
Abstract
There is no knowledge about the differential capacity of canola genotypes to take up potassium (K) and produce dry matter under conditions of low soil K availability. Hence, 84 canola genotypes were screened for K efficiency in the glasshouse. Plants were grown in sealed pots containing K-responsive, sandy soil without or with K added. Twelve genotypes were selected for advanced screening in the glasshouse in a different K-responsive soil. Genotypes with a mean K efficiency ratio (the ratio of shoot dry weight at deficient and adequate K supply) greater than one standard error above or below the median genotype value were classified as K-efficient or K-inefficient, respectively. There were significant differences between genotypes in the K efficiency ratio in both screening experiments, indicating that genotypes responded differently to K availability. In the initial screening experiment, 19 genotypes were rated as K-efficient and nine genotypes rated as K-inefficient based on the K efficiency ratio. In the advanced screening experiment with 12 genotypes, three genotypes were rated as K-efficient and two as K-inefficient. Genotypes Wesbarker and Rainbow were K-efficient and Genkai K-inefficient in both experiments. Correlation of the K efficiency ratio with (i) shoot K content in the initial and advanced screening and (ii) shoot K concentration in the advanced screening, indicates that the observed differences in K efficiency were due to genotypic differences in both the uptake and the utilization of K. K-efficient genotypes have a potential to improve canola yields on soils with low K availability.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Canola (Brassica napus L.) is the main oilseed crop in Australia, with 1 million ha sown in 2005. South-western Australia accounts for about 44% of the national canola production (Oilseeds 2006). Typical soils are deeply weathered; sandy topsoils with kaolinite clay fractions predominate (Tennant et al. 1992). Most soil K exists either in the exchangeable fraction or is within silicate minerals; therefore, continued removal of K from the exchangeable fraction in grain and other farm products may result in the depletion of soil K (Pal et al. 2001). Since the early 1990s, the incidence of K deficiency and profitable responses to K application in cereal crops (Leach 1981; Anderson et al. 1992; Edwards 1997; Brennan et al. 2004) has become prevalent on sandy and duplex soils in the region. Yield responses to applied K have now also been reported for canola crops on sandy soils in south-western Australia (Brennan and Bolland 2006), where application of K fertilizer is widely adopted to avoid K deficiency and achieve high yields from canola crops.
Nutrient-efficient canola varieties are capable of producing high yields with low nutrient inputs and therefore have the potential to increase the productivity and sustainability of grain production systems on soils with variable or low nutrient availability. Several studies have reported significant differences between canola genotypes in efficiency of nitrogen (N) uptake and utilization (e.g. Grami and La Croix 1977; Yau and Thurling 1987; Svečnjak and Rengel 2006). No study has investigated the genetic variation for K efficiency in canola. However, significant intraspecific variation for K efficiency has been reported for many other crops, including wheat (Woodend and Glass 1993; El Bassam 1998; Guoping et al. 1999; Damon and Rengel 2004), barley (Jensen and Petterson 1980; Glass and Perley 1980), rice (Yang et al. 2004), snap beans (Shea et al. 1968), soybean (Sale and Campell 1987) and tomato (Chen and Gabelman 1995).
Potassium-efficient crop genotypes are tolerant to low K availability in soil and have the capacity to produce high yields in soils with adequate and sub-optimal K availability. The K efficiency ratio is the ratio of growth at deficient and adequate K availability (cf. Rengel and Graham 1995), hence it is a quantitative measure of genotypic response to soil K availability. Sattelmacher et al. (1994) proposed that nutrient efficiency can be attributed to two multi-factorial components: (i) effectiveness in taking up nutrients from the soil and/or (ii) the efficiency with which nutrients are used within the plant to produce yield (utilization efficiency). By assessing genotypes according to their K efficiency ratio, we consider the combined effects of both uptake and utilization efficiency on the growth of canola.
The aims of this study were to (i) determine the extent of genotypic variation in K efficiency of canola, and (ii) identify canola genotypes contrasting in K efficiency. Potassium-efficient and -inefficient genotypes identified in this study will be later tested in the field. Further work will identify relevant genes and/or markers for use in canola breeding programmes.
Materials and methods
Experiment 1—Preliminary screening
The experiment was conducted in an evaporatively cooled glasshouse in Perth, Western Australia (31.58 S, 115.49 E) from October to November, with mean day/night temperatures of 24/15°C and a mean daylength of 12 h and 15 min. The experiment was set-up in a randomized block design with 84 genotypes, two K treatments, and two replicates.
Virgin brown sand (Uc4.22, Northcote 1971) was collected from a bushland site 15 km south-east of Lancelin, Western Australia (31.08 S, 115.29 E). The soil contained 16 mg/kg bicarbonate-extractable K (Colwell and Esdaile 1968), 2.7 mg/kg bicarbonate-extractable P (Colwell 1963), 10.3 mg/g organic carbon (Walkley and Black 1934) and pH (1:5 soil:0.01 M CaCl2 w/v) (Rayment and Higginson 1992) was 4.95. Air-dried soil sieved to 2 mm was placed in plastic-bag-lined pots (9 cm square and 18 cm deep) at 1,000 g per pot. Basal and treatment nutrients were applied in solution to the soil surface and allowed to dry before being thoroughly mixed through the entire soil volume. Basal nutrients were supplied at the following rates (mg/kg soil): 40.9 Ca; 20.9 P; 65.8 N; 54.2 S; 7.1 Mg; 2.9 Zn; 3.2 Mn; 0.6 Cu; 0.1 B; 0.1 Co; and 0.1 Mo. Nitrogen was resupplied at the same rate at 3 and 5 weeks after sowing. Potassium was supplied at 88 mg/kg soil for the adequate K treatment, or not supplied for the deficient K treatment.
Eighty four canola (Brassica napus) genotypes were selected to achieve the widest genetic diversity. Seeds were sown at 10 per pot and seedlings were thinned to three plants per pot after 14 days. Polythene beads were used to cover the soil surface to reduce evaporation. Pots were watered with deionized water daily and weighed to field capacity (10% w/w) on alternate days.
Plants were harvested 37 days after sowing in the late vegetative stage. At harvest, shoots were removed at the cotyledon node and dried in a forced-air oven at 65°C for 7 days before recording dry weights. Shoots were ground and digested in HNO3/HClO4 acids (modified from Johnson and Ulrich 1959) before measuring K concentration by inductively coupled plasma atomic emission spectrometry (Fisons ARL, model 3580 B, Los Angeles, USA). Shoot K content was calculated as the product of shoot K concentration and dry weight.
The K efficiency ratio quantifies the relative tolerance of genotypes to low K availability in terms of shoot growth (growth at sub optimal K supply/growth with adequate K). Data were subjected to analysis of variance and correlation analysis (GenStat 8 Committee 2005). Significance intervals were constructed for the K efficiency ratios according to Rengel and Graham (1995), whereby genotypes with a mean K efficiency ratio more than one standard error above or below the median were classified as K-efficient or K-inefficient, respectively.
Genotypes were selected from Experiment 1 for advanced screening in Experiment 2 based on their K efficiency ratio and shoot dry weight at deficient and adequate K supply. Significance intervals were constructed for shoot dry weight in the same way as for the K efficiency ratio. Means greater than one standard error above or below the median value were classified as good or poor growth, respectively. Shoot dry weights not significantly different from the median were classified as moderate growth.
Significance intervals were constructed for shoot K concentration and content in the same way as described for shoot dry weight and the K efficiency ratio. Shoot K concentration was assumed to indicate efficiency for K utilization (Gerloff 1987), as it represents the relative amount of shoot biomass produced per unit of K taken up; genotypes with a low shoot K concentration produced a large shoot biomass relative to the amount of K taken up and were efficient for K utilization. Shoot K content was assumed to represent efficiency for K uptake; genotypes with high shoot K content took up a large amount of K and were efficient for K uptake.
Experiment 2—Advanced screening
Experiment 2 was conducted in an evaporatively cooled glasshouse in Perth, WA, from April to May, with mean day/night temperatures of 21/16°C and a mean day length of 11 h and 6 min. The experiment was set up in a randomized block design with 12 genotypes, two K treatments, and three replicates. Genotypes were selected from Experiment 1 as either K-efficient (seven genotypes) or K-inefficient (five genotypes) based on K efficiency ratio, K uptake, and growth at deficient and adequate K supply.
Pale brown Karakatta sand (Uc5.11, Northcote 1971) was collected from a fallowed paddock at the University of Western Australia Research Station in Shenton Park, Perth (31.57 S, 115.48 E). The soil contained 19 mg/kg bicarbonate-extractable K (Colwell and Esdaile 1968), 46 mg/kg bicarbonate-extractable P (Colwell 1963), 6.9 mg/g organic carbon (Walkley and Black 1934) and pH (1:5 soil:0.01 M CaCl2 w/v) (Rayment and Higginson 1992) was 6.4. Air-dried soil sieved to 2 mm was placed in plastic-bag-lined pots (20 cm diameter and 15 cm deep) at 5 kg per pot. Basal and treatment nutrients were applied as in Experiment 1. Nitrogen was re-applied at the same rate at 3 weeks after sowing and every 2 weeks thereafter. Potassium was applied at 0 or 88 mg/kg soil for the deficient and adequate K treatments, respectively. Canola seed was dusted with Thiram powder (800 g/kg tetramethylthiuram disulfide) and sown at 10 seeds per pot. Plants were thinned to six per pot on emergence. Pots were watered with deionized water daily and weighed to field capacity (10 % w/w) on alternate days.
Six weeks after sowing three average-sized plants per pot were harvested by removing whole shoots at the cotyledon node. Shoots were dried, weighed and digested as in Experiment 1 and analysed for K by atomic absorption spectrometry (AAnalyst 300, Perkin Elmer, Norwalk, CT, USA). The K efficiency ratios and significance intervals were constructed in the same way as in Experiment 1. Data were subjected to analysis of variance and correlation analysis as in Experiment 1.
Results
Genotypic differences in plant growth response to K deficiency
Plants started to develop leaf chlorosis consistent with K deficiency symptoms (interveinal and leaf margins) 22 days after sowing in Experiment 1 and 34 days after sowing in Experiment 2. The extent and timing of leaf chlorosis varied between genotypes. All genotypes were showing reduced growth and chlorosis at deficient K supply by harvest in Experiment 1 (day 37) and 6 weeks after sowing in Experiment 2. In both experiments genotypes differed significantly in terms of shoot dry weight at deficient (P < 0.001) and adequate K supply (P < 0.001) (Table 1).
Mean shoot dry weight at deficient K supply ranged from 0.34 to 0.86 g/plant in Experiment 1 and from 0.34 g/plant (Genkai) to 1.31 g/plant (BLN301) in Experiment 2. At adequate K supply, mean shoot dry weight ranged from 0.80 to 1.62 g/plant in Experiment 1 and from 0.75 g/plant (Wesbarker) to 1.9 g/plant (BLN5) in Experiment 2.
There was a significant interaction between the genotype effect and K supply for shoot dry weight in Experiment 1 (P < 0.001) (Table 1), indicating that genotypes responded differently to K treatment. The K efficiency ratio (relative shoot weight at deficient and adequate K supply) varied significantly between the 84 genotypes in Experiment 1 (P < 0.001) (Table 1), ranging from 0.35 to 0.84 (Fig. 1). Based on the K efficiency ratio, 19 genotypes were ranked as K-efficient and 9 as K-inefficient. Of the 19 K-efficient genotypes, 12 had poor growth at adequate K supply (Table 2). However, the K efficiency ratio was correlated with shoot weight at both adequate and deficient K supply and was not biased by the influence of either K treatment (Table 3).

K efficiency ratios for 84 genotypes of canola grown for 37 days in Lancelin soil (fertilized with 0 or 88 mg K/kg soil) in the glasshouse (Experiment 1). The vertical dashed lines separate K efficiency intervals at one standard error of the genotype effect above and below the median. I = K-inefficient, M = medium and E = K-efficient
Although there was no significant interaction between the genotype effect and K supply for shoot dry weight in Experiment 2 (Table 1), the K efficiency ratio differed significantly among genotypes (P < 0.001) (Table 1) ranging from 0.42 to 0.88 (Fig. 2). Genotypes IB1368 and Genkai were ranked as K-inefficient and BLN 301, Rainbow and Wesbarker as K-efficient based on the significance interval constructed at one standard error (of the genotype effect) above and below the median genotype value.

K efficiency ratios for 12 genotypes of canola grown in Karrakatta sand (fertilized with 0 or 88 mg K/kg soil) in the glasshouse (Experiment 2). The horizontal dashed lines are one standard error of the genotype effect above and below the median. E = K-efficient, M = medium and I = K-inefficient. Letters denote differences among means based on lsd (5%). Mean values followed by the same letter are not significantly different
Genotypic differences in K content and concentration in shoots
Potassium concentration in shoots varied significantly between genotypes at deficient (P < 0.001) and adequate K supply (P < 0.001) (Table 1) in both experiments. When K supply was deficient, K concentration in shoots ranged from 2.4 to 7.5 mg/g in Experiment 1 and from 6.7 (BLN301) to 16.4 mg/g (Genkai) at 6 weeks after sowing in Experiment 2 (Fig. 3). Shoot K concentration at deficient K supply was significantly correlated with shoot K concentration at adequate K supply in Experiment 1, but not in Experiment 2 (data not shown).

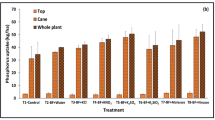
Shoot K content (columns) and tissue concentration (points) for canola plants grown in the glasshouse in K-deficient Karrakatta soil without K applied (Experiment 2). Plants were harvested 8 weeks after sowing. Error bars are ±s.d.. Genotypes are in order of ascending mean K efficiency ratio
There was a significant interaction between genotype and K treatment for shoot K concentration in both experiments (Table 1). Genotypes differed in their capacity to accumulate luxury concentrations of K in shoots where K supply in soil was adequate, or in their capacity to tolerate low concentrations of K in shoots when K supply in soil was deficient. Shoot K concentration at deficient K supply was negatively correlated with the K efficiency ratio and shoot weight at deficient K supply in Experiment 2 (Table 3), suggesting a dilution effect by increased growth. However, there was no such significant correlation in Experiment 1 (Table 3). Ability to tolerate a low K concentration in shoots under deficient K supply was a significant factor determining the response to K in Experiment 2, but not in Experiment 1.
There were significant differences between genotypes in shoot K content at deficient and adequate K supply (P < 0.001) and a significant interaction between genotype and K treatment (P < 0.001) in both experiments (Table 1). Shoot K content in Experiment 1 ranged from 1.51 to 5.28 mg K/plant at deficient K supply and from 18.4 to 28.6 mg K/plant at adequate K supply. IB1368 had the highest shoot K content at deficient K supply. However, despite its excellent capacity to accumulate K, IB1368 was not efficient at utilizing K and did not produce a significantly high shoot weight at either K treatment.
The correlation between the content and concentration of K in shoots at deficient K supply differed between Experiments 1 and 2. In Experiment 1, genotypes with a high shoot K concentration at deficient K supply accumulated a large amount of K in shoots (r = 0.79, P < 0.001) (Table 3). Capacity to take up K was not limited by capacity to tolerate low shoot K concentration and the K efficiency ratio was not correlated with shoot K concentration at deficient K supply. For Experiment 2, however, genotypes with low shoot K concentration at deficient K supply accumulated a large amount of K in shoots (r = −0.42, P < 0.01), produced large shoot dry weight at deficient K supply (r = −0.90, P < 0.001) and were K-efficient in terms of the K efficiency ratio (r = −0.40, P < 0.01) (Table 3). In Experiment 2, capacity to tolerate low concentrations of K in shoots was highly correlated with the capacity of canola plants to take up K from soil and produce shoot dry weight at deficient K supply. It appears that the K efficiency ratio was influenced by an interaction of K uptake and utilization at deficient K supply (Table 3); however, there was no consistent or simple relationship between K efficiency ratio and capacity to take up or utilize K for both experiments.
K-efficient and K-inefficient genotypes
Genotypes Monty, CBWA 002, Ripper and BLN 313 were selected from Experiment 1 for advanced screening as K-inefficient genotypes for their low K efficiency ratio and good-to-moderate growth at adequate K supply according to the significance intervals (Table 2). Although Genkai had poor growth at adequate K, it was selected as K-inefficient for its extremely low K efficiency ratio and poor growth at deficient K supply (Fig. 1, Table 2). Wesbarker, Pinnacle and Rainbow were selected for advanced screening as K-efficient genotypes for their high K efficiency ratio and good growth at deficient K supply (Table 2). Genotypes 988, BLN 5 and BLN313 had low-to-moderate K efficiency ratios, but were selected for further investigation as K-efficient genotypes for their excellent growth at both K treatments. IB1368 was selected as K-efficient for its high K efficiency ratio and excellent K uptake at deficient K supply (Table 2).
In Experiment 2, Genkai and IB1368 were inefficient and Wesbarker, Rainbow and BLN301 were efficient for shoot dry weight (Fig. 2). IB1368 was K-efficient in Experiment 1, with the highest K uptake at deficient K supply but inefficient K utilization (high K concentration). However in Experiment 2, IB1368 was inefficient for K utilization (high shoot K concentration), but it had low K uptake at deficient K supply relative to other genotypes and was categorized as K-inefficient due to its poor K efficiency ratio.
Discussion
Significant differences were found among canola genotypes in efficiency for K uptake and utilization in the pot study. No other study to date has reported genotypic variation in K-efficiency for canola. Genotypes were identified that were consistently efficient (Wesbarker and Rainbow) or inefficient (Genkai) for uptake and utilization of K in the two experiments.
The potential mechanisms for genotypic variation in K efficiency are numerous and should be considered according to the two principal components: (i) effectiveness in taking up nutrients from the soil (uptake efficiency) and/or (ii) the efficiency with which nutrients are used within the plant to produce yield (utilization efficiency) (Sattelmacher et al. 1994). In terms of K utilization efficiency, genotypes may differ in capacity to translocate K at a cellular or whole plant level, or to substitute K+ with other osmolytes (eg. NO3 −, soluble sugars or amino acids) or cations (eg. Na+, Ca2+). In terms of K uptake efficiency, genotypes may differ in root morphological parameters (e.g. root length, rate of root turnover or the number and length of root hairs) or in root physiological parameters (e.g. capacity for high affinity K uptake or capacity to alter K availability in the rhizosphere) (see Sattelmacher et al. 1994).
Genotypic differences in K utilization efficiency have been reported for species other than canola. Several studies have associated genotypic differences in grain yield or shoot biomass at sub-optimal K supply with differences in K concentrations in shoots for a variety of species (e.g. Shea et al. 1968; Woodend and Glass 1993; Yang et al. 2004). For canola, a study of genotypic differences in N efficiency (Svečnjak and Rengel 2006) reported that N-efficient cultivars produced larger plant biomass and contained lower concentrations of N in shoots than N-inefficient cultivars. Yau and Thurling (1987), similarly, reported that capacity to utilize N efficiently (growth rate per unit of N taken up) was correlated with growth rate of canola plants in the field at the pre-flowering stage. In the present study, K concentration at deficient K supply was negatively correlated with shoot dry weight at deficient K supply and the K efficiency ratio in Experiment 2, but not Experiment 1. Shoot K content at deficient K supply was positively correlated with shoot dry weight at deficient K supply and the K efficiency ratio in the two experiments. However, for K efficiency (as the ratio of growth at deficient and adequate K availability), there is no simple relationship with either K uptake or K concentration in shoots. Correlation of the K efficiency ratio with both K concentration and K content in shoots indicates that the observed differences in K efficiency in this study were associated with differences in both K uptake and K utilization efficiency.
Yau and Thurling (1987) reported that the six most efficient cultivars for N utilization out of 40 cultivars studied were derived from hybridization of B. napus and B. rapa (ssp. campestris). They proposed that the capacity for high N-utilization efficiency of these cultivars might have been inherited from B. rapa. In the study presented here, the B. rapa genotypes IB1368, IB1377 and IB1524 were inefficient for K utilization (having high concentration of K in shoot), but were efficient for K uptake. The Japanese cultivars Genkai and Tokiwa, derived from hybridization of B. napus and B. rapa (see Yau and Thurling 1987) were inefficient for K utilization and uptake. B. rapa, as a species, may be less efficient for K utilization and more efficient for K uptake than B. napus. The inefficiency for K-utilization observed in Japanese genotypes Genkai and Tokiwa might have been inherited from the B. rapa parent line, whereas efficiency for K uptake was not inherited.
Although K efficiency is best evaluated in terms of yield, which in the case of canola is grain and specifically, oil, we have chosen to look only at the vegetative stage in this experiment. Practical limitations restrict the number of genotypes that can be assessed if plants are to be grown to maturity. Field experiments in the next phase of research will assess differences in K efficiency in terms of grain and oil yield at maturity for a small number of genotypes identified in this study to differ in K efficiency.
Yau and Thurling (1987) reported that genotypic response of B. napus to N at the pre-flowering stage was highly correlated with response to N at maturity in terms of grain yield. With regard to K efficiency, studies with other species have reported that K efficiency for grain yield was a product of K efficiency for shoot growth and harvest index (Woodend and Glass 1993; Yang et al. 2004). Whether the variation among canola genotypes in K efficiency for shoot growth in the present study will translate to similar variation in K efficiency for grain yield is yet to be determined in the field at various locations in the wheatbelt of Western Australia.
References
Anderson WK, French RJ, Seymour M (1992) Yield responses of wheat and other crops to agronomic practices on duplex soils compared with other soils in Western Australia. Aust J Exp Agric 32:963–970
Brennan RF, Bolland MDA (2006) Soil and tissue tests to predict the potassium requirements of canola in south-western Australia. Aust J Exp Agric 46:675–679
Brennan RF, Bolland MDA, Bowden JW (2004) Potassium deficiency, and molybdenum deficiency and aluminium toxicity due to soil acidification, have become problems for cropping sandy soils in south-western Australia. Aust J Exp Agric 44:1031–1039
Chen JJ, Gabelman WH (1995) Isolation of tomato strains varying in potassium acquisition using a sand-zeolite culture system. Plant Soil 176:65–70
Colwell JD (1963) The estimation of the phosphorus fertilizer requirements of wheat in southern New South Wales by soil analysis. Aust J Exp Agric Ani Husb 3:190–197
Colwell JD, Esdail RJ (1968) The calibration, interpretation, and evaluation of tests for the phosphorus fertilizer requirements of wheat in northern New South Wales. Aust J Soil Res 6:105–120
Damon PM, Rengel Z (2004) Genotypic variation in potassium efficiency of wheat. In: R Horne, N Morgan, W Parker, K Regan, M Harries and A Douglas (eds) Agribusiness crop updates 2004: Cereals. 18–19 February 2004, Perth, Western Australia, pp 47–48. http://www.agric.wa gov.au Cited January 2007
Edwards N (1997) Potassium fertilizer improves wheat yield and grain quality on duplex soils. In: Wong MTF, Pal Y, Edwards N (eds) Proceedings of the first workshop on potassium in Australian agriculture, UWA Press, Perth, pp 69–75
El Bassam N (1998) A concept for selection of ‘low input’ wheat varieties. Euphytica 100:95–100
GenStat 8 Committee (2005) GenStat Reference Manual. VSN International, Oxford, UK
Gerloff GC (1987) Intact-plant screening for tolerance of nutrient deficiency stress. Plant Soil 99:3–16
Glass ADM, Perley JE (1980) Varietal differences in potassium uptake by barley Hordeum vulgare. Plant Physiol 65:160–164
Grami B, La Croix LJ (1977) Cultivar variation in total nitrogen uptake in rape. Can J Plant Sci 57:619–624
Guoping Z, Jingxing C, Tirore A (1999) Genotypic variation for potassium uptake and utilization efficiency in wheat. Nutr Cycl Agroecosyst 54:41–48
Johnson CM, Ulrich A (1959) Analytical methods for use in plant analysis. Californian Agricultural Experimental Station. Bulletin no. 766
Leach BJ (1981) Potassium deficiency with continuous cropping of wheat on sandplain soils. Our Land 13:11–14
Northcote KH (1971) A factual key for the recognition of Australian soils. Rellim, Glenside, South Australia
Oilseeds WA (2006) WA Canola Production Benchmarks 2006. http://www.australianoilseeds.com/__data/page/311/Canola_Benchmarks_for_WA_2006.pdf. Cited 8 June 2006
Pal Y, Gilkes RJ, Wong MTF (2001) Soil factors affecting the availability of potassium to plants for Western Australian soils: a glasshouse study. Aust J Soil Res 39:611–625
Jensen P, Petterson S (1980) Varietal variation in uptake and utilization of potassium (rubidium) in high-salt seedlings of barley. Physiol Plant 48:411–415
Rayment GE, Higginson FR (1992) Australian laboratory handbook of soil and water chemical methods. Inkata Press, Melbourne, Vic
Rengel Z, Graham RD (1995) Wheat genotypes differ in Zn efficiency when grown in chelate-buffered nutrient solution. I. Growth. Plant Soil 176:307–316
Sale PWG, Campbell LC (1987) Differential response to K deficiency among soybean cultivars. Plant Soil 104:183–190
Sattelmacher B, Horst W, Becker HC (1994) Factors that contribute to genetic variation for nutrient efficiency of crop plants. Zeit für Pflanz und Bodenk 157:215–224
Shea PE, Gerloff GC, Gabelman WH (1968) Differing efficiencies of potassium utilization in strains of snapbeans, Phaseolus vulgaris L. Plant Soil 28:337–346
Svečnjak Z, Rengel Z (2006) Canola cultivars differ in nitrogen utilization efficiency at vegetative stage. Field Crops Res 97:221–226
Tennant D, Scholz JD, Purdie B (1992) Physical and chemical characteristics of duplex soils and their distribution in the south-west of Western Australia. Aust J Exp Agric 32:827–843
Walkey A, Black IA (1934) An examination of the Degtjareffe method for determining soil organic matter and a proposed modification of the chromic acid titration method. Soil Sci 37:29–38
Woodend JJ, Glass ADM (1993) Genotype–environment interaction and correlation between vegetative and grain production measures of potassium use-efficiency in wheat (T. aestivum L.) grown under potassium stress. Plant Soil 151:39–44
Yang XE, Liu JX, Wang WM, Ye ZQ, Luo AC (2004) Potassium internal use efficiency relative to growth vigor, potassium distribution and carbohydrate allocation in rice genotypes. J Plant Nut 27:837–852
Yau SK, Thurling N (1987) Variation in nitrogen response among spring rape (Brassica napus) cultivars and its relationship to nitrogen uptake and utilization. Field Crops Res 16:139–155
Acknowledgements
We thank Mr M. Sanders and Prof. W. Cowling, Canola Breeders WA; Mr K. Murray, Australian Temperate Field Crops Collection; Mr T. Trent and Mr P. Carmody, Department of Agriculture WA; and Dr P. Si, The University of Western Australia for providing the seed used in this experiment. This research was funded by the Grains Research and Development Corporation, Australia.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Damon, P.M., Osborne, L.D. & Rengel, Z. Canola genotypes differ in potassium efficiency during vegetative growth. Euphytica 156, 387–397 (2007). https://doi.org/10.1007/s10681-007-9388-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10681-007-9388-4