (1998) The effect of egg fatty acid concentrations on embryo viability in wild and domesticated walleye (Stizostedion vitreum). Aquat Living Resour 11:371–378
Eroldoğan O, Kumlu M, Aktaş M (2004) Optimum feeding rates for European sea bass Dicentrarchus labrax L. reared in seawater and freshwater. Aquaculture 231:501–515
Folch J, Lees M, Sloane-Stanley G (1957) A simple method for the isolation and purification of total lipids from animal tissues. J Biol Chem 226:497–509
Francis DS, Turchini GM, Jones PL, De Silva SS (2007a) Dietary lipid source modulates in vivo fatty acid metabolism in the freshwater fish, Murray cod (Maccullochella peelii peelii). Journal of agricultural and food chemistry 55:1582–1591
Francis DS, Turchini GM, Jones PL, De Silva SS (2007b) Effects of fish oil substitution with a mix blend vegetable oil on nutrient digestibility in Murray cod, Maccullochella peelii peelii. Aquaculture 269:447–455
Furukawa A, Tsukahara H (1966) On the acid digestion method for the determination of chromic oxide as an indicator substance in the study of digestibility in fish. Bulletin of the Japanese Society for the Science of Fish 32:502–506
González-Rovira A, Mourente G, Zheng X, Tocher DR, Pendón C (2009) Molecular and functional characterization and expression analysis of a Δ6 fatty acyl desaturase cDNA of European Sea Bass (Dicentrarchus labrax L.). Aquaculture 298:90–100
Harpaz S, Hakim Y, Slosman T, Eroldoğan OT (2005) Effects of adding salt to the diet of Asian sea bass Lates calcarifer reared in fresh or salt water recirculating tanks, on growth and brush border enzyme activity. Aquaculture 248:315–324
Izquierdo MS, Obach A, Arantzamendi L, Montero D, Robaina L, Rosenlund G (2003) Dietary lipid sources for seabream and seabass: growth performance, tissue composition and flesh quality. Aquac Nutr 9:397–407
Maynard L, Loosli J, Hintz H, Warner R (1979) Animal nutrition. 7th. The carbohydrates and their metabolism. McGraw-Hill, Inc., New York, pp 74–103
Metcalfe L, Schmitz A (1961) The rapid preparation of fatty acid esters for gas chromatographic analysis. Anal Chem 33:363–364
Montero D, Robaina L, Caballero MJ, Gines R, Izquierdo MS (2005) Growth, feed utilization and flesh quality of European sea bass (Dicentrarchus labrax) fed diets containing vegetable oils: a time-course study on the effect of are-feeding period with a 100% fish oil diet. Aquaculture 248:121–134
Morais S, Koven W, Rønnestad I, Dinis MT, Conceição LEC (2005) Dietary protein:lipid ratio and lipid nature affects fatty acid absorption and metabolism in a teleost larva Brit. J Nutr 93:813–820
Mourente G, Bell J (2006) Partial replacement of dietary fish oil with blends of vegetable oils (rapeseed, linseed and palm oils) in diets for European sea bass (Dicentrarchus labrax L.) over a long term growth study: effects on muscle and liver fatty acid composition and effectiveness of a fish oil finishing diet. Comp Biochem Physiol Part B 145:389–399
Mourente G, Dick JR (2002) Influence of partial substitution of dietary fish oil by vegetable oils on the metabolism of 1-C-14 18: 3n–3 in isolated hepatocytes of European sea bass (Dicentrarchus labrax L.). Fish Physio Biochem 26:297–308
Mourente G, Dick JR, Bell JG, Tocher DR (2005) Effect of partial substitution of dietary fish oil by vegetable oils on desaturation and β-oxidation of [1-14-C] 18:3n–3 (LNA) and [1-14-C] 20: 5n–3 (EPA) in hepatocytes and enterocytes of European sea bass (Dicentrarchus labrax L.). Aquaculture 248:173–186
National Research Council (2011) Nutritional requirements of fish and shrimps. The National Academic Press, Washington, DC, p 376
Olsen RE, Henderson RJ, Ring E (1998) The digestion and selective absorption of dietary fatty acids in Arctic charr, Salvelinus alpinus. Aquac Nutr 4:13–21
Rosenlund G, Corraze G, Izquierdo MS, Torstensen BE (2010) The effects of fish oil replacement on nutrition and organoleptic qualities of farmed fish. In: Turchini GM, Ng WK, Tocher DR (eds) Fish oil replacement and alternative lipid sources in aquaculture feeds. CRC Press, Boca Raton, pp 487–552
Santigosa E, Geay F, Tonon T, Le Delliou H, Kuhl H, Reinhardt R, Corcos L, Cahu C, Zambonino-Infante JL, Mazurais D (2011) Cloning, tissue expression analysis, and functional characterization of two Δ6-desaturase variants of sea bass (Dicentrarchus labrax L.). Mar Biotechnol 13:22–31
Sargent JR, Tocher DR, Bell JG (2002) The lipids. In: Halver JE, Hardy RW (eds) Fish nutrition, 3rd edn. Academic Press, San Diego, pp 181–257
Senadheera SPSD, Turchini GM, Thanuthong T, Francis DS (2012) Effects of dietary iron supplementation on growth performance, fatty acid composition and fatty acid metabolism in rainbow trout (Oncorhynchus mykiss) fed vegetable oil based diets. Aquaculture 343:80–88
Stubhaug I, Frøyland L, Torstensen BE (2005) β-oxidation capacity of red and white muscle and liver in Atlantic salmon (Salmo salar L.)-effects of increasing dietary rapeseed oil and olive oil to replace capelin oil. Lipids 40:39–47
Stubhaug I, Lie Ø, Torstensen BE (2006) β-Oxidation capacity in liver increases during parr-smolt transformation of Atlantic salmon fed vegetable oil and fish oil. J Fish Biol 69:504–517
Stubhaug I, Lie Ø, Torstensen B (2007) Fatty acid productive value and β-oxidation capacity in Atlantic salmon (Salmo salar L.) fed on different lipid sources along the whole growth period. Aquac Nutr 13:145–155
Thanuthong T, Francis DS, Senadheera SPSD, Paul LJ, Turchini GM (2011) LC-PUFA biosynthesis in rainbow trout is substrate limited: use of the whole body fatty acid balance method and different 18:3n-3/18:2n-6 ratios. Lipids 46:1111–1127
Tibbetts SM, Milley JE, Lall SP (2006) Apparent protein and energy digestibility of common and alternative feed ingredients by Atlantic cod, Gadus morhua (Linnaeus, 1758). Aquaculture 261:1314–1327
Tocher DR (2003) Metabolism and functions of lipids and fatty acids in teleost fish. Rev Fish Sci 11:107–184
Tocher DR, Fonseca-Madrigal J, Bell JG, Dick JR, Henderson RJ, Sargent JR (2002) Effects of diets containing linseed oil on fatty acid desaturation and oxidation in hepatocytes and intestinal enterocytes in Atlantic salmon (Salmo salar). Fish Physio Biochem 26:157–170
Torstensen B, Stubhaug I (2004) β-Oxidation of 18:3n-3 in atlantic salmon (Salmo salar L.) hepatocytes treated with different fatty acids. Lipids 39:153–160
Torstensen BE, Tocher DR (2010) The effects of fish oil replacement on lipid metabolism of fish. In: Turchini GM, Ng WK, Tocher DR (eds) Fish oil replacement and alternative lipid sources in aquaculture feeds. CRC Press, Boca Raton, pp 487–522
Torstensen BE, Frøyland L, Lie Ø (2004) Replacing dietary fish oil with increasing levels of rapeseed oil and olive oil—effects on Atlantic salmon (Salmo salar L.) tissue and lipoprotein lipid composition and lipogenic enzyme activities. Aquac Nutr 10:175–192
Turchini GM, Francis DS (2009) Fatty acid metabolism (desaturation, elongation and b-oxidation) in rainbow trout fed fish oil-or linseed oil-based diets. Br J Nutr 102:69–81
Turchini GM, Gunasekera RM, De Silva SS (2003) Effect of crude oil extracts from trout offal as a replacement for fish oil in the diets of the Australian native fish Murray cod Maccullochella peelii peelii. Aquacult Res 34:697–708
Turchini GM, Caprino F, Giani I, Panseri S, Bellagama F, Moretti MV, Valfré F (2005) The relative absorption of fatty acids in brown trout (Salmo trutta) fed a commercial extruded pellet coated with different lipid sources. It J Anim Sci 4:241–252
Turchini GM, Francis DS, De Silva SS (2007) A whole body, in vivo, fatty acid balance method to quantify PUFA metabolism (desaturation, elongation and beta-oxidation). Lipids 42:1065–1071
Turchini G, Francis D, De Silva S (2008) A whole body, in vivo, fatty acid balance method to quantify PUFA metabolism (desaturation, elongation and beta-oxidation). Lipids 43:977
Turchini GM, Torstensen BE, Ng WK (2009) Fish oil replacement in finfish nutrition. Revi Aquac 1:10–57
Turchini GM, Francis DS, Senadheera SPSD, Thanuthong T, De Silva SS (2011) Fish oil replacement with different vegetable oils in Murray cod: evidence of an “omega-3 sparing effect” by other dietary fatty acids. Aquaculture 315:250–259
Yıldız M, Şener E (1997) Effect of dietary supplementation with soybean oil, sunflower oil or fish oil on the growth of seabass (Dicentrarchus labrax L. 1758). Cah Options Mediterr 225–233
Yıldız M, Şener E (2004) The effect of dietary oils of vegetable origin on the performance. Body composition and fatty acid profiles of sea bass (Dicentrarchus labrax L., 1758) juveniles. Turk J Vet Anim Sci 28:553–562
Zheng X, Torstensen BE, Tocher DR, Dick JR, Henderson RJ, Bell JG (2005) Environmental and dietary influences on highly unsaturated fatty acid biosynthesis and expression of fatty acyl desaturase and elongase genes in liver of Atlantic salmon (Salmo salar). BiochimBiophys Acta 1734:13–24
Acknowledgments
This study was supported by The Scientific & Technological Research Council of Turkey (TÜBİTAK) (project 106O195) and part of a PhD thesis of A.H.Y. who was granted by the Research Fund (SUF2011D1) of University of Çukurova. The authors also express their acknowledgments to Dr. Oğuz Taşbozan for his support in formulation of the diets.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Eroldoğan, T.O., Yılmaz, A.H., Turchini, G.M. et al. Fatty acid metabolism in European sea bass (Dicentrarchus labrax): effects of n-6 PUFA and MUFA in fish oil replaced diets. Fish Physiol Biochem 39, 941–955 (2013). https://doi.org/10.1007/s10695-012-9753-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10695-012-9753-7