
Abstract
Even trace amounts of cadmium (Cd), a non-essential metal, are known to be toxic to aquatic organisms. Here we investigated the relationship between cadmium ion (Cd2+) exposure and oxidative damage and apoptosis in the hepatopancreas of the clam Meretrix meretrix. Clams were exposed to different concentrations of Cd2+ (0, 1.5, 3, 6 and 12 mg L−1) for 5 days. We monitored both antioxidant enzyme activity, including that of superoxide dismutase (SOD), catalase (CAT), glutathione peroxidases (GPx), and levels of malondialdehyde (MDA), glutathione (GSH) and glutathione disulfide (GSSG). Apoptosis of hepatopancreatic cells was detected by DNA laddering and AO/EB double fluorescent staining. The results show that the rate of apoptotis, MDA levels, and caspase-3 activity, increased with Cd2+ concentration, whereas GPx activity and the ratio of GSH/GSSG, decreased. SOD and CAT enzyme activity first increased, then decreased, with increasing Cd2+ concentration; peak activity of these enzymes was recorded in the 3 mg L−1 Cd2+-treatment group. These results show that Cd-induced oxidative damage can both induce, and aggravate, apoptosis in the hepatopancreatic cells of clams, even at Cd2+ concentrations far below the semi-lethal dose for adult clams. The observed changes in caspase-3 activity enhanced significantly at lower Cd2+ concentrations, indicating that caspase-3 is a suitable biomarker for heavy metal pollution, especially cadmium pollution, in marine organisms.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Heavy metal pollution in aquatic ecosystems probably has long-lasting effects, even in marine ecosystems. Cadmium (Cd) is a widespread heavy metal in the marine environment that poses a serious risk for human health when it enters and becomes bio-magnified in food webs (Wagner et al. 1998; Waalkes 2000; Wang et al. 2008). Cd is a non-essential metal that can cause a series of toxicological changes in organisms (Geret et al. 2002; Ketata et al. 2007; Ji et al. 2012). As peroxide inducers, cadmium and other heavy metal contaminants can stimulate the production of excess free radicals.
Oxidative stress occurs when the generation of reactive oxygen species (ROS) exceeds an organism’s capacity to neutralize these molecules (Wang et al. 2011; Cao et al. 2012). Excess free radicals have a range of deleterious effects, including causing lipid peroxidation and blocking the mitochondrial electron transport chain (Bolduc et al. 2004; Shih et al. 2004). These effects can cause DNA damage and other abnormalities leading to cellular damage and apoptosis (Nzengue et al. 2008; Wang et al. 2011; Ali et al. 2012; Ji et al. 2012; Zhang et al. 2012). MDA has recently been suggested as a potential biomarker of oxidative stress (Otitoloju and Olagoke 2011) in which case quantification of MDA could allow oxidative stress to be detected before lethal, or pathological, effects occur (Wang et al. 2013; Zhang et al. 2014). Caspase-3, an effector caspase, has been shown to play a critical role in apoptosis (Shih et al. 2004; Liu et al. 2013).
Organisms have evolved effective anti-ROS defense systems, mainly comprised of low molecular weight compounds, such as vitamin A, C, E, glutathione (GSH), uric acid, and major antioxidant enzymes, such as catalase (CAT), superoxide dismutase (SOD), glutathione peroxidase (GPx), glutathione transferase (GST) and others (Chandran et al. 2005; Ivanina et al. 2008; Loro et al. 2012; Zhang et al. 2014). These anti-ROS systems work by scavenging free radicals, thereby reducing oxidative stress and inhibiting apoptosis (Angkeow et al. 2002; Wang et al. 2013). The potential of using elements of the antioxidant defense system as biochemical biomarkers for environmental monitoring made it the focus of intensive research (Otitoloju and Olagoke 2011).
Bivalve mollusks inhabiting intertidal zone sediments are widely distributed globally. Because these invertebrates are suspension feeders with poor locomotor ability, they are particularly vulnerable to heavy metal contamination. For this reason, bivalves (mussels, oysters, clams, etc.) have been widely used in the biological monitoring of marine heavy metal pollution (Byrne and O’Halloran 2001; Hamad et al. 2011; Meng et al. 2013).
Meretrix meretrix (Bivalvia, Veneridae) is an economically important shellfish that lives in the sediments of the neritic intertidal zone, including estuaries, along the coast of China. Several studies have shown that heavy metals, such as zinc (Zn2+) and cadmium (Cd2+), have serious toxic effects on the Veneridae, including retarding the development of both gonads and larvae (Wang et al. 2009a, b; Zhang et al. 2011). For example, exposure to these heavy metals is followed by a dramatic decrease in GSH levels and GPx activity in the gonads, membrane lipid peroxidation (Zhang et al. 2011), and a significant increase in antioxidant enzyme activity in M. meretrix larvae (Wang et al. 2010). The effects of Cd2+ exposure on the antioxidant system of adult M. meretrix and hepatopancreatic cells are, however, currently unknown.
The present study makes use of several biochemical parameters, including DNA fragmentation, antioxidants (SOD, CAT, and GPx), the key enzymes of apoptosis (caspase-3), and the level of GSH, GSSG and MDA in the M. meretrix exposed to different concentrations of Cd2+. We particularly focused on the relationships between Cd2+ exposure, antioxidant defenses, oxidative damages and apoptosis in the hepatopancreatic cells of M. Meretrix. The main objective of this study was to explore the toxic effect of Cd2+ on the clam, and to improve our understanding of the antioxidant defense mechanisms of clam against Cd2+ stress. A secondary goal was to provide baseline information of biomarkers’ selection for the assessment of Cd2+ pollution in aquatic ecosystems.
Materials and methods
Reagents
Analytical grade cadmium chloride (CdCl2·2.5 H2O) was purchased from Tianlian Fine-Chemical Co., ltd. (Shanghai, China). A DNA extraction assay kit was obtained from Tian Gen Biotech Co., Ltd (Beijing, China). Caspase-3, GSSG/GSH assay kits were acquired from Beyotime Institute of Biotechnology (Shanghai, China). MDA, SOD, GPx, and CAT assay kits were obtained from Nanjing Jiancheng Bioengineering Institute (Nanjing, China). AO/EB double fluorescent stain was purchased from Nanjing Keygen Biotech Co., Ltd (Nanjing, China).
Animals and treatments
Meretrix meretrix clams which was 2 years old were purchased from the Linkun aquafarm, Wenzhou, Zhejiang, China, between June and July 2013. The Cd concentration levels of total fraction in seawater and deposit sediment of the clam farm were 0.54 μg L−1 and 0.89 mg kg−1, respectively. Clams were acclimated in 15 % artificial sea water without food for 2 days in auto-temperature control aquarium at a temperature of 22 ± 1 °C before beginning experiments. The sea water was renewed, and the aquarium thoroughly cleaned, daily. Any dead clams were immediately removed. After acclimation, 300 healthy clams (see Table 1 for size and weight) were selected for a semi-lethal Cd2+ concentration (LC50) test. The LC50 toxicity experiment was conducted using a modified method outlined by Wang (Wang et al. 2009a, b). The 48 h LC50 and 96 h LC50 Cd2+ levels were 40.78 and 15.01 mg L−1, respectively.
Clams were then randomly assigned to either a control group, or one of four Cd2+-treatment groups, each comprised of 30 clams. The control and treatment groups each had three replicates so the experiment involved a total of 450 clams, accommodated in 15 aquariums. The four treatment groups were exposed to 1.5, 3, 6 or 12 mg L−1 concentrations of cadmium solution, corresponding to 1/10, 1/5, 1/2.5, 1/1.25 of the previously determined 96 h LC50 (15.01 mg L−1). To maintain both good water quality and heavy metal absorption, the different concentrations of cadmium solution were renewed daily throughout the experiment. All other experimental conditions were kept the same as during acclimation. No mortality occurred in any treatment group. All clams were collected and dissected after 5 days of cadmium exposure, after which the hepatopancreas of each animal was quickly removed and immediately frozen in liquid nitrogen before being stored at −80 °C in a refrigerator until required for analysis. Basic parameters of the clams used in the LC50 test and acute toxicity experiments are listed in Table 1.
Cadmium content determination
Concentrations of Cd were measured following the Chinese National Standard for determination of cadmium in food (GB1738.6-2007) with some modifications. After exposure, hepatopancreas tissues were excised and weighted (approximate 2 g wet weight), cut into small pieces, and excess water on surface of tissues was removed with absorbent paper. Then tissues were digested in HNO3 (analytical grade) and HClO4 (analytical grade) over a hot plate at about 120–150 °C, under a reflux cap. Cd concentrations of each sample were measured with an atomic absorption spectrophotometer (Shimadzu AA-6300, Japan). Standard Cd solution was used for the analysis of metal concentrations. The carrier gas was acetylene. Cd content was expressed as mg kg−1 wet weight tissue.
Detection of apoptosis with fluorescent microscopy
Following the methods described by Ribble et al. (2005) and Kasibhatla et al. (2006), acridine orange/ethidium bromide (AO/EB) staining of hepatopancreatic cells was carried out to estimate the degree of cell damage induced by Cd2+. 60 mg of hepatopacreatic tissue was prepared as a cell suspension in precooling 1 % PBS (pH = 7.2). A 25 μL suspension of each sample was mixed with 1 μl aqueous AO/EB solution (100 μg ml−1 of AO in PBS; 100 μg ml−1 of EB in PBS) and incubated for 5 min before being examined under fluorescent microscopy (Nikon Ti-s, Japan).
Because AO penetrates intact cell membranes and stains nuclear DNA with bright green fluorescence, whereas EB penetrates damaged cell membranes and stains nuclear DNA with orange fluorescence, fluorescent microscopy allows to distinguish different types of cells. Viable cells had bright green nuclei, whereas early-stage apoptotic cells were yellow with nuclear chromatin condensation, and late-stage apoptotic cells and dead cells had orange or red nuclei with intact structures. A minimum of 100 cells were counted at least 3 times from each treatment group. Data from the AO/EB test were expressed as the percentage of apoptotic cells.
Detection of apoptosis by DNA laddering
50 mg of hepatopancreatic tissue from the treatment and control groups was added to separate vessels containing 500 μL physiological saline, thoroughly ground into a suspension on ice and centrifuged at 11,200 g (4 °C) for 1 min. After centrifugation, the supernatant was removed and the sediment used for DNA fragment extraction. DNA was extracted with a DNA extraction kit (Beyotime, C008) according to the manufacturer’s instructions. A DNA sample mixture (8 μL DNA extract, 3 μL Bromophenol blue) was added to each lane of a 1 % agarose gel (containing 0.1 mg ml−1 ethidium bromide) in 0.5 × TBE buffer and subject to electrophoresis at 58 V for 1 h. The resultant bands were photographed under ultraviolet light with Gel Doc 2000 (Tannon, Shanghai, China).
Caspase-3 activity assay
Caspase-3 activity was measured with a corresponding caspase-3 Activity Assay Kit (Beyotime, Shanghai, China). Nine hepatopancreatic tissue samples from each treatment group were randomly collected, weighted and homogenized on ice (5 % w/v) in lysate with a SCIENTZ DY89-II type motor-driven homogenizer (Zhejiang, China). After homogenization, samples were incubated for 5 min on ice, then centrifuged at 18,000 g for 20 min at 4 °C. The supernatant was collected for caspase-3 activity measurements according to the kit manufacturer’s instructions. One unit of enzymatic activity was taken to be the amount of enzyme required to cleave 1.0 nmol of the colorimetric substrate to produce 1.0 nmol pNA per hour at 37 °C under saturated substrate concentrations. Optical density values were read at 405 nm on a multifunction microplate detector (Biotek Epoch, Vermont, USA).
Antioxidant enzyme activity assay
Hepatopancreas samples were macerated on ice to form a 10 % (w/v) homogenate in 0.9 % physiological saline using a SCIENTZ DY89-II type motor-driven homogenizer. Supernatants were collected after centrifugation at 3500 rpm for 15 min at 4 °C for SOD, GPx and CAT activity measurement. All enzyme activity was assayed according to the kit manufacturer’s instructions (Nanjing Jiancheng, China) and defined as units of activity per mg of protein (U mg−1).
MDA, GSH and GSSG assay
Tissue MDA content, a biomarker of lipid peroxidation, was determined with a colorimetric MDA assay kit (Nanjing Jiancheng, China) according to the manufacturer’s instructions. GSH and GSSG in the hepatopancreas were measured using GSH and GSSG Assay Kits (Beyotime, China) based on the method described by Tietze (1969), according to the kit manufacturer’s instructions. GSH and GSSG levels were expressed in mg per g protein (mg g−1), and MDA levels as nmol per mg protein (nmol mg−1).
Total protein assay
The total protein content of the hepatopancreas was measured according to the method described by Bradford (1976) using bovine serum albumin as a standard.
Statistical analysis
The data were analyzed using the SPSS Statistical Package (Version 16.0, Chicago, USA for Windows). The significance of differences among experimental groups was determined using a one-way ANOVA followed by Tukey’s Multiple Comparison Test. P < 0.05 was taken to be statistically significant, and P < 0.01 was considered highly significant. The Least Significant Difference test (LSD) was used to perform multiple comparisons among different treatment groups. All data are expressed as mean ± standard error (SE).
Results
Cd bio-accumulation in hepatopancreas of M. meretrix
The original pre-exposure Cd concentration levels of the hepatopancreas of M. meretrix was 0.29 ± 0.01 mg kg−1 wet tissue. After Cd exposure for 5 days, Cd levels of the hepatopancreas increased sharply, and reached maximum levels in the 12 mg L−1 exposure groups (Table 2). There was significantly different among control group and all Cd2+-treatment groups (P < 0.05). And Cd concentrations in the hepatopancreas of M. Meretrix showed a higher bio-accumulation and concentration-dependent manner.
Morphological observation of apoptosis with fluorescent microscope
Three types of cells could be distinguished under fluorescent microscopy after AO/EB staining; live cells (green), early-stage apoptotic cells (yellow), late-stage apoptotic (orange), and dead cells (red) (Fig. 1a–e). The control group had almost all normal cells (green) indicating no apoptosis (Fig. 1a). As the Cd2+ concentration increased, both the number of apoptotic cells and the number of late-stage apoptotic cells, significantly increased (Fig. 1b–e). At the highest concentration (12 mg L−1) (Fig. 1e), almost all cells were in the late stage of apoptosis (orange), and some were already dead.

Results of AO/EB double fluorescent staining of M. meretrix hepatopancreatic cells exposed to different Cd2+ concentrations; a control group (0 mg L−1 Cd2+); b 1.5 mg L−1 Cd2+; c 3.0 mg L−1 Cd2+; d 6.0 mg L−1 Cd2+; e 12 mg L−1 Cd2+. Green indicates healthy cells whereas yellow, red or orange indicate different stages of apoptosis (Color figure online)
We calculated the rate of apoptosis in each group according to the following formula: apoptosis ratio = (early-stage apoptotic cells + late-stage apoptotic cells)/total cells × 100 %. The result showed that the apoptosis ratio increased with the Cd2+ concentration, and that the rate in each Cd2+-treatment group was significantly different to that in the control group (P < 0.05) (Table 3). The apoptosis ratio in the 12 mg L−1 Cd2+-treatment group reached 90.75 %, whereas that in the control group was just 3.89%. This indicated that the apoptosis ratio was positively correlated with the Cd2+ concentration.
DNA fragmentation analysis by agarose gel electrophoresis
Agarose gel electrophoresis was used to detect DNA degradation in hepatopancreatic cells (Fig. 2, lanes M–E). With the exception of two bright DNA bands in the low-molecular area near the loading slot, no obvious DNA laddering was detected in the control group (Fig. 2; lane E). In contrast, DNA ladders were apparent in all Cd2+-treatment groups (Fig. 2, lanes A–D). The distances between each ladder were nearly 200 bps, a metric which is indicative of apoptosis. The degree of apoptosis increased with Cd2+ concentration, and maximal DNA fragmentation occurred in the 12 mg L−1 Cd2+-treatment group, which suggests that Cd2+ induced apoptosis in a dose-dependent fashion.

Laddering of M. meretrix hepatopancreatic cell DNA induced by exposure to different Cd2+ concentrations. Lane labels are: M marker, A 12 mg L−1 Cd2+, B 6 mg L−1 Cd2+, C 3 mg L−1 Cd2+, D 1.5 mg L−1 Cd2+, E control group
GSH and GSSG content
Figure 3 shows the trend in GSH and GSSG levels in hepatopancreatic cells under different Cd2+ concentrations. GSH levels in the 3 and 6 mg L−1 Cd2+-treatment groups were significantly lower than in the control group (P < 0.05), and the GSH level in the 12 mg L−1 Cd2+-treatment group was extremely low (0.31 ± 0.01 mg g−1) compared to the control (P < 0.01). In contrast, with the exception of the 12 mg L−1 Cd2+-treatment group which was significantly different to all other groups (P < 0.05), no significant differences in GSSG levels were found among the groups.

GSH and GSSG levels, and GSH/GSSG ratio, in the hepatopancreas of M. meretrix induced by exposure to different concentrations of Cd2+. Note Significant differences between groups are indicated by different letters above. Bars (P < 0.05)
The ratio of GSH/GSSG declined with Cd2+ concentration (Fig. 3). The lowest ratio was recorded in the 12 mg L−1 Cd2+-treatment group, which was significantly different to all other groups (P < 0.01). There was no significant difference in GSH/GSSG ratio between the 1.5 mg L−1 Cd2+-treatment group and the control group (Fig. 3).
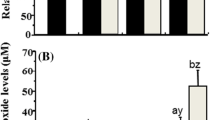
MDA content
MDA levels increased with Cd2+ concentration reaching the highest recorded level of 2.94 ± 0.20 nmol mg−1 protein in the 12 mg L−1 Cd2+-treatment group; significantly higher than in the control group and all other Cd2+-treatment groups. There was no significant difference in MDA levels of the control, the 1.5 mg L−1 treatment group, or the 3 mg L−1 Cd2+-treatment group (Fig. 4).

MDA levels in the hepatopancreas of M. meretrix induced by exposure to different concentrations of Cd2+
Caspase-3 enzyme activity
Caspase-3 activity increased sharply with Cd2+ concentration (Fig. 5) and was significantly higher in all Cd2+-treatment groups than in the control (P < 0.01).

Caspase-3 activity in the hepatopancreas of M. meretrix induced by exposure to different Cd2+concentrations
Antioxidant enzyme activity
Cellular defenses were taken against oxyradicals including enzymes, particularly superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidases (GPx). GPx activity was lowest in the 12 mg L−1 Cd2+-treatment group (15.20 ± 1.0 U mg−1 protein) (P < 0.01), and was significantly lower in the 3 and 6 mg L−1 Cd2+-treatment groups compared to the control group (Fig. 6). GPx activity in the 1.5 mg L−1 Cd2+-treatment group did not differ significantly from that in the control group (Fig. 6).

Effects of exposure to different concentrations of Cd2+ on GPx, SOD, and CAT activity in the hepatopancreas of M. meretrix
The relationship between SOD activity and Cd2+ concentration was not linear. SOD activity in the 3 mg L−1 Cd2+-treatment group (36.51 ± 1.27 U mg−1 protein) was significantly higher than in all other treatment groups (P < 0.05) except the 6 mg L−1 Cd2+-treatment group (Fig. 6). SOD activity in the 12 mg L−1-treatment group (28.08 ± 3.10 U mg−1 protein) was, however, lower than in all other treatment groups (Fig. 6; P < 0.05).
The relationship between CAT activity and Cd2+ concentration was similar to that between SOD and Cd2+ concentration (Fig. 6). CAT activity in the 3 mg L−1 Cd2+-treatment group (2.95 ± 0.13 U mg−1 protein) was significantly higher than that in the control group and the 12 mg L−1 Cd2+-treatment group (Fig. 6; P < 0.05). There was no significant difference in the CAT activity of the 1.5, 3, and 6 mg L−1 Cd2+-treatment groups, and CAT activity in the 12 mg L−1 Cd2+-treatment group was similar to that in the control group (1.75 ± 0.21 U mg−1 protein) (P > 0.05) and significantly lower than in all other treatment groups (Fig. 6; P < 0.05).
Discussion
Previous research has shown that the distribution and accumulation of heavy metal pollutants in aquatic systems can differ between different aquatic organisms and between different organs in the same organism (Silvestre et al. 2005; Rainbow and Black 2007; Zhao et al. 2012). Waterborne heavy metals are taken in via the gills and intestines by aquatic animals. Then they are transported to other parts of the body, commonly by the circulatory system if this is present. Digestive gland (hepatopancreas) is one of the most vulnerable and sensitive organs to heavy metal pollutants because of the function of metal homoeostasis, excretion and detoxification (Orbea et al. 2000; Yan et al. 2007; Liu et al. 2008).
Therefore, the bio-accumulation of Cd in hepatopancreas of M. meretrix was investigated in this study. The Cd concentration level of total fraction in seawater of the clam farm and the hepatopancreas of M. Meretrix were 0.54 μg L−1 and 0.29 mg kg−1, respectively. When two years’ old M. meretrix was exposed to different concentrations of Cd for 5 days, the concentration of Cd in hepatopancreas increased significantly (Table 2). The bio-accumulation of Cd was in a concentration-dependent manner. And the maximum values of accumulation in 12 mg L−1 treatment groups reached 37.90 ± 0.73 mg kg−1, which was 130.69 fold than pre-exposure group. This observation supported the result of Li (Li et al. 2015), which demonstrated that exposure to Cd led to a significant accumulation of Cd in the hepatopancreas of Sinopotamon henanense in a time and concentration-dependent manner. The accumulation of Cd2+ in hepatopancreas of M. meretrix would induce apoptosis and oxidative damage. The number of apoptotic and dead cells was significantly increased with increasing Cd2+ concentration (Table 3; Figs. 1, 2), which is in accordance with the previous study of the effects of Cd2+ on the freshwater crab S. henanense (Liu et al. 2013). Apoptosis was associated with increased DNA fragmentation, which also increased with Cd2+ concentration (Xiang et al. 2001). That Cd2+ caused DNA damage is evident from the fact that clams exposed to higher Cd2+ concentrations had significantly more DNA fragmentation than those exposed to lower concentrations (Fig. 2, lanes A–C). Taken together, the data presented in Figs. 1, 2 and Table 3 demonstrate that Cd2+ induced apoptosis of hepatopancreatic cells in M. meretrix in a dose-dependent fashion. This is consistent with the findings of other studies. For example, Wätjen et al. (2002) found that Cd2+ induced apoptosis of C6 glioma cells in a both time and concentration-dependent way.
Caspase-3 activity also increased with increasing Cd2+ concentration (Fig. 5). Caspases are a family of cysteine-dependent aspartate directed proteases that play a critical role in the initiation and execution of apoptosis (Hagara and Malki 2014). Our results showed that exposure to Cd2+ increased the activity of caspase-3 in the hepatopancreas of M. Meretrix. The activation of the caspase family of proteases is consistent with the hypothesis that cadmium exposure causes DNA damage leading to apoptosis (Shih et al. 2004; Liu et al. 2013). However, in contrast, Liu et al. (2013) found that caspase-3 activity in the hepatopancreas of S. henanense was lower in animals exposed to higher levels of Cd2+ than in those exposed to more moderate levels. They postulated that Cd-induced apoptosis occurred via a mitochondrial caspase-dependent pathway in low-dosage treatment groups.
The role of antioxidants as a possible defense against heavy metal toxicity has been a focus of toxicological research for the last decade (Orbea et al. 2000; Yan et al. 2007; Wang et al. 2011; Zhang et al. 2012). When heavy metals induce excessive ROS production, the resultant oxidative stress damages macromolecules, including DNA, proteins, and membranes. Many organisms have evolved an antioxidant defense system to protect themselves from these harmful effects of oxidative stress (Geret et al. 2002; Chandran et al. 2005; Ali et al. 2012). Endogenous antioxidants such as GSH, SOD, CAT and GPx, are key components of such systems (Geret et al. 2002; Otitoloju and Olagoke 2011; Loro et al. 2012).
Glutathione (GSH) is an important non-enzymatic antioxidant that is the first line of defense against ROS (Waalkes 2000; Liu et al. 2008; Cao et al. 2012). GSH can inhibit free radical formation through its ability to stabilize components in its oxidative state (Yan et al. 2007; Ivanina et al. 2008; Loro et al. 2012). Under oxidizing conditions, two molecules of GSH are linked by a disulfide bridge to form a molecule of oxidized glutathione (GSSG). Thus, changes in total GSH content, or in the ratio of GSH/GSSG, reflect changes in the normal redox state of cells.
Our results showed that although both GSH and the GSH/GSSG ratio decreased with Cd2+ concentration, there was no obvious change in GSSG levels, except in the 12 mg L−1 Cd2+-treatment group (Fig. 3). A decrease in GSH in the digestive gland appears to be a common response of mollusks to heavy metal exposure that can partly be explained by the GSH molecule’s high affinity for metal (Chandran et al. 2005; Ali et al. 2012).
We observed that a decline in GSH levels, and the GSH/GSSG ratio, was accompanied by an increase in MDA levels (Figs. 3, 4). This negative correlation between GSH and MDA is consistent with the results of previous studies (Chandran et al. 2005; Otitoloju and Olagoke 2011; Ali et al. 2012). We used MDA as an indicator of lipid peroxidation because of its sensitivity to oxidative injury in cells (Geret et al. 2003; Wang et al. 2013; Zhang et al. 2014). Differences in MDA levels between treatment groups clearly indicate that Cd2+ induced lipid peroxidation in hepatopancreatic cells, and that MDA levels increased significantly with Cd2+ concentration (Fig. 4). This suggested that Cd2+ exposure damaged hepatopancreatic cells by elevating ROS levels. A negative relationship between MDA and GSH levels is consistent with this hypothesis.
Antioxidant enzymes are recognized as important biomarkers of heavy metal contamination (Ivanina et al. 2008; Zhang et al. 2009; Loro et al. 2012). SOD, CAT, and GPx play important roles in scavenging free radicals (Orbea et al. 2000; Valavanidis et al. 2006). With GSH as a catalyst, GPx protects tissues from oxidative damage by converting free hydrogen peroxide into water and disulfide glutathione (GSSG) (Geret et al. 2003). Cd2+-induced GPx activity was, however, down-regulated in the hepatopancreas of M. meretrix (Fig. 6). This finding is consistent with that of Chandran et al. (2005) who found that cadmium suppressed GPx activity in the kidney and digestive gland of Achatina fulica. Similarly, Wu et al. (2013) found that GPx activity in the hepatopancreas, intestine and stomach of S. henanense were lower in a high (29.0 mg L−1) Cd2+-treatment group than that in the control group (P < 0.01). Collectively, these results indicate that Cd2+ has a significant inhibitory effect on GPx. GPx activity is dependent on selenium levels. By binding to Cd, GPx’s active site (Se-Cys) can reduce the toxic effects of Cd on an organism. In addition to changes at the active site, this causes GPx to become deactivated (Iszard et al. 1995).
SOD and CAT are also part of the cellular enzymatic antioxidant defense systems that act in combination to counteract oxidative stress (Waalkes 2000; Zhang et al. 2009; Feng et al. 2013; Zheng et al. 2013). SOD catalyzes the breakdown of superoxide radicals to hydrogen peroxide. CAT prevents oxidative stress by degrading hydrogen peroxide (Wang et al. 2011). There is evidence that different concentrations of Cd2+ produce different trends in the activity of these two enzymes in different organisms (Geret et al. 2002; Yan et al. 2007; Wang et al. 2011; Feng et al. 2013). We found the same trend in both CAT and SOD levels in response to Cd2+ concentration. There was a negative relationship between the activity of these enzymes and higher Cd2+ concentration, and the peak activities of CAT and SOD were observed in the 3 mg L−1 Cd2+-treatment group (Fig. 6). This is consistent with the results observed by Yan et al. (2007) in the hepatopancreas of S. yangtsekiense, and those of Wu et al. (2013) who found a negative correlation between Cd2+ concentration and SOD and CAT activity in the hepatopancreas, intestine, and stomach of S. henanens with peak activities in the 7.25 mg L−1 Cd2+-treatment group.
These results can be explained by an initial increase in SOD activity in response to the formation of superoxide radicals induced by Cd2+ exposure. Superoxide is considered as a central component of the signal transduction system that triggers the genes responsible for antioxidant enzyme synthesis, including SOD and CAT (Cong et al. 2012). Low exposure to heavy metals may activate antioxidant protective mechanisms but higher exposure may suppress SOD and CAT activity through the binding of heavy metal ions to the active center of these enzymes, ultimately leading to cytotoxicity (Zhang 2010; Wang et al. 2013).
Exposure to 0.5 and 1 mg L−1 of CdCl2 for 2 days was followed by a decrease in both SOD and CAT activity in the digestive gland and kidney of Achatina fulica (Chandran et al. 2005). Similarly, Company et al. (2006) also found that SOD and CAT activity in the gills of the mussel Bathymodiolus azoricus decreased significantly (P < 0.05) after exposure to Cd2+ (100 μg L−1) for 24–48 h. After 6 days, a significant increase in SOD activity occurred in the gills but there was no significant difference in CAT activity between the control and Cd2+-exposed treatment groups. These results indicate that, irrespective of whether antioxidant enzyme activity is up- or down-regulated, certain dosages and durations of heavy metal exposure can have adverse effects on the activity of some antioxidant enzymes in mollusks (Chandran et al. 2005; Zhang et al. 2009; Cong et al. 2012).
Conclusions
Cadmium induced significant apoptosis and oxidative stress in the hepatopancreas of the clam M. meretrix, even at concentrations far below the LC50. The higher the Cd2+ will be the more cell damage will be caused. Cd2+ inhibited GSH and GPx activity but increased MDA levels and caspase-3 activity. Low Cd2+ concentrations stimulated CAT and SOD activity, but higher Cd2+ concentrations inhibited the activity of these enzymes.
On the basis of these results, we expect that exposure to low levels of cadmium activates the antioxidant system but higher levels inactivate it resulting in cell damage and apoptosis. Caspase-3 activity in M. meretrix was enhanced at lower Cd2+ concentrations, suggesting Caspase-3 as a suitable biomarker for the detection of low levels of Cd2+ in aquatic systems.
Abbreviations
- AO/EB:
-
Acridine orange/ethidium bromide
- CAT:
-
Catalase
- Cd:
-
Cadmium
- GPx:
-
Glutathione peroxidases
- GSH:
-
Glutathione
- GSSG:
-
Glutathione disulfide
- GST:
-
Glutathione transferase
- LC50:
-
Medial lethal concentration
- MDA:
-
Malondialdehyde
- PBS:
-
Phosphate-buffered saline
- ROS:
-
Reactive oxygen species
- SOD:
-
Superoxide dismutase
References
Ali D, Alarifi S, Kumar S, Ahamed M, Siddiqui MA (2012) Oxidative stress and genotoxic effect of zinc oxide nanoparticles in freshwater snail Lymnaea luteola L. Aquat Toxicol 124–125:83–90
Angkeow P, Deshpande SS, Park YC (2002) Redox factor-1: an extra-nuclear role in the regulation of endothelial oxidative stress and apoptosis. Cell Death Differ 9(7):717–725
Bolduc JS, Denizeau F, Jumarie C (2004) Cadmium-induced mitochondrial membrane-potential dissipation does not necessarily require cytosolic oxidative stress: studies using rhodamine-123 fluorescence unquenching. Toxicol Sci 77(2):299–306
Bradford MM (1976) A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein–dye binding. Anal Biochem 72:248–254
Byrne PA, O’Halloran J (2001) The role of bivalve molluscs as tools in estuarine sediment toxicity testing: a review. Hydrobiologia 465:209–217
Cao H, Shi CL, Jia XY (2012) Cadmium exposure to shading frog testis induced and its effect on the protein oxidative damage mechanism of ROS. Acta Ecol Sin 32(13):4199–4206
Chandran R, Sivakumar AA, Mohandass S, Aruchami M (2005) Effect of cadmium and zinc on antioxidant enzyme activity in the gastropod, Achatina fulica. Comp Biochem Physiol Part C 140:422–426
Company R, Serafim A, Cosson R, Camus L, Shillito B, Fiala-Médioni A, Bebianno MJ (2006) The effect of cadmium on antioxidant responses and the susceptibility to oxidative stress in the hydrothermal vent mussel Bathymodiolus azoricus. Mar Biol 148:817–825
Cong M, Wu HF, Liu XL, Zhao JM, Wang X, Lv JS, Hou L (2012) Effects of heavy metals on the expression of a zinc-inducible metallothionein-III gene and antioxidant enzyme activities in Crassostrea gigas. Ecotoxicology 21:1928–1936
Feng MB, Li Y, Qu RJ, Wang LS, Wang ZY (2013) Oxidative stress biomarkers in freshwater fish Carassius auratus exposed to decabromodiphenyl ether and ethane, or their mixture. Ecotoxicology 22:1101–1110
Geret F, Serafim A, Barrira L, Bebianno MJ (2002) Effect of cadmium on antioxidant enzyme activities and lipid peroxidation in the gills of the clam Ruditapes decussates. Biomarkers 7(3):242–256
Geret F, Serafim A, Bebianno MJ (2003) Antioxidant enzyme activities, metallothioneins and lipid peroxidation as biomarkers in Ruditapes decussatus? Ecotoxicology 12:417–426
Hagara H, Malki WA (2014) Betaine supplementation protects against renal injury induced by cadmium intoxication in rats: role of oxidative stress and caspase-3. Environ Toxicol Pharmacol 37:803–811
Hamad A, Amel EG, Saleh F, Magdy EH (2011) Evaluation of heavy metal pollution in the Arabian Gulf using the clam Meretrix meretrix Linnaeus, 1758. Water Air Soil Pollut 214:499–507
Iszard MB, Liu J, Klaassen CD (1995) Effect of several metallothionein inducers on oxidative stress defense mechanisms in rats. Toxicology 104:25–33
Ivanina AV, Cherkasov AS, Sokolova IM (2008) Effects of cadmium on cellular protein and glutathione synthesis and expression of stress proteins in eastern oysters, Crassostrea virginica Gmelin. J Exp Biol 211:577–586
Ji YL, Wang Z, Wang H, Zhang C, Zhang Y, Zhao M, Chen YH, Meng XH, Xu DX (2012) Ascorbic acid protects against cadmium-induced endoplasmic reticulum stress and germ cell apoptosis in testes. Reprod Toxicol 34(3):357–363
Kasibhatla S, Amarante-Mendes GP, Finucane D, Brunner T, Bossy-Wetzel E, Green DR (2006) Acridine orange/ethidium bromide (AO/EB) staining to detect apoptosis. Cold Spring Harbor Protoc. doi:10.1101/pdb.prot4493
Ketata I, Smaoui-Damak W, Guermazi F, Rebai A, Hamza-Chaffai A (2007) In situ endocrine disrupting effects of cadmium on the reproduction of Ruditapes decussatus. Comp Biochem Physiol Part C 146:415–430
Li YJ, Wu H, Wei XX, He YJ, Li BZ, Li YQ, Jing WX, Wang L (2015) Subcellular distribution of Cd and Zn and MT mRNA expression in the hepatopancreas of Sinopotamon henanense after single and co-exposure to Cd and Zn. Comp Biochem Physiol Part C 167:117–130
Liu N, Yan B, Li YQ, Wang Q, Wang L (2008) Effects of Cd2+ on Glutathione system of hepatopancreas and gills in freshwater crab Sinopotamon yangtsekiense. Chin J Environ Sci 29(8):2302–2306
Liu DM, Yang J, Li YJ, Zhang M, Wang L (2013) Cd-induced apoptosis through the mitochondrial pathway in the hepatopancreas of the freshwater crab Sinopotamon henanense. PLoS One 8(7):e68770
Loro VL, Jorge MB, Silva KR, Wood CM (2012) Oxidative stress parameters and antioxidant response to sublethal water borne zinc in a euryhaline teleost Fundulus heteroclitus: Protective effects of salinity. Aquat Toxicol 110–111:187–193
Meng FP, Wang ZF, Cheng FL, Du XP, Fu WC, Wang Q, Yi XY, Li YF, Zhou Y (2013) The assessment of environmental pollution along the coast of Beibu Gulf, northern South China Sea: an integrated biomarker approach in the clam Meretrix meretrix. Mar Environ Res 85:64–75
Nzengue Y, Steiman R, Garrel C, Lefèbvre E, Guiraud P (2008) Oxidative stress and DNA damage induced by cadmium in the human keratinocyte HaCaT cell line: role of glutathione in the resistance to cadmium. Toxicology 243(1–2):193–206
Orbea A, Fahimi HD, Cajaraville MP (2000) Immunolocalization of four antioxidant enzymes in the digestive glands of mollusks and crustaceans and fish liver. Histochem Cell Biol 114(5):393–404
Otitoloju A, Olagoke O (2011) Lipid peroxidation and antioxidant defense enzymes in Clarias gariepinus as useful biomarkers for monitoring exposure to polycyclic aromatic hydrocarbons. Environ Monit Assess 182:205–213
Rainbow PS, Black WH (2007) Cadmium, zinc and the uptake of calcium by two crabs, Carcinus maenas and Eriocheir sinensis. Aquat Toxicol 72(72):42–65
Ribble D, Goldstein NB, Dorris DA, Shellnan YG (2005) A simple technique for quantifying apoptosis in 96-well plates. BMC Biotechnol 5:12–18
Shih CM, Ko WC, Wu JS, Wei YH, Wang LF, Chang EE, Lo TY, Cheng HH, Chen CT (2004) Mediating of caspase-independent apoptosis by cadmium through the mitochondria-ROS pathway in MRC-5 fibroblasts. Cell Biochem 91(2):384–397
Silvestre F, Duchene C, Trausch G, Devos P (2005) Tissue-specific cadmium accumulation and metallothionein-like protein levels during acclimation process in the Chinese crab Eriocheir sinensis. Comp Biochem Physiol 140:39–45
Tietze F (1969) Enzymatic method for quantitative determination of nanogram mounts of total and oxidized glutathione-applications to mammalian blood and other tissues. Anal Biochem 27:502–522
Valavanidis A, Vlahogianni T, Dassenakis M, Scoullos M (2006) Molecular biomarkers of oxidative stress in aquatic organisms in relation to toxic environmental pollutants. Ecotoxicol Environ Saf 64:178–189
Waalkes MP (2000) Cadmium carcinogenesis in review. J Inorg Biochem 79:241–244
Wagner C, Steven R, Koziol C (1998) Apoptosis in marine sponges: a biomarker for environmental stress (cadmium and bacteria). Mar Biol 131(3):411–421
Wang YW, Wei YS, Liu JX (2008) Studies on the accumulation model of heavy metals in aquatic organisms. Acta Scientiae Circumstantiae 28(1):12–20
Wang SH, Zhu FX, Song CM (2009a) River estuary clam breeding status and quality security policy. China Fish 9:83–86
Wang XY, Wang Q, Yang HS (2009b) Acute toxicities of Cd2+ and Hg2+ on Mactra veneriformis Reeve. Mar Sci 33(12):24–29
Wang Q, Wang XM, Wang XY, Yang HS, Liu BH (2010) Analysis of metallotionein expression and antioxidant enzyme activities in Meretrix meretrix larvae under sublethal cadmium exposure. Aquat Toxicol 100(4):321–328
Wang L, Xu T, Lei WW, Liu DM, Li YJ, Xuan RJ, Ma JJ (2011) Cadmium-induced oxidative stress and apoptotic changes in the testis of freshwater crab, Sinopotamon henanense. PLos One 6(11):e27853
Wang XF, Chen WH, Zhang Z, Chen HG, Jia XP (2013) Effects of water-borne mercury and cadmium exposure on lipid peroxidation and antioxidant enzymes in mangrove red snapper Lutjanus argentimaculatus. Res J Appl Sci Eng Technol 6(13):2395–2401
Wätjen W, Cox M, Biagioli M, Beyersmann D (2002) Cadmium-induced apoptosis in C6 glioma cells: mediation by caspase9-activation. Biometals 15:15–25
Wu H, Xuan RJ, Li YJ, Zhang XM, Wang Q, Wang L (2013) Effects of cadmium exposure on digestive enzymes, antioxidant enzymes, and lipid peroxidation in the freshwater crab Sinopotamon henanense. Environ Sci Pollut Res 20(6):4085–4092
Xiang LX, Shao JZ, Meng Z (2001) Apoptosis induction in fish cells under stress of six heavy metal ions. Prog Biochem Biophys 28(6):866–869
Yan B, Wang L, Li YQ, Liu N, Wang X (2007) Effects of cadmium on hepatopancreatic antioxidant enzyme activity in a freshwater crab Sinopotamon yangtsekiense. Acta Zool Sin 53(6):1121–1128
Zhang X (2010) Research advances on superoxide dismutase (SOD). Inn Mong Petrochem 116:14–15
Zhang QS, Hou JJ, Liu XJ, Luo JX, Xiong BX (2009) Effects of copper on antioxidant enzyme activities and metallothionein concentration of Bellamya purificata. Acta Hydrobiol Sin 33(4):717–725
Zhang YK, Song XK, Liu AY, Liu LJ, Jiang XY, Ma JX (2011) Acute toxicity effects of Cd2+ and Cu2+ on Meretrix meretrix Linnaeus. Trans Oceanol Limnol 3:51–56
Zhang HJ, Cai CC, Shi CL, Cao H, Han ZL, Jia XY (2012) Cadmium-induced oxidative stress and apoptosis in the testes of frog Rana limnocharis. Aquat Toxicol 122–123:67–74
Zhang H, Pan LQ, Tao YX (2014) Antioxidant responses in clam Venerupis philippinarum exposed to environmental pollutant hexabromocyclododecane. Environ Sci Pollut Res 21:8206–8215
Zhao S, Feng CH, Quan WM (2012) Role of living environments in the accumulation characteristics of heavy metals in fishes and crabs in the Yangtze River Estuary, China. Mar Pollut Bull 64:1163–1171
Zheng SM, Wang YY, Zhou QX, Chen CH (2013) Responses of oxidative stress biomarkers and DNA damage on a freshwater snail (Bellamya aeruginosa) stressed by ethylbenzene. Arch Environ Contam Toxicol 65:251–259
Acknowledgments
We would like to thank the native speaker Prof. Ron Moorhouse and Sam Denny for revising the English. We would also especially like to thank two anonymous reviewers for correcting some of the English expressions and giving suggestions for improving this manuscript.
Funding
This study was funded the Nature Science Foundation of Zhejiang Province, China (Grant Number LY13C040003) (to XPY), the Science-Technology Innovation Team of Wenzhou City (Grant Number C201210351012) (to XPY), the National Student Innovation Training Project (Grant Number 2014049) (to LPX), MOST 104-2621-M-037-001 to T.H. SHIH, A newcomer project from KMU to HUD (Grant Number KMU-Q104018).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
Liping Xia declares that he has no conflict of interest. Sihan Chen declares that she has no conflict of interest. Hans-Uwe Dahms declares that he has no conflict of interest. Xueping Ying declares that she has no conflict of interest. Xue Peng declares that she has no conflict of interest.
Research involving human and animal rights
All applicable international, national, and/or institutional guidelines for the care and use of animals were followed. This article does not contain any studies with human participants performed by any of the authors.
Rights and permissions
About this article

Cite this article
Xia, L., Chen, S., Dahms, HU. et al. Cadmium induced oxidative damage and apoptosis in the hepatopancreas of Meretrix meretrix . Ecotoxicology 25, 959–969 (2016). https://doi.org/10.1007/s10646-016-1653-7
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10646-016-1653-7