
Abstract
Increasing seed oil content is an important breeding goal for Brassica napus L. (B. napus). The identification of quantitative trait loci (QTL) for seed oil content and related traits is important for efficient selection of B. napus cultivars with high seed oil content. To get better knowledge on these traits, a molecular marker linkage map for B. napus was constructed with a recombinant inbred lines (RIL) population. The length of the map was 1,589 cM with 451 markers distributed over 25 linkage groups. QTL for seed oil content, seed hull content and seed coat color in three environments were detected by composite interval mapping (CIM) tests. Eleven QTL accounted for 5.19–13.57% of the variation for seed oil content. Twelve QTL associated with seed hull content were identified with contribution ranging from 5.80 to 22.71% and four QTL for seed coat color accounted for 5.23–15.99% of the variation. It is very interesting to found that co-localization between QTL for the three traits were found on N8. These results indicated the possibility to combine favorable alleles at different QTL to increase seed oil content, as well as to combine information about the relationship between seed oil content and other traits.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Brassica napus L. (B. napus) represents an excellent cash crop in the world. The economic value of derived products such as oil is used for both nutritional and industrial purposes. China is the largest producer with 7.5 million ha of harvested area and 11.9 million tons of production in 2004. Global demand is developing rapidly, but production is presently unable to meet that demand. Improvement in seed oil content of B. napus is one of the important breeding objectives for breeders.
Among the agronomic traits, seed oil content in B. napus is a typical quantitative trait under polygenetic control, which is controlled by a large number of genes with mainly additive and few epistatic gene actions (Zhao 2002) and influenced by environment (Zhao et al. 2005). Furthermore, seed oil content is positively correlated with seed coat color (Rahman et al. 2001), erucic acid content (Cheung and Landry 1998) and negatively correlated with protein content (Zhao et al. 2005) and seed hull content (Wang et al. 2003).
Seed oil content can be improved through the development of yellow-seeded cultivars, which are known to contain higher oil and less fiber content. Yellow seeds of B. rapa and B. napus have 5% and 3–4% lower fiber content, respectively, compared with those of dark-seeded varieties (Stringam et al. 1974). Seeds of yellow-seeded cultivars of B. rapa and B. napus have 2.5% higher oil content than those of the dark-seeded varieties (Daun and DeClercq 1988). The higher seed oil content in yellow-seeded B. napus is partially attributed to the lower seed hull content. The palisade layer is reduced to half to two-third of its thickness in yellow seeds compared with black seeds (Stringam et al. 1974). There seeds also have some other advantages, including higher transparency of the crude oil, higher protein content and lower fiber content with better feeding value for livestocks (Liu et al. 2006).
The identification of QTL represents a first step toward dissecting the molecular basis of these complex traits. Gül (2002) identified six oil QTL, of which two were detected by Ecke et al. (1995) and four of them showed a close negative linkage with protein QTL, explaining the conflict between seed oil content and protein content in seed, while two oil QTL and one protein QTL might be inherited independently, demonstrating the possibility of combining high seed oil and protein content. Burns et al. (2003) found seed oil content QTL on N3, N6, N8, N13, N14, N18, and N19 in B. napus by an intervarietal set of part chromosome substitution lines. Qiu et al. (2006) identified six seed oil content QTL on N4, N8, N10, N12, N13 and N17 by DH population. Eight QTL with additive effects and nine pairs of loci with additive × additive epistasis with high genotype × environment interactions were detected in a cross involving a Chinese and a European parental line (Zhao et al. 2005). Delourme et al. (2006) detected also many genomic regions involved in seed oil content in two DH populations, such as QTL on N3 was revealed in all the studies and the QTL on N1, N8 and N13 were revealed in three studies out of five. Several QTL linked to seed coat color and the hereditary pattern of seed coat color in Brassica species have been reported (Fu et al. 2007). By now, there is some QTL report about the seed hull content, which is likely to be controlled by a number of genes, and therefore appropriate for QTL analysis (Badani et al. 2006).
The objectives of this study were to enrich knowledge on seed oil content and its correlation with seed hull content and seed coat color, through a well constructed linkage map and phenotypic and QTL analysis synchronously in three different environments.
Materials and methods
Plant materials and field experiment
In this study, the RIL were developed through successive selfing up to six generations from a cross between yellow-seeded female parent GH06 and black-seeded male parent P174 by single seed descent. GH06 is a yellow-seeded inbred line and also the parent of the yellow-seeded cultivar “Yuhuang 1” (Li et al. 2001). The RIL were cultivated in 2005/Beibei, 2006/Beibei and 2006/Wanzhou to measure seed oil content, seed hull content and seed coat color. The altitude of Beibei and Wanzhou is about 260 and 1,000 m, respectively. Each plot contained 3 rows with 15 plants per row. Seeds were harvested from open-pollinated plants for trait analysis. Genomic DNA was extracted from 1 g leaves of F2:6 individual plants using the modified CTAB method (Murry and Thomspon 1980) in 2005/Beibei.
Traits measurements
The seed oil content was measured by Soxhlet extractor method (Chinese National Standard GB 2906-82). Seed oil content (%) = 100 × (B − C)/(B − A), A represents paper package weight; B contains paper package weight and sample weight; C contains paper package weight and extracted sample weight.
The seed hull content measurement was conducted according to Wang et al. (2003). Seed was dried at 105°C for 5 h and weighed as seed weight and then hull and embryo were separated. The hull was dried in 105°C for 5 h and weighed as seed coat weight. Seed hull content (%) was estimated as seed coat weight/seed weight × 100%.
The measurement of seed coat color was as described by Fu et al. (2007).
The analysis of SRAP, SSR, AFLP and TRAP markers
The SRAP procedure was performed according to a previous description (Ferriol et al. 2003). In total, 64 forward primers and 64 reverse primers were used, resulting in 4,096 primer combinations. For the design of SRAP primers, the method described by Li and Quiros (2001) was used and the amplified system and procedure were performed with minor modifications.
Sequences of public SSR primer pairs were downloaded from the Brassica database (http://www.ukcrop.net/perl/ace/search/BrassicaDB) and other SSR primer pairs were obtained from Piquemal et al. (2005). The composition of the mixture and the PCR procedure were the same as those reported by Piquemal et al. (2005).
The AFLP analysis was conducted according to Vos et al. (1995) with minor modifications. The genomic DNA was double-digested with restriction enzymes EcoR I and Mse I.
In this study, three genes (RuBPc, FAEl, and FAD2) were analyzed with TRAP markers and the fixed primer was designed according to the EST sequence of the target gene; each forward or reverse fixed primer was combined with different reverse or forward arbitrary primers of SRAP. The amplified system and procedure were conducted refer to Hu and Vick (2003).
The sequence of all these primers was synthesized by Shanghai Sangon Biological Engineering Service Co. Ltd. (China). The PCR reaction was carried out in a PTC-200 thermocycler. The SRAP, SSR, and TRAP PCR products were detected using non-denaturant polyacrylamide gel electrophoresis, while the AFLP PCR products were detected using denaturant polyacrylamide gel electrophoresis.
The SRAP and AFLP polymorphic loci were named according to the primer combination employed, e.g., SRAP marker EM1/ME22 designates the fragment generated by the combination of primer EM1 and ME22 and AFLP marker E3/M6 designates the fragment generated by the combination of primer E3 and M6. The TRAP polymorphic loci were coded according to the combination of the arbitrary primer and the fixed primer of the corresponding gene, e.g., RuBPc ME64 designates the fragment generated by combination of the forward arbitrary primer ME64 and the reverse fixed primer RuBPc. All SSR markers adopted their original names in the Brassica database. If a primer combination detected multiple polymorphic loci, small letters will be assigned to these loci according to the descending fragment size.
Linkage map construction and QTL mapping
The analyses of correlations, means, frequency distributions and other phenotypic analysis were carried out by SAS programme (1989). JoinMap 3.0 (Stam and Ooijen 1995) was used to construct a linkage map with minimum LOD score 3.0. Composite interval mapping (CIM) was conducted to locate QTL with additive effect by WinQTLCart 2.5 (Basten et al. 1999). CIM analysis were calculated using forward regression, a walking speed of 2 cM and a window size of 10 cM and the number of markers for the background control was set to 5. A permutation-based LOD threshold value of more than 2.5 was used to declare a putative QTL (LR ≥ 11.5). The LOD peaks were considered as the most likely position of the QTL. The QTL were named according to the suggestions of McCouch et al. (1997).
Results
Phenotypic analysis of seed oil content, seed hull content and seed coat color
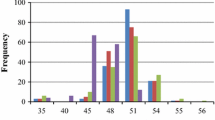
Yellow seeds had significantly higher seed oil content but lower seed hull content than black seeds in the three different environments (Table 1). The difference between the two parents was significant at a level of P < 0.01. Curves depicted the distribution of frequencies of the three traits (Fig. 1). Analyses of variances (ANOVA) demonstrated the presence of significant variability for the three traits in the RIL. Seed oil content and seed hull content in the three environments displayed approximately continuous variations.

The frequency distribution of seed oil content, seed hull content and seed coat color. Abscissa: Phenotypic value of the traits, Ordinate: The number of lines
The correlations of seed oil content with seed hull content and seed coat color were analyzed in the three different environments (Table 2). The correlation between seed oil content and seed hull content were −0.321(P < 0.01), −0.493(P < 0.01) and −0.709(P < 0.01) in 2005/Beibei, 2006/Beibei and 2006/Wanzhou, respectively. Seed oil content was positively related with seed coat color in 2005/Beibei, 2006/Beibei and 2006/Wanzhou [0.208(P < 0.01), 0.272(P < 0.01) and 0.455(P < 0.01), respectively]. Seed hull content was significantly negatively correlated with seed coat color in the three environments, the correlations of 2005/Beibei, 2006/Beibei and 2006/Wanzhou were −0.538(P < 0.01), −0.615(P < 0.01) and −0.715(P < 0.01), respectively.
Construction of the linkage maps
The numbering of the 25 linkage groups was mainly based on the map of Piquemal et al. (2005). The numbers of linkage groups were more than 19 as expected, which implied the interruption within chromosomes caused by large gaps. Twenty-four of our linkage groups were partially homologous to corresponding Piquemal`s linkage groups. One linkage group was constructed by AFLP markers only, which was failed to compare with any other published linkage group. We assigned this linkage group as LG14, which might be N14, because there was no corresponding N14 marker in our linkage groups.
This linkage map covered 1,589 cM and was consisted of 198 SRAP, 140 SSR, 107 AFLP 6 TRAP. The length of each linkage group ranged from 31 to 123 cM and the number of marker on each linkage group varied from 4 to 46. The average distance between adjacent markers was 3.52 cM and ranged from 1.4 to 10.6 cM.
QTL mapping for seed oil content, seed hull content and seed coat color
Results from CIM are summarized in Table 3 and Fig. 2. A total of 5 QTL in 2005/Beibei, 4 QTL in 2006/Beibei and 2 QTL in 2006/Wanzhou were detected for seed oil content. Alleles increasing seed oil content came from GH06 (N3, N4, N5, N7, N8 and N13) and P174 (N1 and LG14), respectively. The qOC05B-1 colocalised with qOC06B-1 and qOC06W-1, all of which had similar additive effects and explained 7.67–13.57% of the phenotypic variation.

Linkage map of B. napus and the locations of quantitative traits loci (QTL) for seed oil content, seed hull content and seed coat color. The QTL and markers were draw by the MAPCHART Version 2.0 software (Voorrips 2002). Cumulative recombination distances are shown on the left (in centiMorgans) and to the right of each linkage group is the marker name
A total of 3 QTL in 2005/Beibei, 6 QTL in 2006/Beibei and 3 QTL in 2006/Wanzhou were detected for seed hull content. Alleles increasing seed hull content came from GH06 (N9 and N12) and P174 (N3, N4, N7 N8, N12 and N15), respectively. Co-location between seed hull content QTL for 2005/Beibei, 2006/Beibei and 2006/Wanzhou was identified on N8 and the contributions for the seed hull content were 17.40, 15.20 and 20.00%, respectively. The additive effects of these three loci were negative, which indicated that P174 increased seed hull content. Two co-localized QTL with similar additive effects were identified on N12 in 2006/Beibei and 2006/Wanzhou, which accounted for 19.10 and 14.40%, of the phenotypic variation for seed hull content, respectively.
Only four seed coat color QTL distributed over the environments accounted for 5.23–15.99% of the phenotypic variation. Alleles increasing seed coat color all came from GH06. The qSC06B-8 was tightly linked with qSC06W-8, both of which had similar additive effects and accounted for 7.03 and 15.99% of the phenotypic variation.
The most important QTL cluster was on N8. One seed oil content QTL accounting for 9.58% variation detected in 2006/Wanzhou was tightly linked with seed hull content QTL detected in the three environments (R 2 = 15.20–20.00%) and two seed coat color QTL detected in 2006/Beibei and 2006/Wanzhou (R 2 = 7.03 and 15.99%).
Discussion
In B. napus, the genome structure is very complex, as the species arose by the hybridization of B. rapa and B. oleracea, although each of the progenitor genomes largely retains its original organization (Parkin et al. 2003). Lombard and Delourme (2001) predicted the length of B. napus genome size is about 2,127–2,480 cM, the length of B. napus genetic map in our study is 1,589 cM. One of our linkage groups was composed of AFLP markers entirely, because maps based on a high number of AFLP, SSR and RFLP frequently exhibited areas of dense clusters (Lacape et al. 2003). Because of the lack of consensus SSR markers for N14, we should increase the number of consensus SSR markers for the convenience of map comparison.
Some research (Ochodzki et al. 2003) indicated that the yellow-seeded B. napus had lower seed hull content. The results of our research strongly confirmed that the seed hull content was negatively correlated with seed oil content and seed coat color. Due to the negative correlation between seed oil and hull content, some of the detected QTL might increase one compound while decreasing the other one. However, few QTL have been successfully used for rapeseed breeding, perhaps because QTL are environmentally sensitive and QTL-environment interaction has been considered as a common phenomenon (Piepho 2000; Cao et al. 2001). Then, genetic analysis and identification of QTL involved in hull content or conditional QTL mapping of seed oil content with respect to seed hull content have to be performed in order to identify regions that influence seed oil content independently from seed hull content in future.
Phenotypic analysis showed that the environment has a significant effect on seed traits and also few QTL was identified in three different environments. The seed oil content in 2005/Beibei trials was lower than that obtained in 2006/Beibei and 2006/Wanzhou. Similarly, the seed hull content in 2006/Wanzhou was lower than that detected in 2005/Beibei and 2006/Beibei. Significant environmental contribution to seed oil content variation was also observed by Zhao et al. (2005) who performed the experiments in very contrasted locations (Germany and China). Similar results were obtained in seed hull content and seed coat color when QTL studies were performed in different environments. Climate and harvest time generally have effects on seed coat color to certain extent, which result in instability of phenotype and QTL analysis.
Six QTL located in a same genomic regions on N8 have effect on three traits. Three of them (qHC05B-8, qHC06B-8 and qHC06W-8) were detected for seed hull content in the different environments, suggesting that they are not, or less affected by the environment. One QTL for seed oil content detected in 2006/Wanzhou and two QTL for seed coat color were identified in this same region on N8, which could co-localized with published QTL detected in other B. napus populations.
Delourme et al. (2006) found that seed oil content QTL on N8 were revealed in only three studies out of five. Burns et al. (2003) found seed oil content QTL on N8 in B. napus by an intervarietal set of part chromosome substitution lines. Qiu et al. (2006) identified six seed oil content QTL on N4, N8, N10, N12, N13 and N17 by DH population. The distribution for seed oil content QTL identified in the RIL populations confirmed the polygenic determinism of this trait. The alleles for increasing seed oil content were mainly derived from GH06 parent. Additive effect of individual QTL ranged from 0.66 to 1.22 in seed oil content. The results were consistent with previous studies performed in oilseed rape where quantitative genetic studies concluded to the prevalence of additive gene action in the control of seed oil content and the alleles increasing oil content are mainly derived from the parent with high seed oil content (Grami and Stefansson 1977; Engqvist and Becker 1991; Shen et al. 2005).
In this research, we first detected a co-localized QTL of seed oil content, seed hull content and seed coat color on N8. And the results also provide important clues for cloning of the candidate gene corresponding to these stable QTL. Further study includes increasing markers and determining how the detected QTL would be used in increasing seed oil content of B. napus potentially by marker-assisted selection.
Abbreviations
- QTL:
-
Quantitative trait loci
- RIL:
-
Recombinant inbred lines
- SRAP:
-
Sequence related amplified polymorphism
- SSR:
-
Simple sequence repeat
- AFLP:
-
Amplified fragment length polymorphism
- TRAP:
-
Target region amplified polymorphism
- cM:
-
Centimorgans
- CIM:
-
Composite interval mapping
- PCR:
-
Polymerase chain reaction
- DNA:
-
Deoxyribonucleic acid
- CTAB:
-
Cetyl trimethyl ammonium bromide
- LOD:
-
Log likelihood ratio
- OC:
-
Seed oil content
- HC:
-
Seed hull content
- SC:
-
Seed coat color
References
Badani AG, Wittkop B, Lühs W, Baetzel R, Horn R, De Haro A, Font R, Friedt W, Snowdon RJ (2006) Colocalization of a partially dominant gene for yellow seed color with a major QTL influencing acid detergent fiber (ADF) content in different crosses of oilseed rape (Brassica napus). Genome 49:1499–1509
Basten CJ, Weir BS, Zeng ZB (1999) QTL cartographer, Version 2.5: programme in statistical genetics. Raleigh, North Carolina State University
Burns MJ, Barnes SR, Bowman JG, Clarke MHE, Werner CP, Kearsey MJ (2003) QTL analysis of an intervarietal set of substitution lines in Brassica napus: (i) seed oil content and fatty acid composition. Heredity 90:39–48
Cao G, Zhu J, He C, Gao Y, Yan J, Wu P (2001) Impact of epistasis and QTL and environment interaction on the developmental behavior of plant height in rice (Oryza sativa L.). Theor Appl Genet 103:153–160
Cheung WY, Landry BS (1998) Molecular mapping of seed quality traits in Brassica juncea L. Czern. and Coss. Acta Hortic 459:139–147
Daun JK, DeClercq DR (1988) Quality of yellow and dark seeds in Brassica campestris canola varieties Candle and Tobin. J Am Oil Chem Soc 65:122–126
Delourme R, Falentin C, Huteau V, Clouet V, Horvais R, Gandon B, Specel S, Hanneton L, Dheu JE, Deschamps M, Margale E, Vincourt P, Renard M (2006) Genetic control of oil content in oilseed rape (Brassica napus L.). Theor Appl Genet 113:1331–1345
Ecke W, Uzunova M, Weißleder K (1995) Mapping the genome of rapeseed (Brassica napus L.). II. Localization of genes controlling erucic acid synthesis and seed oil content. Theor Appl Genet 91:972–977
Engqvist GM, Becker HC (1991) Relative importance of genetic parameters for selecting between oilseed rape crosses. Hereditas 115:25–30
Ferriol M, Pico B, Nuez F (2003) Genetic diversity of some accessions of Cucurbita maxima from Spain using RAPD and SRAP markers. Genet Resour Crop Evol 50:227–238
Fu FY, Liu LZ, Chai YR, Chen L, Yang T, Jin MY, Ma AF, Yan XY, Zhang ZS, Li JN (2007) Localization of QTLs for seed color using recombinant inbred lines of Brassica napus in different environments. Genome 50:840–854
Grami B, Stefansson BR (1977) Gene action for protein and oil content in summer rape. Can J Plant Sci 57:625–631
Gül MK (2002) QTL-Kartierung und Analyse von QTL × Stickstoff Interaktionen beim Winterraps (Brassica napus L.). Ph.D. thesis, Uni Göttingen, Germany, pp 34–38
Hu JG, Vick BA (2003) Target region amplification polymorphism: a novel marker technique for plant genotyping. Plant Mol Biol Rep 21:289–294
Lacape JM, Nguyen TB, Thibivilliers S, Bojinov B, Courtois B, Cantrell RB, Burr B, Hau B (2003) A combined RFLP-SSR-AFLP map of tetraploid cotton based on a Gossypium hirsutum × Gossypium barbadense backcross population. Genome 46(4):612–626
Li G, Quiros CF (2001) Sequence-related amplified polymorphism (SRAP), a new marker system based on a simple PCR reaction: its application to mapping and gene tagging in Brassica. Theor Appl Genet 103:455–461
Li JN, Chen L, Tang ZL, Zhang XK, Yan SJ (2001) Genetic study and commercial application of the yellow-seeded rapeseed (Brassica napus L.). In: Proceedings of the international symposium on rapeseed science, Science Press, New York, Date Month Year, pp 28–34
Liu LZ, Meng JL, Lin N, Chen L, Tang ZL, Zang XK, Li JN (2006) QTL mapping of seed coat color for yellow seeded Brassica napus. Acta Genetica Sinica 33:181–187
Lombard V, Delourme R (2001) A consensus linkage map for rapeseed (Brassica napus L.): construction and integration of three individual maps from DH populations. Theor Appl Genet 103:491–507
McCouch SR, Cho YG, Yano M, Paul E, Blinstrub M, Morishima H, Kinoshita T (1997) Report on QTL nomenclature. Rice Genet Newslett 14:11–13
Murry HG, Thomspon WF (1980) Rapid isolation of high molecular weight DNA. Nucleic Acids Res 8:4321–4322
Ochodzki P, Piotrowska A, Sorensen H (2003) Physical properties and chemical composition of whole seeds, hulls and meals of yellow-seeded rape grown in Poland. 1lth International Rape seed Congress, Copenhagen, Denmark, July, 21
Parkin IAP, Sharpe AG, Lydiate DJ (2003) Patterns of genome duplication within the Brassica napus genome. Genome 46:291–303
Piepho HP (2000) A mixed-model approach to mapping quantitative trait loci in barley on the basis of multiple environment data. Genetics 156:2043–2050
Piquemal J, Cinquin E, Couton F, Rondeau C, Seignoret E, Doucet I, Perret D, Villeger MJ, Vincourt P, Blanchard P (2005) Construction of an oilseed rape (Brassica napus L.) genetic map with SSR markers. Theor Appl Genet 111:1514–1523
Qiu D, Morgan C, Shi J, Long Y, Liu J, Li R, Zhuang X, Wang Y, Tan X, Dietrich E, Weihmann T, Everett C, Vanstraelen S, Beckett P, Fraser F, Trick M, Barnes S, Wilmer J, Schmidt R, Li J, Li D, Meng J, Bancroft I (2006) A comparative linkage map of oilseed rape and its use for QTL analysis of seed oil and erucic acid content. Theor Appl Genet 114:67–80
Rahman MH, Joersbo M, Poulson MH (2001) Development of yellow-seeded Brassica napus of double low quality. Plant Breed 120:473–478
SAS Institute (1989) SAS/STAT users guide: version 6, vol. 2, fourthed. SAS Institute Inc. Cary, North Carolina, USA
Shen JX, Fu TD, Yang GS, Ma CZ, Tu JX (2005) Genetic analysis of rapeseed self-incompatibility lines reveals significant heterosis of different patterns for yield and oil content traits. Plant Breed 124:111–116
Stam PV, Ooijen JW (1995) Join Map(tm) version 3.0: software for the calculation of genetic linkage map, CPRO-DLO Wagemingen, The Netherlands, pp 1–5
Stringam GR, McGregor DI, Pawlowski SH (1974) Chemical and morphological characteristics associated with seed coat color in rapeseed. In: Wratten N, Salisbury PA (eds) Proceedings of the 4th International Rapeseed Conference, The Regional Institute, Gosford, Australia, pp 99–108
Voorrips RE (2002) MapChart: software for the graphical presentation of linkage maps and QTLs. J Hered 1:77–78
Vos P, Hogers R, Bleeker M, Reijans M, van de Lee T, Hornes M, Frijters A, Pot J, Peleman J, Kuiper M, Zabeay M (1995) AFLP: a new technique for DNA fingerprinting. Nucleic Acids Res 23:4407–4414
Wang R, Li JN, Chen L, Tang ZL, Zhang XK (2003) Genetic correlation analysis for main characters in yellow-seeded rapeseed lines (Brassica napus L.). Chin J Oil Crop Sci 25:8–11
Zhao J (2002) QTLs for oil content and their relationships to other agronomic traits in an European × Chinese oilseed rape population. PhD thesis, faculty of agricultural sciences, George-August Uni Göttingen, Germany
Zhao J, Becker HC, Zhang D, Zhang Y, Ecke W (2005) Oil content in an European × Chinese rapeseed population: QTL with additive and epistatic effects and their genotype-environment interactions. Crop Sci 45:51–59
Acknowledgments
Financial support from the State “973” Project (2006CB101604) of China and a grant (30330400) from the Natural Science Foundation of China.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Yan, X.Y., Li, J.N., Fu, F.Y. et al. Co-location of seed oil content, seed hull content and seed coat color QTL in three different environments in Brassica napus L.. Euphytica 170, 355–364 (2009). https://doi.org/10.1007/s10681-009-0006-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10681-009-0006-5