
Abstract
Deeper plant root systems are desired for improved water and nitrogen uptake in leaching environments. However, phenotyping for deep roots requires methods that enable plants to develop deep roots under realistic conditions. Winter cereals raise further complications as early growth occurs under low light and temperature during autumn and winter—conditions not met in standard glasshouse facilities. This study used tube rhizotrons of 2 m length, positioned outdoor under a rainout shelter to screen for depth penetration rates (DPR) of roots. Rooting depths of 1 to 1.5 m were achieved with 23 widely grown North European winter wheat cultivars in two autumn/winter and two summer experiments and nine of the cultivars were represented in two or more experiments. Heritability of DPR of roots was only consistent in autumn/winter experiments (27 %) signifying the importance of phenotyping in relevant seasons and environments. Depth penetration rate of roots varied significantly within the tested cultivars, from 1.39 (±0.35) mm °C−1 day−1 for cv. Tuareg to 2.07 (±0.34) mm °C−1 day−1, for cv. Mercedes. This study documented consistent differences of DPR among North-European winter wheat cultivars in long tube rhizotrons under semi-natural conditions, which may form part of future phenotyping facilities for deep rooting traits.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Winter wheat is one of the most widely cultivated crops in temperate climates. It is sown during the autumn and develops slowly during the dark and cold period of the Northern European winter with frequently high precipitation. Leaching of nitrogen below the root zone of crops is frequent during winter time, hence it is a challenge to keep nitrogen within the root zone in this environment. Nitrogen leaching causes environmental problems in groundwater, lakes, fjords and other coastal areas. As a consequence strict regulations on nitrogen fertilization have been implemented in the European Union. The ideotype for improved resource capture from deeper soil layers have been suggested to include deep roots with reduced or increased root density at the surface depending on target environment (Mi et al. 2010; Wasson et al. 2012; Lynch 2013). Breeding for deeper root systems is an unexploited approach that has the potential to increase the NUE (Foulkes et al. 2009) as rooting depth has been shown to be the most important limiting factor for deep nitrogen uptake in the field (Kristensen and Thorup-Kristensen 2004a, b). Deeper rooted crops will allow for utilisation of some of the nitrogen that would otherwise be lost to the environment. It is further widely accepted that deep root systems increase yield under drought stress (Christopher et al. 2008; McKenzie et al. 2009; Wasson et al. 2012). The value of such deeper root systems has been further stressed by Hammer et al. (2009) who showed that historical increases in US maize yields with increasing plant densities can only be explained by steeper and deeper root systems in the newly released maize cultivars.
Development of deeper root systems therefore has two potential benefits: (1) Increased NUE and thereby decreased nitrogen loss to groundwater and ecosystems and (2) increased drought tolerance.
Many different root traits have been studied using a variety of root phenotyping methods and >650 QTLs for these traits have been identified in rice (Champoux et al. 1995; Courtois et al. 2009). Desired root traits linked to drought resistance have been bred into high yielding rice using marker assisted back crossing (Steele et al. 2006). Thus there are promising prospects for future breeding for a wide range of root traits, but breeding for rooting depth remains problematic.
One major reason that selection for deep rooting is currently not part of crop breeding efforts is the difficulties in phenotyping the trait. The environmental factors and their interacting effects have a large influence on the rooting depth in the field (Botwright and Wade 2012; Rich and Watt 2013). More efficient methods to indirectly assess deep root traits are of interest to both plant breeders and scientists, as studies directly in field soil are resource-demanding. Plants grown in hydroponics, growth pouches and transparent growth media such as gels have been widely applied to root studies. Such approaches can greatly reduce the workload and exclude the complicating factors of experiments in field soil, however, they are generally not successful in predicting rooting depths of mature plants in the field (Wasson et al. 2012; Rich and Watt 2013; Watt et al. 2013).
Though there is a need to phenotype deep root traits in soil, few such studies have been reported. In wheat, differences in deep root traits (rooting depth >1 m) has to the authors’ knowledge only been reported by Gregory et al. (2005), Ford et al. (2006), Botwright and Wade (2012) and Rich and Watt (2013). For these reasons it is still largely unknown if agronomically significant differences in deep root growth traits among currently cultivated wheat cultivars exist. This knowledge is of special interest because deep rooting traits of currently grown wheat cultivars may be exploited faster in breeding programs than genotypes with more exotic backgrounds. Furthermore, because they are agronomically adapted and comparable in environmental adaptions and yield potentials they are immediately valuable for further studies of the importance of deeper rooting for water and nutrient utilisation.
For root studies, large growth compartments with transparent interfaces in the shape of tube rhizotrons offer an intermediate between field experiments and experiments performed in hydroponics, growth pouches, gels and paper rolls. Therefore tube rhizotrons has been proposed as part of the most rewarding strategy for the future (Gregory et al. 2009). It has been shown previously for pea genotypes that such systems can be used to distinguish genotypic differences in deep root traits and that these differences correlate well with genotypic differences measured in the field (Thorup-Kristensen 1998). For relevant phenotyping of deep root traits, growth compartments of at least 1.5 m are needed as shorter tube rhizotrons will not capture variations in root architecture at depth.
Root growth patterns are affected both by plant phenology as well as temperature and light conditions (Kaspar and Bland 1992; Mozafar et al. 1993; Nagel et al. 2009). This offers a special challenge for phenotyping root trait of winter cereals, due to their need for vernalisation to initiate reproductive growth. Under natural conditions this happens simultaneously with low light intensity and short photoperiod. Experiments mimicking winter climate take several months and offer special challenges to the facilities whereas experiments mimicking summer climate are fast and pose less challenges to facilities. Therefore summer experiments are both the fastest and easiest alternative. It is, however, still unknown how such results from summer experiments relate to winter root growth as seasonal × genotype effects on root traits have not been clarified.
The objectives of this study were: (i) To evaluate the use of long tube rhizotrons as an approach for study of deep root traits (ii) to determine if differences exist among current available cultivars of winter wheat in depth penetration rate, (iii) to determine if such differences in root depth affect the uptake of nitrogen measured as total nitrogen in leaf and stem.
Materials and methods
Experimental series and design
Four experiments were conducted with winter wheat grown in tube rhizotrons. The first two experiments were used as an initial screen for cultivar differences in major root growth traits. In later experiments only the most contrasting cultivars were included. This was done to validate differences from the first experiments and study seasonal effects. Improvements to the experimental setup were continuously implemented based on prior experience. A total of 23 cultivars were initially included in the study, while a total of 9 cultivars were included in two or more experiments (Table 1). Two of the experiments took place during summer (Exp.1S and Exp.3S) and two in autumn/winter (Table 3). All experiments were conducted as complete randomized block designs with four blocks in each experiment and one replicate per block except for Exp.3S, which contained four replicates per block.
Experimental procedure and tube rhizotron preparation
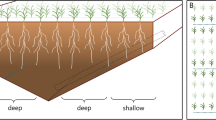
The experimental tube rhizotrons were made of transparent PVC tubes (OTV Plast A/S) filled with soil. The tubes were 2 m long, 74 mm in inner diameter 80 mm in outer diameter. The bottoms of the tubes were covered with gauze with mesh size of 1.4 mm. Tubes were fixed on frames each holding 20 tube rhizotrons (Fig. 1). Each tube rhizotron was seeded with three seeds. At leaf one stage the plants were thinned out and only one plant was left in each tube rhizotron, corresponding to a plant density of 232 plants m−2 soil surface area. The experiments were carried out under a rain out shelter in a field in Taastrup, Zealand, Denmark 55°40′90.35″N and 12°18′24.84E, 23 m above sea level. The rain-out shelter transmitted minimum 50 % of the photosynthetic active radiation depending on the height of the sun and amount of diffuse light.

Schematic drawing of frames containing 20 tube rhizotrons. The drawing does not show the doors of the box covering the frames. Doors were removed during root measurements
Winter wheat cultivars selected for the experiments were among the most popular cultivars presently grown in Denmark, Sweden, Germany, Great Britain, France and the Baltic countries. They were chosen to represent material from different breeders and with a broad range of parents. Information on selected cultivars is shown in Table 1. Breeders were consulted to ensure that the cultivars differed in their observed above-ground growth patterns.
The soil filled into the tubes was taken from a field at the experimental farm. Topsoil (surface 0–30 cm) and subsoil (30–60 cm) were collected separately and used to mimic the field soil profile. The subsoil was filled in the bottom of the tube rhizotrons and topsoil in the top except for Exp.1S where the whole profile consisted of topsoil. The topsoil was a loamy sand/sandy loam (9,6 % clay (<0,002 mm), 10 % silt (0,002–0,02 mm), 32,3 % fine sand (0,02–0,2 mm), 46,9 % coarse sand (0,2–2,0 mm), 2,5 % humus). The subsoil was a sandy soil (3,9 % clay (< 0,002 mm), 4,7 % silt (0,002–0,02 mm), 33,3 % fine sand (0,02–0,2 mm), 57,1 % coarse sand (0,2–2,0 mm), 1 % humus). The air dry soil was sieved on a 10 mm sieve. Soil information can be seen in Table 2.
In Exp.1S each tube received 12.5 kg topsoil. The soil was filled into the tubes stepwise at portions of approximately 0.5 kg. An internal steel piston was used to compact each portion soil by dropping it 30 cm 10 times per portion. In the following experiments subsoil was used in the lower parts of the tubes and soil was more uniformly compacted. This was achieved by adding soil loosely into the upright tubes while they were held on a vibrating plate powered by a NTK 25 AL piston oscillator (NTK® Oscillators) set at 6 bar and an amplitude of 5.8 mm. Soil was added to a set height of 1.75 m in Exp.2S and Exp.3W and 1.80 m in Exp.4W. Each tube rhizotron was vibrated for a total of 3 min. The weight of each tube rhizotron was checked subsequently. The differences in mean bulk density among tube rhizotrons did not exceed ±5 %, ±0.6 % and 2 % in Exp.2W, Exp.3S and Exp.4W respectively (Table 2).
In all experiments topsoil was added to a set height of 1.5 cm below the top of the tube. In Exp.1S this was done by adding 1 kg soil loosely to the last 10 cm of the tube and compact it by hand. In Exp.2W, Exp.3S and Exp.4W topsoil was likewise added loosely to the tubes and compacted by hand. In Exp.1S and Exp.3S the soil was air-dry at the time of filling followed by addition of water until field capacity was reached. In Exp.2W and Exp.4W water was mixed into the soil before filling the tubes. In Exp.2W this was done by using a shovel and in Exp.4W it was done by spraying water onto the dry soil while it was rotating in a mixer. This produced a gravimetric moisture content of 5.4 and 9.1 % in the subsoil and 5.4 and 9.0 % in the topsoil in Exp.2W and Exp.4W respectively.
Insulation and exclusion of light
In Exp.1S each tube rhizotron was covered by a plastic sheet, white on the outside and black on the inside. The 20 tube rhizotron on each frame were then further wrapped within a curtain of the same plastic product. In later experiments this approach was improved. Each frame was insulated on all sides and on the top by a “box” made of RIALET® Foamalux 10 mm plates (Brett Martin Ltd. UK), a white foamed PVC product. Holes were made in the top for the tube rhizotron to go through the insolating layer. The soil surface and above ground parts of the plant were exposed to the environment while the rest of the tube rhizotron was enclosed within the box. In Exp.2W the individual tube rhizotron were not wrapped individually but in Exp.3S and Exp.4W each tube rhizotron was further wrapped individually in flexible corrugated cardboard to exclude all light. In Exp.4W frost protection was applied to the experiment: From the 1st of December each frame was wrapped in four insulating mats (ROCKWOOL A/S, Denmark) measuring 50 × 1,000 × 3,000 mm covering all sides of the frames. The top was covered in bubble wrap 12.7 mm. The bottom 0.5 m of the tube rhizotron was wrapped with a heating cable (Frost Safe M/T 8.0 m 107 W from Heatcom, Denmark). Each frame had a 4 m long cable (53 W), which was regulated by a thermostat turning on the heat at temperatures below 3 °C and switched off again at 13 °C. The heater and the bubble wrap were used only during periods of frost. Temperatures were recorded by temperature and air humidity loggers (Tinytag Ultra 2 from Gemini Data Loggers) on the top of the frames and inside the frames in different heights.
Irrigation
In Exp.1S the tube rhizotron were continuously irrigated from below using a saucer placed under each tube rhizotron. On the 10th of June the watering was terminated to prevent anaerobic conditions in the lower part of the root zone and for the last 20 days of the experiment the plants survived on the stored soil moisture. In the following experiments this was improved by irrigating each tube rhizotron individually from the top using a drip water dispenser. In Exp.2W and Exp.3S the drip watering dispensers were the type Iriso enkeltdryppere (Modious ApS, Denmark) and in Exp.4W they were the type Universal systems (AQUASOLO SYSTEMS, France). In Exp.3S spare experimental tube rhizotron (4) were included in the experiment. These were removed and weighed periodically to estimate water use without disturbing the experimental tube rhizotrons which were maintained throughout the experiment at 50–80 % field capacity. In Exp.2W and Exp.4W the tube rhizotrons were irrigated once a week according to average monthly precipitation for this period. Thereby the plants were exposed to a situation with excess of irrigation compared to evapotranspiration. Throughout the winter period water was observed to leach out of the tube rhizotrons. During frost periods the plants were not irrigated. During thaw the plants were irrigated with the amount of water withheld during the frost period.
Additional management procedures
Each experiment received one treatment with fungicides, when leaf diseases were observed. In Exp.1S it was a mixture of Tern (from Maktheshim-Agan), Bell and Comet (from Basf A/S), in Exp.2w it was Folicur EC 250 (from Bayer A/S), in Exp.3S and Exp.4W it was Zenit (from Syngenta Crop Protection A/S). In Exp.4W all seeds were treated with CELEST formula M (Syngenta Crop Protection A/S) before seeding. In Exp.4W plants were given a nutrient solution during spring equivalent to 153 kg N ha−1, 25.6 kg P ha−1 and 121 kg K ha−1.
Root and shoot measurements
Shoot measurements were made on experiment termination date. The summer experiments were terminated when the roots reached the bottom of the tube rhizotrons. In Exp.2W plant growth was terminated by frost. In Exp.W the frost protection maintained a temperature in the root zone between 2 and 5 °C above the outside air, in windy and still weather, respectively. Still the plants suffered severe frost damage during winter and Exp.4W was terminated when this became apparent.
The rooting depth was measured on individual tube rhizotrons by recording the depth of the deepest root seen through the transparent interface on the front side of the tube rhizotron. This was done on a weekly to monthly basis depending on depth penetration rates (DPR) of roots and experimental conditions.
Root intensity was measured in the individual tube rhizotrons using the root intersection method as follows. A transparent plastic sheet with a marked grid of eight horizontal and vertical lines (1 × 1 cm grid) of 8 cm height was placed on the tube rhizotrons at selected heights and the number of roots intersecting the lines was recorded. In Exp.2W, Exp.3S and Exp.4W these measurements were taken down the whole soil profile two to three times per experiment. In all experiments root intensity at selected heights were measured more frequently. In Exp.1S the grid contained 64 cm counting line for each depth. In Exp.2W this was reduced to 32 cm counting line for each depth, for Exp.3S the counting line was increased to 48 cm and in Exp4W the counting line was 96 cm for each depth.
Shoots were harvested at the end of each experiment by cutting below the crown of the plant at the soil surface. Dry weight (70 °C for 72 h) and nitrogen content (flash combustion using a FLASH 2000 HT Elemental Analyzer (Thermo Scientific) was determined for each plant.
Statistical analysis
Analysis of variance of cultivar differences within each experiment used the GLM (General Linear Model) procedure of the SAS statistical package (SAS Institute Inc.). For cultivar, cultivar × experiment and cultivar × season interaction across all experiments, analysis of variance used the type 3 test of fixed effects of the MIXED procedure of the SAS statistical package (SAS Institute inc.). Significant differences in the adjusted mean values were determined by the Tukey–Kramer Test of pairwise comparisons at a significance level of p < 0.05. Proc Mixed of SAS was used for estimation of genetic and non-genetic components of variance for subsequent calculation of broad sense heritability within and across experiment.
Calculation of depth penetration rate of roots
Depth penetration rates of roots were compared on a single measuring date for each experiment. Selection criteria for the measuring dates were that mean rooting depths were between 1 and 1.5 m and frost had not yet caused damage. For analysing root growth across different experiments, DPR were calculated as described for wheat in Pedersen et al. (2010). According to Pedersen et al. (2010) the lag phase from seeding to root penetration is 100 °C day−1 at which date the root penetration depth is 100 mm.
The accumulated thermal units (TU) were calculated as follow:
where TU i is the accumulated thermal units (°C day−1) at the root measuring date for experiment i, n i is the total number of days from seeding to the root measuring date in experiment i, T mj is the mean daily temperature on day j, T b is the base temperature, TU ij is the thermal unit in experiment i at day j.
DPR were calculated as follow:
where DPR ik (mm °C−1day−1) is the depth penetration rate in experiment i and tube rhizotron k and R ik is the rooting depth (mm) in experiment i and tube rhizotron k.
Results
Root and shoot growth in 2 m tube rhizotrons
The plants grew well in the 2 m long tube rhizotrons during favourable temperatures, producing around 0.8 to 3.3 g dry biomass per plant, reaching rooting depths of 0.5 to 1.95 m when experiments were terminated. Plants grew larger in the two summer experiments, even though the experimental period was shorter than during autumn/winter experiments (Table 3; Fig. 2). In Table 3 the date and age of the plants for root and shoot measurements are shown.

a Depth penetration rate, b shoot biomass and c nitrogen content of shoots for wheat cultivars represented in two or more experiments. Values are mean ± s.e. for each cultivar. d Heritability of depth penetration rate (DPR), shoot biomass and nitrogen content of shoots in individual experiments and in different seasons
Also depth penetration rate of roots were significantly higher in the summer experiments compared with autumn/winter. Likewise the root intensities in depth were generally higher for the summer experiments. On the other hand the autumn/winter experiments had fast DPR compared to root intensities as seen in Fig. 3b, c.

Root intensity at depth for Exp.2W and Exp.3S at selected dates for wheat cultivars represented in two or more experiments. Values are mean ± s.e. for each experiment. a Root intensity for 52 days after seeding in Exp.2W. b Root intensity at measuring date for root depth penetration rate in Exp.2W. c Root intensity 8 days before measuring date for root depth penetration rate in Exp.3S. d Root intensity 70 days after seeding in Exp.3S
Seasonal and vernalisation effects
In the summer experiments plants produced more tillers compared to the autumn/winter experiments since summer grown plants lacked vernalisation. The shoot biomass varied greatly between the experiments (Fig. 3), with generally more biomass during summer than in the autumn/winter experiments. In contrast, nitrogen concentration in shoots was much higher in autumn/winter (Exp.2W) than in summer experiments (Exp.1S and Exp.3S, Fig. 2). Heritability of shoot biomass was 24–60 % across all experiments whereas heritability of depth penetration rate was only consistently seen in winter experiments (27 %) but not in summer experiments (0.01 %) (Fig. 2d).
Cultivar differences in depth penetration rate of roots, shoot biomass and shoot N content
Across all experiments DPR of roots differed significantly between the cultivars. For the nine cultivars represented in two or more experiments the cultivar with the lowest depth penetration rate was ‘Tuareg’ with an adjusted mean of 1.38 mm °C−1 day−1, and the highest was ‘Mercedes’ with 2.07 mm °C−1 day−1. There was a consistent heritability of shoot nitrogen percentage (12–37 %, Fig. 2d) but not of total shoot nitrogen content (0–61 %). The location of the breeding companies had an influence on depth penetration rate of roots as cultivars bred in Denmark had significantly faster depth penetration than cultivars from other EU countries (p < 0.05) (Fig. 4).

Depth penetration rate plotted against shoot biomass for all wheat cultivars. Values are adjusted means for each cultivar. Cultivars divided in groups according to location of breeding company. Depth penetration rate significantly different for the two groups (p < 0.05)
There was no overall relationship between shoot biomass and DPR of roots, as shoot biomass only correlated with depth penetration rates in Exp.1S and Exp.2W (Fig. 5a). Likewise a correlation between depth penetration rate and total nitrogen in shoot could only be found in Exp.2W (Fig. 5b). Cultivar Mercedes was the only cultivar having both high DPR and total nitrogen content consistently in all experiments (Fig. 2).

a Depth penetration rate in relation to shoot biomass for all wheat cultivars. b Depth penetration rate in relation to nitrogen content in shoot. Values are mean ± s.e. for each cultivar. Linear regressions have been fitted to each experiment
The individual experimental conditions affected the cultivar DPR of roots differently as a significant experiment × cultivar interaction was found (p < 0.05) but no season × cultivar interaction could be detected (p = 0.51). However consistency in ranking of depth penetration was evident for the slowest cultivars (Tuareg, Hereward, Genius) and fastest cultivars (Cordiale, Mercedes) in all experiments (Fig. 3a). For shoot biomass, the experiment had a strong effect and the experiment × cultivar interaction was significant (p < 0.05) but no significant cultivar × season interaction was evident (p = 0.08). Shoot nitrogen percentage was much higher during autumn/winter than summer, but no interaction between cultivar and experiment or season was found.
Discussion
Methodical aspects of long tube rhizotrons, including experimental season for deep root traits studies
Breeding for deeper root systems is desired by plant breeders, but fast root phenotyping methods for soil grown plants have been lacking (Gregory et al. 2009). Growth containers, like the tube rhizotrons used in the present study, have been shown to be a good shortcut to estimate potential root growth in the field (Hurd 1974; Thorup-Kristensen 1998). Using tube rhizotrons in this study we found heritability of 27 % in depth penetration rate of roots during autumn/winter in modern North European winter wheat cultivars. This finding shows the potential of using long tube rhizotrons for studies of root depth development. The experimental set up used in this study has two big advantages compared to a field screening system; (1) Uniformity and control of soil compaction and water potential (2) they are cheaper to set up and it is faster to record the root growth, allowing more studies and comparisons to be made.
The different seasons introduced substantial differences in temperature, light intensity and day length. It was clear that the time of year strongly affected the growth of the plants both above and below ground. The autumn/winter experiments had a much lower biomass production, N uptake and depth penetration rate of roots but higher nitrogen concentration in the plant material. In contrast to heritability of shoot biomass, heritability of depth penetration rate of roots was only consistent in autumn/winter experiments. Likewise it has previously been shown that temperature alone can affect other root traits differently in different genotypes (Kaspar and Bland 1992).
The autumn/winter experiments showed DPR of roots between 1.0 and 1.6 mm °C−1 day−1 whereas the summer experiments had 2.2–2.5 mm °C−1 day1. Previous field studies have found linear relationships between temperature sum and rooting depth of wheat (Liao et al. 2006; Kirkegaard and Lilley 2007; Thorup-Kristensen et al. 2009), but at lower rates than found here, mostly from 0.7 and 1.4 mm °C−1 day1. The DPR from the present study are approximately twice as high as in the field studies, in accordance with an earlier observation that that DPR in growth boxes may be approximately twice as high as in the field (Thorup-Kristensen 1998). However, temperature sum does not take into account the large seasonal differences in day length and irradiation, and the low root DPR during autumn/winter experiments may be explained by the low irradiation during this period. It was observed in the present study that at similar maximum rooting depths the root intensities were lower in the autumn/winter experiments than the summer experiments.
So far only little is known about how root development responds to a combination of low light and low temperatures combined with short days. Regarding rooting depth, field experiments during winter have found a linear relationship between rooting depth and temperature sum during the entire growing season (Gregory et al. 1978; Thorup-Kristensen et al. 2009). More controlled studies have shown that under low temperatures (<12 °C) but normal light intensities the root/shoot ratio increases (Vincent and Gregory 1989; Engels 1994; Equiza et al. 1997) and this response is stronger in spring compared to winter wheat (Equiza et al. 1997). Decreasing light intensities under constant temperature affects root growth more than shoot growth resulting in decreasing root/shoot ratios (Vincent and Gregory 1989; Gloser et al. 1996; Poorter and Nagel 2000; Laghari et al. 2004). In wheat low light intensity is known to decrease seminal root growth (Page et al. 2012) and number of tillers and subsequently the number of nodal root axes decrease (Vincent and Gregory 1989).
Fluctuations in soil temperature in the field are slow and decrease with depth. In Denmark the soil temperature is close to 8 °C at 1 m depth all year round. In the present study the temperatures of the entire root zone were close to air temperature and thus without a vertical gradient, which may obviously affect root growth. Recent studies have shown that the lack of a vertical gradient will decrease both shoot and root growth and alter the root architecture of towards more roots in the deeper soil layers in barley seedlings (Füllner et al. 2012) and less lateral roots in oilseed rape (Nagel et al. 2009). For winter crops the season further influences the phenology of the plants. In the present study the combination of low temperatures and low light intensity combined with short day length in autumn/winter experiments affected the root growth towards a less dense root system.
The outdoor location and constant adjustment of the set-ups meant that environmental factors other than temperature and light differed somewhat between the experiments and a significant experiment × cultivar interaction was observed. However the cultivars tended to have a consistent ranking across the experiments. It is known that environmental factors such as soil compaction and soil moisture levels can influence the ranking in root phenotyping (Motzo et al. 1993). In this study the autumn/winter experiments were closest to the natural conditions of young winter wheat crops, but frost conditions in the rooting zone became more severe, deeper and fluctuated more in the tube rhizotrons than under field conditions, damaging the plants during winter. This is a clearly complicating factor in such experiments, and to perform more representative studies during the autumn/winter season and into the spring better insulated boxes or other solutions protecting the tube rhizotrons from too low temperatures are needed. As heritability of depth penetration rate was absent in summer experiments, autumn/winter experiments seem a necessity for phenotyping deep roots of winter cereals.
Cultivar differences in depth penetration rate
We were able to detect highly significant genotypic differences in DPR between cultivars through combined analysis of four experiments ranging from 1.39 (±0.35) mm °C−1 day−1 to 2.07 (±0.34) mm °C−1 day−1. This would correspond to a difference in rooting depth of around 40 cm in the period from end September to the start of January (growth stage 23–26 on the BBCH scale under Danish temperature conditions). Assuming that the depth penetration rate of roots in tube rhizotron may be twice what is seen in a field soil, this would correspond to a difference of 20 cm rooting depth in field soil. The method reveals cultivar effects, environmental effects as well as cultivar × environmental effects on depth penetration rate of roots and shows potential for initial screening of cultivars for deep root traits using tube rhizotrons ensuring that environmental effects can be accounted for.
The results showed that the cultivars from Danish breeding companies as a group had significantly faster depth penetration than cultivars bred in other countries. As the experiments were performed in Denmark the Danish genotypes were growing in the climate where they were selected but the others not. Part of such local environment conditions are the effect the strict Danish N fertilizer regulation which might have created a selection pressure for increased nitrogen use efficiency and thereby for deep roots. More research will be needed before firm conclusions on effects of origin can be drawn. However, it is promising that the method was able to give results allowing such comparisons.
The few available data on wheat cultivar differences in root traits in the field appear to give consistent results to those in the present study. Cultivar Hereward showed low DPR and low root intensity at depth in this study, which corresponds well with the study of (Ford et al. 2006) where ‘Hereward’ was also amongst the cultivars with a medium to low root density in the whole profile. Gregory et al. (2005) also found that ‘Hereward’ was among the densest in 0.1–0.2 m depths, but intermediate in deeper soil layers. Cultivar Hereford, which in the present study showed intermediate depth penetration rate and high root intensity at depth, in field studies at our institute has shown high root intensity at depth compared to other cultivars (Rasmussen, unpublished data). Further work to compare the ranking of cultivars across field and tube rhizotron studies are required to provide further confidence that it provides a useful initial screening methodology for deep root traits.
Depth penetration rate in relation to NUE and shoot biomass
Rooting depth has been found to correlate well with maximum depths of nitrogen uptake in the field (Thorup-Kristensen 2001; Kristensen and Thorup-Kristensen 2004b). Soil N depletion was not measured in the present study, and a correlation between depth penetration rate and shoot N content was only seen in one out of tree experiments. Only one cultivar—‘Mercedes’—had consistently high depth penetration rate and high nitrogen content in the shoot across the experiments.
It has often been argued that selection for above ground performance such as shoot biomass would indirectly lead to selection for desired root traits such as high DPR of roots. In this study a positive correlation between shoot biomass and DPR were only seen in two out of four experiments. This indicates that a more direct selection for depth penetration rate may be more efficient than the indirect selection for root traits that has historically been applied in plant breeding programs.
Conclusion
Heritability of DPR of roots was 12 and 34 % in two autumn/winter experiments but only 0 and 13 % in two summer experiments, indicating that root screening of winter wheat is not feasible during summer season. Depth penetration was significantly faster for some cultivars than others across experiments (p < 0.0001). The two most contrasting cultivars were ‘Tuareg’ and ‘Mercedes’ with root penetration rates of 1.39 (±0.35) and 2.07 (±0.34) mm °C−1 day−1 respectively.
Significant experiment × genotypic interaction was found indicating other environmental factors playing a major role in genotypic effects on root traits. The high heritability of depth penetration rate in winter experiments indicate tube rhizotrons as a valuable tool for root phenotyping under more relevant conditions and plant/root size than many systems being developed for root phenotyping.
The 2 m length of tube rhizotrons used was appropriate for the purpose of the study. Shorter tube rhizotrons would only allow the study of root growth of very young plants. If frost problems are solved, longer tube rhizotrons may add extra information about root growth of winter cereals in spring and summer.
References
Botwright T, Wade LJ (2012) Genotype × environmental interactins for root depth of wheat. Field Crop Res 137:117–125
Champoux MC, Wang G, Sarkarung S, Mackill DJ, O’Toole JC, Huang N, McCouch SR (1995) Locating genes associated with root morphology and drought avoidance in rice via linkage to molecular markers. Theor Appl Genet 90:969–981
Christopher JT, Manschadi AM, Hammer GL, Borrell AK (2008) Developmental and physiological traits associated with high yield and stay-green phenotype in wheat. Aust J Agric Res 59:354–364
Courtois B, Ahmadi N, Khowaja F, Price AH, Rami J-F, Frouin J, Hamelin C, Ruiz M (2009) Rice root genetic architecture: meta-analysis from a drought QTL database. Rice 2:115–128
Engels C (1994) Effect of root and shoot meristem temperature on shoot to root dry-matter partitioning and the internal concentrations of nitrogen and carbohydrates in maize and wheat. Ann Bot 72:211–219
Equiza MA, Miravé JP, Tognetti JA (1997) Different inhibition of Shoot vs. Root Growth at Low temperatures and its relationship with carbohydrate accumulation in different wheat cultivars. Ann Bot 80:657–663
Ford KE, Gregory PJ, Gooding MJ, Pepler S (2006) Genotype and fungicide effects on late-season root growth of winter wheat. Plant Soil 284:33–44
Foulkes MJ, Hawkesford MJ, Barraclough PB, Holdsworth MJ, Kerr S, Kightley S, Shewry PR (2009) Identifying traits to improve the nitrogen economy of wheat: recent advances and future prospects. Field Crop Res. 114:329–342
Füllner K, Temperton VM, Rascher U, Jahnke S, Rist R, Schurr U, Kuhn AJ (2012) Vertical gradient in soil temperature stimulates development and increases biomass accumulation in barley. Plant, Cell Environ 35:884–892
Gloser V, Scheurwater I, Lambers H (1996) The interactive effect of irradiation and source of nitrogen on growth and root respiration of Calamagrostis epigejos. New Phytol 134:407–412
Gregory PJ, McGowan M, Biscoe PV, Hunter B (1978) Water relations of winter wheat. 1. Growth of the root system. J Agric Sci 91:91–102
Gregory PJ, Gooding MJ, Ford KE, Hendriks PW, Kirkegaard JA, Rebetzke GJ (2005) Genotypic and environmental influences on the performance of wheat root systems. Asp of Appl Biol 73:1–10
Gregory PJ, Bengough AG, Grinev D, Schmidt S, Thomas WTB, Wojciechowski T and Young IM (2009) Root phenomics of crops: opportunities and challenges. Funct Plant Biol (Special Issue: Plant phenomics.) 36, 922–929
Hammer GL, Dong ZS, McLean G, Doherty A, Messina C, Schussler J, Zinselmeier C, Paszkiewicz S, Cooper M (2009) Can changes in canopy and/or root system architecture explain historical maize yield trends in the U.S. Corn Belt? Crop Sci 49:299–312
Hurd EA (1974) Phenotype and drought tolerance in wheat. Agric Meteorol 14:39–55
Kaspar TC, Bland WL (1992) Soil temperature and root growth. Soil Sci 154:290–299
Kirkegaard JA, Lilley JM (2007) Root penetration rate—a benchmark to identify soil and plant limitations to rooting depth in wheat. Aust J Exp Agric 47:590–602
Kristensen HL, Thorup-Kristensen K (2004a) Root growth and nitrate uptake of three different catch crops in deep soil layers. Soil Sci Soc Am J 68:529–537
Kristensen HL, Thorup-Kristensen K (2004b) Uptake of 15N labeled nitrate by root systems of sweet corn, carrot and white cabbage from 0.2-2.5 meters depth. Plant Soil 265:93–100
Laghari KB, Munir M, Farrar JF, Mahar AN (2004) Dry matter partitioning and plant development as function of environmental factors in wheat (Triticum aestivum L.) cv. Avalon. J Food Agric Environ 2:149–156
Liao M, Palta JA, Fillery IRP (2006) Root characteristics of vigorous wheat improve early nitrogen uptake. Aust J Agric Res 57:1097–1107
Lynch JP (2013) Steep, cheap and deep: an ideotype to optimize water and N acquisition by maize root systems in viewpoint: part of a special issue on matching roots to their environment. Ann Bot 112:347–357
McKenzie BM, Bengough AG, Hallett PD, Thomas WTB, Forster B, McNicol JW (2009) Deep rooting and drought screening of cereal crops: A novel field-based method and its application. Field Crop Res 112:165–171
Mi GH, Chen FJ, Wu QP, Lai NW, Yuan LX, Zhang FS (2010) Ideotype root architecture for efficient nitrogen acquisition by maize in intensive cropping systems. Sci China Life Sci 53:1369–1373
Motzo R, Attene G, Deidda M (1993) Genotypic variation in durum wheat root systems at different stages of development in a Mediterranean environment. Euphytica 66:197–206
Mozafar A, Schreiber P, Oertli JJ (1993) Photoperiod and root-zone temperature: interacting effects on growth and mineral nutrients of maize. Plant Soil 153:71–78
Nagel KA, Kastenholz B, Jahnke S, van Dusschoten D, Aach T, Mühlich M, Truhn D, Scharr H, Terjung S, Walter A, Schurr U (2009) Temperature responses of roots: impact on growth, root system architecture and implications for phenotyping. Funct Plant Biol 36:947–959
Page V, Blösch RM, Feller U (2012) Regulation of shoot growth, root development and manganese allocation in wheat (Triticum aestivum) genotypes by light intensity. Plant Growth Regul 67:209–215
Pedersen A, Zhang K, Thorup-Kristensen K, Jensen LS (2010) Modelling diverse root density dynamics and deep nitrogen uptake—a simple approach. Plant Soil 326:493–510
Poorter H, Nagel O (2000) The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: a quantitative review. Aust J Plant Physiol 27:595–607
Rich SM, Watt M (2013) Soil conditions and cereal root system architecture: review and considerations for linking Darwin and Weaver. J Exp Bot 64:1193–1208
Steele KA, Price AH, Shashidhar HE, Witcombe JR (2006) Marker-assisted selection to introgress rice QTLs controlling root traits into an Indian upland rice variety. Theor Appl Genet 112:208–221
Thorup-Kristensen K (1998) Root growth of green pea (Pisum sativum L.) genotypes. Crop Sci 38:1445–1451
Thorup-Kristensen K (2001) Are differences in root growth of nitrogen catch crops important for their ability to reduce soil nitrate-N content, and how can this be measured? Plant Soil 230:185–195
Thorup-Kristensen K, Salmeron Cortasa M, Loges R (2009) Winter wheat roots grow twice as deep as spring wheat roots, is this important for N uptake and N leaching losses? Plant Soil 322:101–114
Vincent CD, Gregory PJ (1989) Effects of temperature on the development and growth of winter wheat roots. I. Controlled glasshouse studies of temperature, nitrogen and irradiance. Plant Soil 119:87–97
Wasson AP, Richards RA, Chatrath R, Misra SC, Prasad SVS, Rebetzke GJ, Kirkegaard JA, Christopher J, Watt M (2012) Traits and selection strategies to improve root systems and water uptake in water-limited wheat crops. J Exp Bot 63:3485–3498
Watt M, Moosavi S, Cunningham SC, Kirkegaard JA, Rebetzke GJ, Richards RA (2013) A rapid, controlled-environment seedling root screen for wheat correlateswell with rooting depths at vegetative, but not reproductive, stages at two field sites. Ann Bot 112:447–455
Acknowledgments
We thank Jeppe Reitan Andersen and Hans Haldrup from Nordic Seed, Else Bøje Nielsen from Ragt Nordics, Finn Borum from Sejet Planteforædling, Claus Nymand and Claire Fremann from KWS, and Dr. Julia Cooper from Newcastle University for providing seeds and information. Special thanks to Dr. John Kirkegaard from CSIRO, Australia for guidance and assistance. Experiments were managed with the help of the staff at University of Copenhagen the Experimental farm in Taastrup. Founds were provided to Dr. K. Thorup-Kristensen by University of Copenhagen, Department of Plant and Environmental Sciences.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Ytting, N.K., Andersen, S.B. & Thorup-Kristensen, K. Using tube rhizotrons to measure variation in depth penetration rate among modern North-European winter wheat (Triticum aestivum L.) cultivars. Euphytica 199, 233–245 (2014). https://doi.org/10.1007/s10681-014-1163-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10681-014-1163-8