
Abstract
The objectives of this study were to evaluate some of the popular rotation crops grown in Hungary for tolerance to low external Mn2+ levels and to determine the critical tissue concentration of Mn2+ deficiency during early stages of growth. The minimum Mn2+ concentration required in soil nutrient contents was 42.5 mg kg−1 for sunflower, 24.3 mg kg−1 for tobacco and 10.2 mg kg−1 for triticale. Sunflower, tobacco and triticale achieved optimum growth at 48.0–65.0 mg Mn2+ kg−1, 24.9–32.1 mg Mnn+ kg−1 and 28.7 to 29.6 mg Mn2+ kg−1, respectively. Critical shoot Mn2+ concentration at early stages of growth was 53.6 mg kg−1 in sunflower, 458.0 mg kg−1 in tobacco and 193.8 mg kg−1 in triticale. Our results demonstrate that the tolerance to low external Mn2+ (triticale: <30.2 mg kg−1; sunflower: <56.2 mg kg−1; tobacco: <69.3 mg kg−1) and the critical tissue Mn2+ levels for deficiency varied significantly between crop species tested.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Manganese-deficient soils are widely distributed around the world (Aubert and Pinta 1977; Adriano 1986; Wong et al. 1998), including the Great Hungarian Plain (IPCS 1981; Sillanpää 1982; Baranyai et al. 1987; David and Philippe 1995; Howe et al. 2004). When savanna land is cultivated, a large amount of nitrogen, phosphatic and potassic fertilizer, which usually contains no manganese materials, must be applied. Loam has sometimes also been recommended for soil amelioration. Although these applications are considered to be fundamental for increasing soil N, P and K availability in the area, it is not always recognized that Mn is simultaneously added to the soils (Uren 1990). Maintenance of field Mn fertility is often overlooked, and Mn-deficiency symptoms in crops are sometimes confused with Fe and Mg deficiency. Since concentrated fertilizers with low Mn content are now widely used, Mn deficiency problems appear more often.
Numerous studies have shown that soil manganese exists principally in two forms: as the divalent cation Mn2+ (soluble, mobile, easily available for plants) and as the tetravalent cation Mn4+ (practically insoluble, immobile and unavailable for plants) (Aubert and Pinta 1977). Manganese materials are easily remobilized and leached by precipitation and often accumulate in subsoil layers (IPCS 1981; Centre for Ecology and Hydrology 1983). Singh (1974) applied 54Mn-labelled MnSO4 and rain water equivalent to 300 mm precipitation to surface soil columns and determined the residual Mn2+ distribution in soil layers. He noted that Mn2+ moved to a depth of 18 cm in soil A-horizon and throughout the whole of soil B-horizon, although a higher concentration of Mn2+ was retained by the upper 3 cm of soil. A similar pattern but with relatively higher contents at depths below 3 cm resulted on leaching with water equivalent to 1,200 mm of precipitation. Hadimani et al. (1974) measured the effect of water-soluble organic substances from teak-, bamboo-, eucalyptus- and wild curry-leaves; casuarina needles; citric acid; glucose solution; and distilled water on the manganese distribution in sandy soil column layers in a semiarid environment. They found that organic extracts as well as citric acid caused movement of manganese but retained it presumably in the form of organo-mineral complex in the upper layers of soil columns. In soil column loaded with glucose solution, the manganese moved and accumulated in deeper layers. Columns treated with distilled water also showed the movement of manganese into deeper layers.
Manganese deficiency symptoms more commonly occur at early stages of growth in fields but disappear at later stages once roots reach deeper layers of soil where there is substantial amount of accumulated Mn2+. Sillanpää (1982), Baranyai et al. (1987) and Feher (2005) examined five Hungarian sandy soils (World Reference Base for Soil Resources: Lamellic Arenosol) and showed that the organic Mn2+ concentration in the A-horizon soils decreased to two-quarters about 20 yr of cropping. Presently, increased yields may accelerate this tendency. Shihua et al. (2004) studied manganese deficiency in wheat (Triticum aestivum L.) under rice (Oryza sativa L.) and wheat rotation in most rice-growing areas in China and showed that both total and available Mn increased with soil depth in soils with rice–wheat rotations showing significant spatial variability of Mn in the soil profile. Manganese leaching was the main pathway for Mn loss in coarse-textured soil with high pH, while excessive Mn uptake was the main pathway for Mn loss in clay-textured and acid soil. When Mn was deficient in the topsoil, sufficient Mn in the subsoil supplied Mn nutrition to wheat and contributed to better growth; however, insufficient Mn in the subsoil resulted in Mn deficiency in wheat.
Baranyai et al. (1987) showed that Mn soil concentration at 68 mg kg−1 was the optimum for maximum 9 t ha−1 yield of winter wheat (Triticum aestivum L.) under early phenological stages and that 1 mg kg−1 Mn increase in concentration would result in a reduction of approximately 65 kg ha−1 grain yield at harvest. Lasztity (2006) also found that Mn applied before seeding in nitrogen (0.2 g kg−1), phosphorous (20.1 g kg−1) and potassium (2.1 g kg−1) fertilizers (Kadar 1991) significantly increased Mn accumulation in shoot dry weight to 550 g ha−1 Mn, shoot dry weight and grain yield of winter wheat (Triticum aestivum L.), but Mn at flowering did not. Furthermore, Mn fertility of the surface soil horizon is important for vigorous crop growth at early stage to control weeds.
However, little attention has been paid to rhizosphere effects of crop plants at an agroecosystem level. Heenan and Chambel (1981) showed that Mn application in substrate at 3.1 μM was the optimum for maximum uptake in soybean [Glycine max (L.) Merr.] roots at early stage and that an increase in application would result in a reduction of approximately 1.7 μM in Mn uptake to 275 μM. Zhang et al. (2004) presented that in a Chainian rotation system of rice (Oryza sativa L.)–wheat (Triticum aestivum L) crops, Mn deficiency in wheat was caused by excessive Mn uptake by rice and Mn leaching from topsoil to subsoil due to periodic cycles of flooding and drying. Furthermore, wheat genotypes tolerant to Mn deficiency tended to distribute more roots to deeper soil layer and thus expanded their rhizosphere zones in the Mn-deficient soils and utilize Mn from the subsoil. Deep ploughing also helped root penetration into subsoil and was propitious to correcting Mn deficiency in wheat rotated with rice. In comparison, oilseed rape (Brassica napus L.) took up more Mn than did wheat through mobilizing sparingly soluble soil Mn, as a result of acidification and reduction processes in the rhizosphere. Thus, oilseed rape was tolerant to the Mn-deficient conditions in the rice–oilseed rape rotation. Masoni et al. (1996) presented that Mn-deficient rhizosphere of sunflower (Helianthus annuus L.) plant at early stage of growth in greenhouse conditions reduced leaf chlorophyll concentration, and subsequently, this reduction increased leaf reflectance and transmittance spectra, decreased leaf absorptance and shortened the red-edge position, defined as the inflection point that occurred in the rapid transition between red and near-infrared. Early physiological effects of developing Mn toxicity in young leaves of burley tobacco (Nicotiana tabacum L.) in glasshouse water-cultured plants grown at high (summer) and low (winter) photon flux Mn accumulation by leaves increased rapidly from 70 on day 0 to 1,700 and 5,000 μg per gram dry matter, and net photosynthesis declined by 20 and 60% after 1 and 9 days, respectively. Besides, Mn deficiency problems will decrease if Mn is supplemented at early growth stages. Manganese concentration in crop tissues decreases over time (Berczi et al. 1983; Heitholt et al. 2002; Alina and Henryk 2000). For Rice, the critical leaf Mn concentration ranged from 3.8 to 4.0 g kg−1 (Dobermann and Fairhurst 2000); for Clover, it ranged from 1.8 to 2.0 g kg−1 (Nable et al. 1984a, b). Accordingly, the Mn criterion of crop at the early stage should be considered as well as that at the minimum growth stage.
The objectives of this study were to evaluate some of the popular rotation crops grown in Hungary for tolerance to low external Mn levels and to determine the critical concentration of Mn deficiency during the early stages of growth under NPKCaMg-fertilization-induced field conditions in soil culture.
Materials and methods
Experimental site and general conditions
The study was conducted under NPKCaMg-induced field conditions on Experiment Station of Research Institute for Soil Science and Agricultural Chemistry of the Hungarian Academy of Sciences at Nyírlugos city (47°44′60″N, 22°8′80″E), Hungary (Fig. 1). The experiment station at Nyírlugos city (Fig. 1) inside of Debrecen and Nyíregyháza region is implemented in the so-called Northern Great Plain, which is located in the eastern-north-eastern part of Hungary, and it has been managed since 1962. This area is located 160 m above sea level. It is a typical lowland field with very poor mineral resources in the soil. There are no major differences in elevation within the region, but the climate is rather variable (Fig. 2). The local climate is somewhat dryer in the summer and a bit warmer in the winter than in the surrounding Hungarian Great Plain. The total number of sunny hours is 1,900–2,000 per year.

Location of the Experiment Station at Nyírlugos city

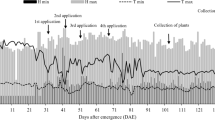
Precipitation variation in Nyírseg region
The minimum/maximum temperatures are about −25°C and +35°C. The annual mean temperature is 10–12°C. The area is very windy (SW and NE). It is one of the driest parts of Hungary (Marton 2005) with an annual precipitation of only 520–550 mm according to Kadar and Szemes (1994). The distribution of precipitation is rather uneven and unpredictable. The site is extremely drought sensitive. This is one of the major constraints, explaining why plant production is less successful. The water table level was found at a depth of 2–3 m.
Experiment soil, fertilizer treatments and management
The field experiment was set up on a sandy acidic lessivated brown forest soil, Haplic Luvisol in 1962. Soil texture had a particle-size distribution in plow-layer (0–25 cm) sand over 0.05 mm 70–85%, loam 0.05–0.002 mm 8–20%, clay under 0.002 mm 3–6%. Clay in colloid accumulation layers makes up to 10–18%. The saturation percentage was 25–30, pH (H2O) 5.4, pHKCl 4.3, organic matter 0.5–0.8%, CEC 3–5 meq 100 g−1.
The trial covered five fertilizers (nitrogen, phosphorus, potassium, calcium and magnesium) in 32 treatments, with four replications, and with a total 128 plots in a randomized block design. The detailed experimental codes, treatments, combinations and nutrient loadings are presented in Table 1a, b. Each plot had a 5 × 10 = 50 m2 plot size. The forms of fertilizers applied were Ca-ammonium nitrate, superphosphate, muriate of potash, powdered limestone and dolomite. The crop sequence of trial (Table 2) was potato–rye in the first 10 years (1963–1972), then potato–winter wheat (8 years; 1973–1980), then followed by different crops the next 10 years [white lupine (1981), wheat (1982), sunflower (1983–1984), grass (1985–1986), barley (1987), tobacco (1988), wheat (1989–1990)]. From 1991, from the 29th year of the trial, there is a triticale monoculture established, which is now 17 years old.
The crops used for this presentation were sunflower (Helianthus annuus L., cultivar: ‘HNK 81’) in 1983, tobacco (Nicotiana tabacum L., cultivar: ‘Hevesi 11’) in 1988 and triticale (x Triticosecale W., cultivar: ‘Presto’) in 1998, as a single year for each crop testing. The sunflower seeds were sown on April 10, 1983; tobacco on May 27, 1988; and triticale on October 03, 1997; and were allowed to grow in line of Lang (1976): sunflower 73 days, tobacco 50 days and triticale 191 days so to reach a phenological stage of sunflower 5–7 leaves, of tobacco 4–6 leaves and of triticale shooting, respectively. The triticale trial were only treated with herbicide (DMA-6; 2,4-D: 1 L ha−1) according to Ocsko et al. (1996) in 1998, and experiments were not treated with fungicides.
Soil sampling and analysis
Soil samples (0–20 cm depth; A-horizon) were collected using a 20-mm diameter core sampler in each cultivation cycle with sunflower on June 21, 1983; tobacco on August 11, 1988; and triticale on April 22, 1998. At each sampling, twenty soil cores per plot were randomly taken from each plot to make a composite sample. After thorough manual root separation, the soil samples of all plots were air-dried and sieved with a 2-mm mesh screen.
For measuring pHKCl, the suspension was made 1 M L−1 with respect to KCl and stirred. The pHKCl was measured after 24 h from a re-stirred suspension by MSZ 08-0206-2 method (Baranyai et al. 1987). Organic matter was determined by Tyurin method (Baranyai et al. 1987). In a 400-cm3 flask, 0.2–1.0 g of soil, 10 cm3 0.067 M L−1 K2Cr2O7 and about 20 cm3 H2SO4 were added. The mixture was kept for 5 min on a hot water bath, allowed to cool for 1 h, and 100 cm3 of water was added and was measured with Mohr’s salt, ferrous ammonium sulphate [FeSO4(NH4)2SO4*6H2O] titrimetrically.
Phosphorus and potassium were extracted with 0.14 N ammonium lactate + 0.4 N acetic acid (Egner et al. 1960) and determined by inductively coupled plasma spectrometry (ICP-AES ‘JY 238 Ultrace’) technics. CaCO3 equivalent of soil was determined by a method described by Trierweiler and Lindsay (1969). Magnesium was extracted with 100 cm3 1 M L−1 KCl, 1 h shaking, and was determined by ICP-AES ‘JY 238 Ultrace’ technics (Baranyai et al. 1987). For sunflower and tobacco, the soil Mnn+ content was extracted by 50 cm3 0.05 M L−1 Na2EDTA + 0.1 M L−1 KCl, 2 h shaking, and was assessed by ICP-AES ‘JY 238 Ultrace’ technics (Baranyai et al. 1987); for triticale, by 50 cm3 0.5 M L−1 NH4-acetate + 0.02 M L−1 EDTA, pH: 4.65, 1 h shaking (Lakanen and Erviö 1971) and was assessed by ICP-AES ‘JY 238 Ultrace’ technics.
Soil chemical properties
Soil analysis results showed the following main soil properties in case of sunflower in 1983: pHKCl from 4.0 to 5.7, organic matter from 0.4 to 0.7%, ammonium lactate–P2O5 from 51.0 to 222.0 mg kg−1, ammonium lactate–K2O from 94.0 to 144.0 mg kg−1, Ca from 155.0 to 260.0 mg kg−1, Mg from 20.0 to 53.0 mg kg−1 and Mnn+ from 29.0 to 74.0 mg kg−1; tobacco in 1988: pHKCl from 3.8 to 4.3, organic matter from 0.6 to 0.8%, ammonium lactate–P2O5 from 56.0 to 199.0 mg kg−1, ammonium lactate–K2O from 85.0 to 163.0 mg kg−1, Ca from 218.0 to 240.0 mg kg−1, Mg from 29.0 to 60.0 mg kg−1 and Mnn+ from 22.1 to 33.1 mg kg−1. Following are the corresponding results for triticale in 1998: pHKCl from 3.4 to 5.2, organic matter from 0.6 to 0.8%, ammonium lactate–P2O5 from 63.0 to 144.0 mg kg−1, ammonium lactate–K2O from 37.0 to 110.0 mg kg−1, Ca from 18.0 to 299.0 mg kg−1, Mg from 3.6 to 42.1 mg kg−1 and Mnn+ from 8.5 to 35.8 mg kg−1.
Plant sampling and analysis
Composite aboveground plant samples (LS) were taken from all the experimental plots at a phenological stage of 5–7 leaves for sunflower, 4–6 leaves for tobacco, shooting for triticale. The number of per-plot samples was 20–20 plants for sunflower (on June 21, 1983), 10–10 plants for tobacco (on July 15, 1988), 1–1 m2 for triticale (on May 12, 1998). Crop was cut and oven-dried at 65°C and weighted. Crop tissue Mnn+ concentration was determined using ICP-AES “JY 238 Ultrace” technics after microwave digestion using cc. HNO3 + cc. H2O2.
Crop Mn2+ properties
Plant analysis results showed that the crop Mn2+ properties were changed in case of sunflower from 62.0 to 263.0 mg kg−1, tobacco from 435.0 to 531.0 mg kg−1 and triticale from 186.0 to 398.0 mg kg−1.
Experimental soil Mn2+ data selection for treatments
From soil Mn2+ analysis results, the content of 42.5, 46.0, 48.0, 55.5, 65.0, 68.0 and 70.0 mg kg−1 (sunflower); 23.7, 24.3, 24.9, 27.2, 31.3, 32.1 and 33.7 mg kg−1 (tobacco); and 10.3, 17.0, 20.4, 28.7, 29.6, 32.2 and 32.7 mg kg−1 (triticale)(Mn-1, Mn-2…Mn-7) was selected and used as a Mn2+ “treatment” in each crop.
Statistical analysis
The results were analyzed using ANOVA and MANOVA, considering the treatment as the independent variable. All statistical analyses were carried out with the program SPSS 11.0 for Windows. All values are expressed as mean values. Significant statistical differences of all variables between the different treatments were established by the Tukey’s test at p < 0.05. A correlation matrix was calculated with the values of different parameters. The significance levels (p < 0.01 and p < 0.05) were based on Pearson coefficients.
Results
Crop growth
All crops were at juvenile stage at sampling. Growth of the tested crops stopped in the absence of Mn supply (Mn-1 treatment): sunflower 73 days, tobacco 50 days and triticale 191 days after sowing. All species would have Mn deficiency symptoms in the Mn-1 and Mn-2 treatments, namely yellowing leaves appearing in areas between veins, while the veins themselves remain green, and dies producing elongated holes in all crops. These symptoms can also be seen in the presence of excess iron, because this condition can block the uptake of manganese. In the Mn-1 treatment, only triticale showed a slight leaf yellowing. Leaf and shoot (LS) dry weight of all crops in the Mn-1 treatment was low(0.45–35.0 g plant−1), but the weight differed between crops in treatments from Mn-2 to Mn-7 (Fig. 3). LS dry weight increased with external Mn concentration, and all crops reached the optimum growth range in the treatments from Mn-2 to Mn-7. The LS dry weight of all crops decreased in the Mn-5 to Mn-7 (sunflower: Mn-5 to Mn-7, tobacco: Mn-6 to Mn-7 and triticale: Mn-5 to Mn-7) treatments. Average ABP dry weight of treatment for tobacco from Mn-3 to Mn-6, for sunflower and triticale from Mn-2 to Mn-6 (where crop growth remained in the optimum range) was used as a control (100% reference) to compare the growth response to external Mn concentration among the crops. Manganese treatments significantly (p < 0.01) affected relative LS dry weights. The relative LS dry weights were significantly (p < 0.01) different between crops, and the growth response to different Mn treatments varied significantly (p < 0.01) between crops. The latest significant difference at the 5% level (LSD0.05) between any pair of relative LS dry weights was 24%. Figure 4 shows the relative LS dry weight of the tested crops in the treatments from Mn-1 to Mn-7. Values in the Mn-1 treatment were omitted since the Mn concentration (x-axis) was represented by a logarithmic scale to express the results of the responses to low external Mn concentration more clearly. The relative LS dry weight of tobacco was about 60% or more in the Mn-2 treatment. The relative LS dry weight of triticale was less than 80% in the Mn-2 treatment but increased to about 85% in the Mn-3 treatment. The relative LS dry weight of sunflower was 90% or more in the Mn-1 and Mn-3 treatments.

Dry weight of three crops grown at different Mn concentrations is soil culture. Manganese concentrations were as follows: sunflower 4.25, 4.60, 4.80, 5.55, 6.50, 6.80 and 7.00 g kg−1; tobacco: 2.43, 2.37, 2.49, 2.72, 3.13, 3.21 and 3.37 g kg−1; triticale: 1.03, 1.70, 2.04, 2.87, 3.22, 2.96 and 3.27 g kg−1 (Mn-1, Mn-2…Mn-7) as Mnn+. Bars indicate standard errors of four replications

Relative LS dry weight of three crops grown at the Mn concentration from 1.03 to 7.00 g kg−1 in soil culture. The average weight of Mn-2, Mn-3, Mn-4, Mn-5 and Mn-6 (sunflower); Mn-3, Mn-4, Mn-5 and Mn-6 (tobacco); Mn-2, Mn-3, Mn-4, Mn-5 and Mn-6 (triticale) treatments was used as a control (100%) in each crop. The LSD0.05 of relative dry weight between any pair was 25% LS
Manganese concentration in crop tissue
Manganese concentration in leaf and shoot (LS) was significantly (p < 0.01) affected by both Mn treatment and crop species. The response of tissue Mn concentration to Mn treatment significantly (p < 0.01) differed between crops. The LSD0.05 among any pair of treatments was 0.30 g kg−1 in case of sunflower, 0.81 g kg−1 in case of triticale and 5.3 g kg−1 in case of tobacco. Manganese concentration increased with external Mn concentration from the Mn-1 to Mn-4 treatments, ranging from 6.20 to 53.1 g kg−1 in the LS dry matter (Fig. 5). The Mn concentration in all crop tissues greatly increased in Mn-2 treatment and remained almost constant up to Mn-2 treatment except for tobacco where the tissue Mn concentration continued to increase with external Mn concentration.

Manganese concentration in three crops grown at different Mn concentration in soil culture. The LSD0.05 among any pair of the treatments was 2.17% in LS. Bars indicate standard errors of four replications
The Mn concentration from the Mn-2 to Mn-4 treatments ranged from 6.20 to 53.1 g kg−1 in LS dry matter. Sunflower and triticale Mn concentration reached their maximum in Mn-6 and Mn-6 treatment, tobacco at Mn-4 and from the Mn-4 to Mn-7 treatments ranged from 6.20 to 53.10 g kg−1. The Mn concentration of sunflower and triticale was lower from Mn-1 to Mn-7 than of tobacco from Mn-1. Mn concentration was almost similar in the case of sunflower and triticale from Mn-4 to Mn-7 treatments and ranged from 6.20 to 39.8 g kg −1. The tobacco was an exception where Mn concentration of LS dry matter was higher in all treatments, and from the Mn-1 to Mn-7 treatments ranged from 43.5 to 53.10 g kg−1.
Discussion
External manganese concentration for optimum crop growth at early stage
Manganese concentration in crop tissue was much lower in Mn treatments below 1.02 g kg−1 (triticale), 2.43 g kg−1 (tobacco) and 4.25 g kg−1 (sunflower) than in the other treatments in all crops, including species that had large relative LS dry matter weight in the Mn-1 treatment, such as tobacco (Figs. 4, 5). Tissue Mn concentration and growth were highest in the Mn-4 treatment in all crops. The tissue concentration of crops increased with external Mn concentration, but LS dry weights of crops did not increase with Mn supply in nutrient solution, thus reflecting luxurious consumption of Mn. Consequently, the external Mn concentration to drive the optimum early growth for all crops was considered to be 2.87 g kg−1 in case of triticale, 2.72 g kg−1 in case of tobacco and 5.55 g kg−1 in case of sunflower in soil culture. This was consistent with the results of Jones and Leeper (1951) and Alina and Henryk (2000) who reported that the critical range of soil Mn extracted with EDTA was 2.0–5.0 g kg−1 in temperate crop fields. Demonstration of critical Mn concentration under greenhouse (closed system) conditions is difficult because of crop physiological Mn behavior that is not similar to natural field circumstances. The required external Mn concentration in soil culture of our study is in agreement with greenhouse-based determination of critical Mn levels. The critical soil Mn concentration for many field crops during the early growth period will likely range from 1.0 to 4.0 g kg−1 in highly leached soils. With this assumption, the rate of required Mn is equivalent to 22–43 kg ha−1 of single superphosphate complemented with MnSO4 (9.5% Mn) or 102–198 kg ha−1 of superphosphate (2% Mn), both of which are readily soluble.
Crop tolerance to low external manganese concentration
Since crops achieved optimum growth at 2.87 g kg−1 in case of triticale, 2.72 g kg−1 in case of tobacco and 5.55 g kg−1 in case of sunflower in soil nutrient solution, we considered the Mn treatments below 2.87 g kg−1 (triticale), 2.72 g kg−1 (tobacco) and 5.55 g kg−1 (sunflower) to correspond to low-Mn conditions. The relative dry weight of triticale was very low in both the Mn-2 and Mn-3 treatments (Fig. 4). The relative dry weight of sunflower was low in the Mn-1 treatment but increased in the Mn-2 treatment. Dry matter production of tobacco did not decrease in the Mn-2 and Mn-3 treatments. Thus, the minimum external Mn requirement for the acceptable growth was about 5.55 g kg−1 (sunflower), 2.87 g kg−1 (triticale) and 2.72 g kg−1 (tobacco). As a result, the tolerance of crops at early stages of growth to low external Mn concentration was in the order of triticale < sunflower < tobacco. Crop that has larger root growth at the lower Mn level, such as tobacco, would likely to be more tolerant to low Mn in the field because of the ability to explore more of the soil profile. Furthermore, crops that have a rapid root elongation rate might have higher tolerance in fields because they would be quicker to explore lower parts of the Mn-accumulated soil profile. Medium- and high-tolerant crops grew with low tissue Mn concentration, but triticale, which is susceptible to low external Mn, did not (Figs. 4, 5). Roots of high- and medium-tolerant crops are likely to have higher level of Mn in tissues and thus act as a buffer to changes in Mn availability in external conditions. In general, root Mn concentration is lower than the shoot Mn concentration in low external Mn condition, and the reverse is the case under high external Mn conditions. Tropical crop species such as sunflower, to warm arid climates, where soils have high inherent potential to supply Mn due to solid-phase manganese, were less tolerant to low external Mn conditions and required higher Mn concentrations for optimum growth (Fig. 4). Tobacco grew better compared with sunflower at lower Mn levels.
Critical manganese concentration in crop leaf and shoot
Figure 6 represents the relationship between LS concentration and relative LS dry weight obtained from the mean values of all replications. The relative LS dry weight increased with increased dry Mn concentration, and optimum growth was achieved for all crops within the range of LS dry matter Mn concentrations evaluated. Critical growth to diagnose plant nutrition status has been suggested at levels of 95% (Fox 1976), 90% or 50% of maximum growth.

Schematic relationship between Mn concentration in crop LS relative and relative LS dry weight of three crops obtained from mean values. The average weight of Mn-2, Mn-3, Mn-4, Mn-5 and Mn-6 (sunflower); Mn-3, Mn-4, Mn-5 and Mn-6 (tobacco); Mn-2, Mn-3, Mn-4, Mn-5 and Mn-6 (triticale) treatments was used as a control (100%) in each crop. The LSD0.05 of Mn concentration between any pair was 45.7 mg kg−1 and that of relative LS dry weight was 25%. Bars indicate standard errors of the four replications for LS Mn concentration and those of four replications for relative LS dry weight. Critical dry matter Mn concentration was reported to be from 1.0 to 2.0 g kg−1 (Marschner 1995), from 2.0 to 3.0 g kg−1 (Champbell 2000), from 2.0 to 5.0 g kg−1 (Alina and Henryk 2000) and from 17.1 to 144.0 g kg−1 (Kamprath and Foy 1985) for crops. Our results are in agreement with the published reports
In the present study, the critical growth of the juvenile crops was assumed to be 75% of relative LS dry matter weight. These levels were chosen because such degree of early growth would depress subsequent growth and grain yield though not as high as that suggested by Fox (1976) because juvenile crops would have more time to recover from the stress. Critical Mn concentration for 75% relative LS growth was significantly (p < 0.01) different between the tested crops (3.0 mg kg−1 LSD0.05) (Table 3; Fig. 7). The critical Mn concentration of crops was grouped for comparison with the mean value by t test (5% level). The critical Mn concentration was low (5.09–5.63 g kg−1) in sunflower; medium (18.41–20.34 g kg−1) in triticale; and high (43.51–48.09 g kg−1) in tobacco.

Characteristics of Mn absorption in three crops at early stages differing in tolerance to low external Mn concentration. Plants were grown for 73 days (sunflower), for 50 days (tobacco) and for 191 days (triticale) in soil culture. The average weight of Mn-2, Mn-3, Mn-4, Mn-5 and Mn-6 (sunflower and triticale) and Mn-3, Mn-4, Mn-5 and Mn-6 (tobacco) treatments was used as a control (100%) in crops. The absorbed Mn in LS of the control plants was calculated. Bars indicate standard errors with the four replications
Characteristics of manganese absorption by crops
The total Mn uptake in the plant is the product of the Mn concentration and dry weight. The amount of Mn absorbed was dominated by the crop growth rate and differed between the tested crops (Table 3; Fig. 7).
The value was relatively small in triticale compared with sunflower and tobacco. Although the control LS dry weight of sunflower was intermediate among the tested crops, the amount of Mn absorbed was intermediate as well due to tissue Mn concentration. Crop tolerance to low external Mn concentration did not relate to the critical Mn concentration in crop LS nor to the absorbed amount of Mn for optimum growth. Even when the critical Mn concentration for deficiency was similar, the absorbed amount of Mn for the optimum growth largely differed between crops. Since there is a large variation in the Mn content among different soil layers, it is difficult to diagnose soil Mn fertility through routine soil analysis.
Manganese fertility in the upper soil layers, which controls early crop growth, can be estimated based on the critical LS Mn concentration of crops at early stages. Once growth at early stages is established, one can expect that the developed roots will exploit the Mn accumulated in deeper soil layers. Manganese supplement to crops at early stages, thus, should be considered to provide adequate Mn supplement throughout the growth cycle. Crop tolerance to low external Mn concentration.
References
Academic Press Geology Series (IPCS). (1981). Manganese (Environmental Health criteria 17). Geneva: Sweden World Health Organization. International Programme on Chemical Safety.
Adriano, D. C. (1986). Trace elements in the terrestrial environment. New York: Springer.
Alina, K. P., & Henryk, P. (2000). Trace elements in soils and plants (3rd ed.). Boca Raton: CRC Press.
Aubert, H., & Pinta, M. (1977). Trace elements in soils. Amsterdam: Elsevier.
Baranyai, F., Fekete, A., & Kovacs, I. (1987). Results of Hungarian soil analysis (In Hungarian). Budapest: Mezőgazdasági Kiadó.
Berczi, A., Olah, Z., & Erdei, L. (1983). Nutrition of winter wheat during the life cycle. I. Yield and accumulation of dry matter, and minerals. Plant Physiology, 58, 124–130.
Centre for Ecology and Hydrology. (1983). Applied environmental geochemistry. London: CEH.
Champbell, C. R. (2000). Reference sufficiency ranges for plant analysis in the southern region of the United States. Raleigh, NC, USA: Agronomic Division of the North Carolina, Department of Agriculture and Consumer Services.
David, S., & Philippe, B. (1995). Europe’s environment. European environment agency. Copenhagen, Denmark.
Dobermann, A., & Fairhurst, T. (2000). Rice nutrient disorders and nutrient management. Handbook series. Los Banos, Laguna, Philippines: Potash and Phosphate Institute (PPI), Potash and Phosphate Institute of Canada (PPIC) and International Rice Research Institute (IRRI).
Egner, H., Riehm, H., & Domingo, W. R. (1960). Untersuchungen über die chemische Bodenanalyse als Grundlage für die Beurteilung des Nahrstoffzustandes der Böden. II. K. Lantbrhögsk. Ann, 26, 199.
Feher, Bne. (2005). Sandy soil properties for asparagus crop production. Seed Researches, Development and Environment, 12, 13–16.
Fox, M. R. S. (1976). In A. S. Prasad (Ed.), Trace elements in human health and disease (Vol. II, pp. 401–416). New York: Academic Press.
Hadimani, A. S., Surya, P. R., Parvathappa, H. C., & Raghavendra, K. R. (1974). Effect of water soluble organic substances from different types of leaf and needle litter on the downward movement of manganese in sandy soil. Plant and Soil, 40, 365–372.
Heenan, D. P., & Champbell, L. C. (1981). Manganese and iron interactions on their uptake and distribution in soybean (Glycine max (L.) Merr.). Plant and Soil, 70, 317–326.
Heitholt, J. J., Sloan, J. J., & MacKown, C. T. (2002). Copper, manganese, and zinc fertilization effects on growth of soybean on a calcareous soil. Journal of Plant Nutrition, 25, 1727–1740.
Howe, P. D., Malcolm, H. M., & Dobson, S. (2004). Manganese and its compounds: Environmental aspects. UK: Monks Wood.
Jones, L. H. P., & Leeper, G. W. (1951). Available manganese oxides in neutral and alkaline soils. Plant and Soil, 3, 154–159.
Kadar, I. (1991). Soil and plant heavy metal analysis. In RISSAC (Ed.), Research of environmental protection (p. 104). Budapest, Hungary: EM-RISSAC.
Kadar, I., & Szemes, I. (1994). Lessons learned from a 30 year old field trial in Hungary. Budapest: RISSAC-HAS.
Kamprath, E. J., & Foy, C. D. (1985). Lime-fertilizer-plant interactions in acid soils. In O. Englestad (Ed.), Fertilizer technology and use (3rd ed.). Madison, Wisconsin, USA: Soil Science Society of America.
Lakanen, E., & Erviö, R. (1971). A comparison of eight extractants for the determination of plant available micronutrients in soil. Acta Agraria Fennica, 123, 223–232.
Lang, G. (1976). Field crop production. Budapest: Agricultural Press.
Lasztity, B. (2006). Mineral nutrient accumulation in cereals. Budapest, Hungary: BME Press.
Marschner, H. (1995). Mineral nutrition of higher plants. London, UK: Academic Press.
Marton, L. (2005). Disasters as drought-, and rainfall excess and artificial fertilization effects on crop yield. Charlotte, USA. In Proceedings of the international conference on energy, environment and disasters-INCEED2005 (pp. 49–50). Charlotte: ISEG, July 24–30, 2005.
Masoni, A., Ercoli, L., & Mariotti, M. (1996). Spectral properties of leaves deficient in iron, sulfur, magnesium, and manganese. Agronomy Journal, 88, 937–943.
Nable, R. O., Bar, A. A., & Loneragen, J. F. (1984a). Functional manganese requirement and its use as a critical value for diagnosis of manganese deficiency in subterranean clover (Trifolium subterraneum L. cv. Seaton Prk). Annals of Botany, 54, 39–49.
Nable, R. O., Houtz, R. L., & Cheniae, G. M. (1984b). Early inhibition of photosynthesis during development of Mn toxicity in tobacco. Plant Physiology, 86, 1136–1142.
Ocsko, Z., Molnar, J., & Eke, I. (1996). Agrochemicals. Budapest: MRD.
Shihua, L., Xuejun, L., Long, L., Fusuo, Z., Xiangzhong, Z., & Caixian, T. (2004). Effects of manganese spatial distribution in the soil profile on wheat growth in rice-wheat rotation. Plant and Soil, 261, 39–46.
Sillanpää, M. (1982). Micronutrients and the nutrient status of soils: A global study (Vol. 48). Rome: FAO Soils Bulletin.
Singh, B. R. (1974). Migration of ions in soils II. Movement of Mn54 from surface application of manganese sulphate in soil columns. Plant and Soil, 41, 629–636.
Trierweiler, J. F., & Lindsay, W. L. (1969). EDTA-ammonium carbonate soil test for zink. Soil Science Society of America Proceedings, 33, 49–54.
Uren, N. C. (1990). The movement and distribution of manganese added to soil. Australian Journal of Soil Research, 28, 677–683.
Wong, M. H., Wong, J. W. C., & Baker, A. J. M. (1998). Remediation and management of degraded lands. Boca Raton: Lewis Publishers.
Zhang, F., Shen, J., Li, L., & Liu, X. (2004). An overview of rhizosphere processes related with plant nutrition in major cropping system in China. Plant and Soil, 260, 89–99.
Acknowledgmets
We express our gratitude to Dr. Kiyoko Hitsuda of Crop Production and Environment Division of Japan International Research Centre for Agricultural Science (JIRCAS), Japan, for their previous suggestions, discussions and comments. We also acknowledge the help from Dr. Láng István and Dr. Kádár Imre of the Hungarian Academy of Sciences, Hungary; and the Hungarian and Spanish Intergovernmental S & T Cooperation Project of E-2/04-OMFB-00112/2005; and Hungarian and Indian Intergovernmental S & T Cooperation Project of IND-3/03/2006-OMFB-00295/2006 for experimental and scientific support.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Marton, L. Crop demand of manganese. Environ Geochem Health 34 (Suppl 1), 123–134 (2012). https://doi.org/10.1007/s10653-011-9405-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10653-011-9405-3