
Abstract
1H NMR-based metabolomics was used to examine the metabolic profile of D2O-buffer extracted tissues of Eisenia fetida earthworms exposed for 2 days to an artificial soil spiked with sub-lethal concentrations of polychlorinated biphenyls (PCBs) (0, 0.5, 1, 5, 10, or 25 mg/kg Aroclor 1254). Univariate statistical analysis of the identified metabolites revealed a significant increase in ATP concentration in earthworms exposed to the highest soil PCB concentration, but detected no significant changes in other metabolites. However, a multivariate approach which considers alterations in multiple metabolites simultaneously, identified a significant linear relationship between earthworm metabolic profiles and PCB concentration (cross-validated PLS-regression with 7 components, R2X = 0.99, R2Y = 0.77, Q2Y = 0.45, P < 0.001). Significant changes in pair-wise metabolic correlations were also detected as PCB concentration increased. For example, lysine and ATP concentrations showed no apparent correlation in control earthworms (r = 0.22, P = 0.54), but were positively correlated in earthworms from the 25 mg/kg treatment (r = 0.87, P = 0.001). Overall, the observed metabolic responses suggest that PCBs disrupted both carbohydrate (energy) metabolism and membrane (osmolytic) function in E. fetida. The ability of 1H NMR-based metabolomics to detect these responses suggests that this method offers significant potential for direct assessment of sub-lethal PCB toxicity in soil.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Although no longer in widespread production or use, polychlorinated biphenyls (PCBs) are a persistent soil contaminant due to their hydrophobicity and resistance to biodegradation (Weber et al. 2008) and remain a source of risk to various ecological receptors (Cooper and Roch 1992; Johnson et al. 2009; Landrum et al. 2004; Meier et al. 1997; Suzuki et al. 1995; Ville et al. 1995). As with other soil contaminants, remediation guidelines for PCBs are often set in relation to a total soil PCB concentration (CCME 1999; O’Halloran 2006), which does not account for differences in PCB mixture composition or variations in PCB bioavailability related to soil properties and age of contamination (Alexander 2000; Semple et al. 2003). Direct assessment of soil contaminant toxicity using ecologically relevant target organisms such as earthworms could help prioritize remediation efforts and decrease site remediation costs (Sousa et al. 2008). Several standardized tests for assessing the toxicity of contaminated soil to earthworms exist (e.g. Environment Canada 2004; OECD 1984, 2004), but these methods remain relatively costly and time-consuming. For instance, the acute earthworm toxicity test requires 14 days (Environment Canada 2004; OECD 1984), and since it only measures lethality it may be insufficient for predicting long-term population fitness following chronic exposure to contaminated soil. For example, Meier et al. (1997) reported that a weathered PCB-contaminated soil (144 mg/kg Aroclor 1260) which caused no lethality to earthworms after 14 days completely suppressed earthworm reproduction after only 21 days. Clearly, inhibition of earthworm reproduction provides a more ecologically relevant sub-lethal endpoint than lethality, but it requires an even longer exposure period for accurate assessment. Standardized earthworm reproduction tests require >50 days to complete (Environment Canada 2004; OECD 2004). Sub-lethal earthworm responses to PCB exposure have also been detected using short duration (5-day) contact tests which demonstrated that earthworms can experience immunosuppression and decreased wound healing after sub-lethal PCB exposures (Cooper and Roch 1992; Ville et al. 1995). However these tests were based on dermal exposure to contaminated filter paper and have not been adapted for direct application to field-contaminated soil. Therefore, there is interest in developing more rapid methods to predict sub-lethal soil contaminant toxicity to earthworms.
Nuclear magnetic resonance (NMR)-based metabolomics has been proposed as an alternative method for measuring sub-lethal toxicity of contaminated soil to earthworms (Brown et al. 2010; Bundy et al. 2004, 2008; Gibb et al. 1997; Guo et al. 2009; Jones et al. 2008; McKelvie et al. 2010; Rochfort et al. 2009). This method provides insight into the biochemical response of a study organism to an environmental stimulus by comparing the concentrations of endogenous metabolites in a collected tissue or biofluid under varied environmental conditions (Viant 2008). Interpretation of any changes in the metabolic profile (‘metabolome’) can reveal information about the organism’s response to contaminant exposure at the metabolic level as well as the contaminant’s toxic mode of action (Hines et al. 2010; Viant 2008; Vulimiri et al. 2010). To date, many metabolomic studies of earthworm responses to contaminated soil have used soil exposure periods which were comparable to or longer than the 54 day exposure required for the earthworm reproduction test (i.e. 21–110 days in Bundy et al. 2004, 2008; Gibb et al. 1997; Guo et al. 2009; Jones et al. 2008) or collected earthworms directly from field-contaminated soil (Bundy et al. 2004, 2007). However, preliminary results by Brown et al. (2010) suggested that NMR-based metabolomics can detect earthworm metabolic responses to contaminated soil after as little as 2 days of soil exposure. Therefore, NMR-based metabolomics may provide a method to assess sub-lethal toxicity of contaminated soil that is much faster than traditional ecotoxicological approaches.
1H NMR-based metabolomics has not yet been applied to assess earthworm responses to PCB contamination. In this study, Eisenia fetida earthworms were exposed for 2 days to PCB-spiked artificial soil (0–25 mg/kg Aroclor 1254) and their metabolic profiles measured using 1H NMR spectroscopy. This represents a range of soil PCB concentrations considered acceptable for protection of human and environmental health depending on land use according to both the U.S. Environmental Protection Agency (EPA 1990) and the Canadian Council of Ministers of the Environment (CCME 1999). Both univariate and multivariate statistical methods were applied to distinguish metabolic responses of the earthworms to PCBs. By measuring and quantifying the metabolic response of earthworms to sub-lethal PCB exposure in freshly spiked artificial soil, this study provides a foundation for future metabolomic studies to directly assess PCB sub-lethal toxicity in aged, field-contaminated soils.
Materials and methods
Soil spiking and total soil PCB concentration
An artificial soil consisting of 10% sphagnum peat (Ward’s Natural Science), 20% kaolin clay (Ward’s Natural Science), and 70% sand (Ward’s Natural Science) was prepared as described in the OECD Earthworm Acute Toxicity test protocol (OECD 1984). Initially, 125 g (dry weight) of this soil was added to each of six 1 l clear glass jars. For the five PCB-exposed treatments, soils were spiked with 10 ml of 25, 50, 250, 500, or 1250 mg/l Aroclor 1254 (Sigma-Aldrich, ON, Canada) in dichloromethane (DCM, HPLC grade, Fisher Scientific). The unexposed control treatment was treated with 10 ml of DCM only. All treatments were left in a fume hood for 16 h to allow the DCM to evaporate (Brinch et al. 2002) and then an additional 375 g (dry weight) of soil was mixed thoroughly into each jar, resulting in total soil PCB concentrations of 0.5, 1, 5, 10, and 25 mg/kg (dry weight) for the PCB-exposed treatments. All soils were then wetted with deionized water to a moisture content of 35% of soil dry weight (OECD 1984) and allowed to absorb the water for 24 h before introducing earthworms to the jars. Spiked soil PCB concentrations were confirmed by soxhlet extraction and quantification via gas chromatography/mass spectrometry following the 48 h earthworm exposures to the soils described in “Earthworm exposure and preparation for 1H NMR” Section, (Section S1, Table S1 in Supplementary Material).
Earthworm exposure and preparation for 1H NMR
Earthworms were selected from a healthy population of E. fetida maintained within our laboratory since 2006 as described in Brown et al. (2008). The original progenitor earthworms were purchased from The Worm Factory (ON, Canada). Ten mature earthworms with a visible clitellum were added to each of the five PCB spiked soils and the control soil. Initial earthworm mass ranged from 240 to 490 mg (mean 360 ± 8.5 mg (standard error) wet weight), with no significant differences in the initial mass between the six treatments (ANOVA, F5,54 = 1.88, P = 0.11). Earthworms were kept in closed jars for 48 h at 21°C in natural light (Eijsackers et al. 2001), then removed and depurated for 96 h on damp filter paper (Brown et al. 2008). Earthworms were then flash-frozen in liquid nitrogen, lyophilized, reweighed and stored frozen until extraction.
Lyophilized earthworms were homogenized in a 1.5 ml centrifuge tube using a 5 mm wide stainless steel spatula. The homogenized tissue was extracted using 1.20 ml of a 0.2 M monobasic sodium phosphate buffer solution (NaH2PO4·2H2O; 99.3%; Fisher Chemicals) containing 0.1% (w/v) sodium azide (99.5% purity; Sigma-Aldrich) as a biocide and 10 mg/l of 2,2-dimethyl-2-silapentane-5-sulfonate sodium salt (DSS; 97%, Sigma-Aldrich) as an internal standard (Brown et al. 2008). Buffer solution was made with D2O (99.9% purity, Cambridge Isotope Laboratories) and adjusted to a pD of 7.4 using NaOD (30% w/w in 99.5% D2O, Cambridge Isotope Laboratories Inc). Samples were vortexed for 30 s using a VX 100 vortexer (Labnet, NJ, USA) and sonicated for 15 min using a FS60 mechanical ultrasonic cleaner (Fisher Scientific) to aid with the extraction. Subsequently, samples were centrifuged at 14,000 rpm (~15,000×g) for 20 min using an International Equipment Company 21000 centrifuge (Fisher Scientific) and the supernatant was transferred into a new 1.5 ml centrifuge tube. The centrifugation procedure was repeated twice and the final sample extracts were transferred into 5 mm High Throughputplus NMR tubes (Norell Inc.; NJ, USA).
1H NMR spectroscopy
1H NMR spectra of the earthworm extracts were acquired with a Bruker Avance 500 MHz spectrometer using a 1H–19F–15N–13C 5 mm Quadruple Resonance Inverse (QXI) probe fitted with an actively shielded Z gradient. 1H NMR experiments were performed using Presaturation Using Relaxation Gradients and Echoes (PURGE) water suppression and 128 scans, a recycle delay of 3 s, and 16 K time domain points (Simpson and Brown 2005). Spectra were apodized through multiplication with an exponential decay corresponding to 0.3 Hz line broadening in the transformed spectrum and a zero filling factor of 2, then manually phased and calibrated to the DSS methyl singlet, set to a chemical shift (δ) of 0.00 ppm. Metabolite peaks were identified in each spectrum by comparison with previously published assignments (Brown et al. 2008; Cui et al. 2008; Lenz et al. 2005).
Data analysis
1H NMR spectra between δ of 0.5 and 10 ppm were divided into 0.01 ppm wide buckets and the area under each segment was integrated with AMIX 3.7.10 (Bruker BioSpin) with the integration mode of sum of intensities. This resulted in peak splitting for some of the identified metabolites, so bucket widths were adjusted for these compounds (Bundy et al. 2007; Guo et al. 2009; Table S2 in Supplementary Material). With the remainder of each spectrum divided into 0.01 ppm wide buckets and the area between δ = 4.70–4.85 ppm excluded to eliminate small residual H2O/HOD signals, this resulted in a total of 906 buckets for each earthworm spectrum. The data were normalized using Probabilistic Quotient Normalization (PQN) to correct for differences in the total NMR signal measured for each sample which can vary with factors such as the mass of tissue extracted (Brown et al. 2009, 2010; Craig et al. 2006; McKelvie et al. 2009, 2010). PQN has been proposed to be a robust normalization method for metabolomic analysis of complex biological mixtures (Dieterle et al. 2006) and has been applied in several recent environmental metabolomics studies (Alam et al. 2010; Bohus et al. 2009; Bundy et al. 2008; Hughes et al. 2009; Taylor et al. 2009). In a test dataset of 24 healthy control earthworms, PQN decreased overall inter-individual metabolic variability in comparison to integral normalization [IN, ‘scaled to total intensity’, (Craig et al. 2006)] and produced a pattern of pair-wise metabolic correlations which more closely matched that observed for quantified metabolite concentrations than did the pattern of pair-wise metabolic correlations calculated using IN data (Whitfield Åslund et al. in press). Since the multivariate statistics commonly applied to interpret metabolomics data [principal components analysis (PCA) and partial least squares (PLS)] are derived from data correlation and/or covariance, PQN normalized data was selected for all further analysis in this study.
Analysis of variance (ANOVA) was used to compare earthworm mass between treatments. For identified metabolites, Dunnett’s multiple comparison tests were applied to compare the PQN normalized bucket intensities between the control earthworms and each of the five PCB-exposed treatments.
Both principal components analysis (PCA) and partial least squares (PLS) regression were applied to detect trends in the earthworm metabolic profile related to PCB exposure. Background and information regarding these methods and their interpretation can be found in Eriksson et al. (2006). PCA was performed using the covariance matrix since the variables have been normalized to the same scale (Jackson 1991). PLS regression, via the NIPALS PLS algorithm (Vendeginste et al. 1998), used the bucketed and normalized NMR spectra as the ‘X’ matrix of multiple predictors and soil PCB concentrations as the ‘Y’ (response) matrix. PLS models were cross validated using leave-one-out cross validation (Hawkins et al. 2003; Varmuza and Filzmoser 2009) and the number of components selected for each final PLS model was determined using the 1CV strategy described by Westerhuis et al. (2008). For each PLS model, the explained variation of X and Y (R2X and R2Y) were reported to indicate how well the model fit the training data (Eriksson et al. 2006) and the internally cross-validated R2Y value [reported as Q2Y (Cramer 1993)] was reported as a preliminary measure of the predictive ability of the model (Hawkins et al. 2003; Varmuza and Filzmoser 2009). An alternative PLS model was also created using only 80% of the data and then used to predict the remaining data in order to simulate how well as PLS model built from this data could predict an external test set. In addition, the significance of each PLS model was estimated through response permutation testing (Alam et al. 2010; Eriksson et al. 2006; Section S2 in Supplementary Material).
Pair-wise Pearson correlation coefficients (r) were calculated between the PQN normalized intensities of all identified metabolites in earthworm extracts grouped by treatment. To control Type I errors due to calculations of multiple pair-wise correlation coefficients, correlation significance was assessed by comparison to a Bonferroni adjusted α value [αadjusted = α/c where c represents the number of simultaneous comparisons (Broadhurst and Kell 2006; Camacho et al. 2005)]. For each metabolite pair, the correlation coefficient measured in control earthworm extracts (r0) was contrasted to that calculated for extracts of earthworms exposed to the highest soil PCB concentration (r25) using the Fisher Z-transformation (1) and comparison to the z-distribution (Spiegel 1991; Morgenthal et al. 2006). Metabolite pairs for which r0 and r25 differed significantly were selected for further examination.
Unless otherwise specified, statistical significance was assessed at α = 0.05. Means were reported as the mean value ± standard error. ANOVAs, Dunnett’s multiple comparison tests and correlation analyses were performed in SPSS 17.0. Multivariate statistics (PCA and PLS) and permutation tests were performed in R (R Development Core Team 2009) using the Chemometrics package (Filzmoser and Varmuza 2010).
Results and discussion
Univariate and multivariate analysis of earthworm metabolomic responses to PCB exposure
Following the approach of Bundy et al. (2008), the identified metabolites were categorized into one of five functional groups (sugars, osmoregulators, energy transfer molecules, amino acids, or organic acids; Fig. 1). Although some metabolites have functions not represented by these groups and/or may belong in multiple categories, this classification fit the patterns of earthworm responses to PCB exposure (Fig. 1). A visual inspection of the normalized metabolite intensities of the identified metabolites suggests that some metabolites increased or decreased following PCB exposure (e.g., possible increases in maltose, glucose, and betaine; Fig. 1), but these differences were statistically insignificant for most metabolites due to high inter-individual metabolic variability (Dunnett’s multiple comparison test, P > 0.05; Fig. 1). The only statistically significant metabolic response was an increase in the PQN normalized intensity of ATP observed in earthworm extracts from the 25 mg/kg PCB soil treatment (Dunnett’s multiple comparison test, P = 8.5 × 10−4, Fig. 1c). Interestingly, Berdanier et al. (1975) reported that PCB exposure increased ATP and decreased ADP, AMP, and Pi in the livers of weanling rats. PCBs were hypothesized to have inhibited active transport pumps fueled by ATP (transmembrane ATPases), resulting in an impairment of membrane function (Berdanier et al. 1975). Similarly, inhibition of ATPases such as Na+/K+ ATPase and Mg2+ ATPase has been reported following PCB exposure in rainbow trout (Davis et al. 1972), rats (La Rocca and Carlson 1979), and Atlantic salmon (Lerner et al. 2007). Furthermore, Gastaldi et al. (2007) suggested that decreased ATPase activity may be part of a general stress response in earthworms, as the earthworm E. andrei exhibited significant decreases in the activity of Ca2+ ATPase in the intestinal epithelium following 1 day of exposure to either copper or benzo[a]pyrene. A similar mechanism may have increased ATP concentrations in PCB-exposed E. fetida in the current study. Since ATPases are critical to the transport of essential nutrients into the cell and for cell osmotic control (Dhalla and Zhao 1988; Lingrel and Kuntzweiler 1994), this suggests that earthworms exposed to the 25 mg/kg Aroclor 1254 spiked soil in this study may have been under considerable stress.

Metabolite responses to PCB exposure, expressed as a percent change from the mean metabolite intensity observed in control (unexposed) earthworms for a Sugars, b Osmoregulators, c Energy transfer molecule, d Amino acids and e Organic acids. Error bars represent standard error of the mean. *Signifies a metabolite intensity in a PCB exposed group found to be significantly different from that of the unexposed control (P < 0.05) using Dunnett’s multiple comparison test
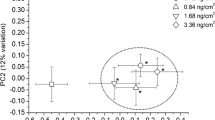
Since multivariate statistics consider many variables simultaneously, they can detect meaningful trends in metabolomic data that may not be identified by traditional univariate analyses (Varmuza and Filzmoser 2009). Initially, PCA was applied as an ‘exploratory data analysis’ tool (Broadhurst and Kell 2006; Verouden et al. 2009) to help identify general similarities and differences in the data. This PCA produced four principal components (PCs) which explained 97.4% of the total data variance (PCs 1, 2, 3, and 4 explained 73, 19, 4.5, and 0.9% of the total variance, respectively). A plot of the sample scores for PC1 versus PC2, which together explained 92% of the total data variance, provided a general separation between control and PCB-exposed earthworms (along PC1) but little dose dependent separation between earthworms exposed to ‘high’ and ‘low’ soil PCB concentrations (Figure S1 in Supplementary Material). For instance, this scores plot demonstrates overlap between the mean PC scores (±standard error) of earthworms exposed to the highest soil PCB concentrations (25 mg/kg) and those exposed to the lowest soil PCB concentration (0.5 mg/kg), and provides a better separation between control earthworms and those exposed to 1.0 mg/kg PCBs in soil than it does between control earthworms and those exposed to 25 mg/kg PCBs in soil (Figure S1 in Supplementary Material). However, PCs are constructed to explain maximum amounts of total data variance without discrimination regarding the source of the variation. Therefore the PCs which explain the largest amounts of data variance may not always be related to the experimental treatment since PCs also incorporate other sources of metabolic variation such as sampling error and natural biological variation (Verouden et al. 2009). As a result, it can be useful to consider higher order PCs in the interpretation of metabolomic datasets (Broadhurst and Kell 2006; Bundy et al. 2004; Johnson et al. 2007; Rochfort et al. 2009; Rousseau et al. 2008; Schock et al. 2010; Scholz and Selbig 2007; Warne et al. 2000). In this study, a better concentration-dependent separation of the PC scores was obtained by plotting PC1 versus PC3 (Fig. 2), which together explained 77.5% of the total variance. This suggests that the metabolic profile of PCB-exposed earthworms could be distinguished from that of control earthworms, as the mean PC scores of earthworms exposed to PCB concentrations greater than 0.5 mg/kg (±standard error) lie outside of the mean PC scores (±standard error) of the control (unexposed) earthworms (Fig. 2). PC1 primarily separated the control earthworms from the PCB-exposed earthworms and PC3 provided some concentration-dependent separation between soils spiked with ‘low’ and ‘high’ PCB concentrations. Since PC1 explained a greater amount of the total data variance than PC3, changes in the earthworm metabolic profile were more pronounced between ‘control’ and ‘exposed’ earthworms than between earthworms exposed to different soil PCB concentrations.

a PCA scores plot (PC1 vs. PC3) for a PCA performed using the PQN normalized bucket table for control and PCB exposed E. fetida aqueous tissue extracts. Each point represents the mean PC score for the treatment class and error bars represent the standard error of the mean. b The corresponding PC1 and c PC3 loadings versus chemical shift. Spectral regions contributing to the observed variance are labeled with selected metabolites. Mal maltose, Glu glucose, Bet betaine, Glut glutamate, Ala alanine, Val valine, Leu leucine
Although PCA detected some concentration-dependent variation in the earthworm metabolic profile, considerable overlap in the scores plot remains between different PCB-exposed treatment groups (Fig. 2a). This may be due to biological metabolic variation, which is not related to the PCB treatment, but is nevertheless included in the unsupervised PCA model (Parsons et al. 2009). Since PLS analysis focuses on variation between treatment groups, it may provide a better discrimination between each of the PCB treated groups (Eriksson et al. 2006). Therefore, a PLS model was constructed using the spiked soil PCB concentrations as the Y-table and the bucket intensities of all earthworms in the study as the X-table. Cross-validated prediction error was minimized for a model with seven components, so this PLS model was saved for further interpretation (Westerhuis et al. 2008). This model fit the training set data well (R2X = 0.99, R2Y = 0.77), but a leave-one-out cross-validation suggested that only 45% of the variation in soil PCB concentration could be predicted by the earthworm metabolomic responses (Q2Y = 0.45) (Eriksson et al. 2006). Nevertheless, a permutation test confirmed that the predictive capacity of this model was significantly better than what would be expected if random treatment labels had been applied to each earthworm (P = 9.0 × 10−5, Figure S2 in Supplementary Material). The best fit linear relationship between the observed and predicted soil PCB concentrations for the cross-validated predictions is given by [PCB]predicted = 0.56([PCB]observed) + 2.9, which has an R²Y of 0.47 and provides a predicted PCB concentration of 2.9 mg/kg for the control soil (Fig. 3). However, the data appear to more closely match a log–log relationship, with a rapid increase between the control group and the lower PCB concentrations and a near plateau at the higher PCB concentrations (Fig. 3). When the data was transformed to eliminate the ‘zero’ PCB concentration of the control using the transformation [PCB] + 1 (Bundy et al. 2007; McKelvie et al. 2010), the cross-validated predictions fit a logarithmic regression relationship of [PCB]predicted = 4.58(ln([PCB]observed +1)) + 0.22, which had a similar R²Y of 0.54 and provided a lower predicted PCB concentration of 0.22 mg/kg for the control soil (Fig. 3). When the PLS model was reconstructed using only 80% of the total data with 20% left out for application as an external test set, similar patterns were noted for the predicted values of the external test set (Section S3, Figures S3, S4 in Supplementary material). This pattern may suggest that the metabolic pathways mediating the earthworm response to PCBs were saturated at the higher soil PCB concentrations so that no further toxicity was expressed (Steenland and Deddens 2004). Ville et al. (1995) noted a similar log–log pattern of response for lysozyme activity in two closely related Eisenia species (E. hortensis and E. andrei) following a five-day contact exposure to PCBs (exposure concentrations of 0, 1, 5, 10, and 30 μg/cm2) which suggests that this pattern of toxic response may be characteristic of the reponse to PCB exposure for the Eisenia species. Overall, the PLS models suggest that the earthworm metabolic profiles can be weakly but significantly correlated to soil PCB concentration, and that the biochemical processes mediating the PCB toxic response modeled by PLS may have become saturated at higher concentrations.

Average predictions of PCB concentrations (\( \hat{y}_{i} \)) given spectra i by the PLS model derived during the leave one out cross-validation procedure with spectra i omitted for PLS models (7 components) constructed with soil PCB concentration as the Y variable and the quotient normalized bucket intensities as the X table. The solid line indicates a linear regression between the actual and predicted values ([PCB]predicted = 0.56([PCB]observed) + 2.9, R²Y = 0.47) and the dashed line indicates the a logarithmic relationship between the actual and predicted values ([PCB]predicted = 4.58(ln([PCB]observed + 1)) + 0.22, R²Y = 0.54). Error bars represent the standard error of the mean
For the PLS model constructed using the complete dataset, the cumulative variable importance to projection (VIP) values were calculated to ascertain which variables contributed most to the model (Eriksson et al. 2006). According to this method, the top ten most important unique and identifiable variables in this PLS model, in decreasing order of importance, were leucine, glucose, betaine, glycine, glutamate, alanine, maltose, valine, lysine, and lactate (Figure S5 in Supplementary Material). The univariate patterns of these metabolite intensities were therefore re-examined, even though no statistically significant difference from the control had been detected when metabolites were considered individually (Fig. 1). The mean intensities of the sugars (maltose and glucose) in the top 10 VIP list were generally elevated in comparison to the control for all PCB-exposed earthworms (Fig. 1). This agrees with several earlier earthworm metabolomic studies which reported increased sugar levels following exposure to pesticides (Rochfort et al. 2009), naphthalene and phenanthrene (Brown et al. 2009) and 3-trifluoromethylalanine (Warne et al. 2000). However, this response is not universal, as other earthworm metabolomic studies have reported decreased sugar levels following earthworm exposure to contaminants such as copper (Bundy et al. 2008) and phenanthrene (Brown et al. 2010). The observed increase in sugars in the current study may be related to a down-regulation in glycolysis, since glycolysis is inhibited by ATP (Horton et al. 2006), which was observed to have increased in PCB-exposed worms (Fig. 1). Furthermore, Leroy et al. (2010) reported that PCBs inhibited two key enzymes in the glycolytic pathway (enolase and glyceraldehyde-3-phosphate dehydrogenase) in a freshwater amphipod, so it is known that PCBs can disrupt carbohydrate metabolism.
For the osmoregulators identified in the VIP list, mean metabolite intensities were generally increased for betaine and decreased for glycine in PCB-exposed earthworms (Fig. 1). As it has been suggested that PCB exposure may lead to complication in cellular osmotic control due to inhibition of transmembrane ATPases (Berdanier et al. 1975; Davis et al. 1972; La Rocca and Carlson 1979; Lerner et al. 2007), it is possible the levels of these organic osmolytes were manipulated in an attempt to stabilize cellular osmotic potential and prevent cell lysis. Other earthworm metabolomics studies have reported increased levels of betaine following exposure to contaminants as diverse as copper (Bundy et al. 2008), cadmium chloride, atrazine, and fluoranthene (Guo et al. 2009). In addition, Guo et al. (2009) demonstrated that increased betaine was strongly correlated with decreased reproduction rate in earthworms following exposure to both inorganic and organic contaminants.
The mean intensities of the amino acids in the top ten VIP list (leucine, glutamate, alanine, valine, and lysine) exhibited a variable response to earthworm PCB exposure (Fig. 1). For earthworms exposed to the low soil PCB concentrations (≤10 mg/kg), mean amino acid intensities were decreased in comparison to those of the untreated control earthworms. However, the mean intensities of these amino acids were increased in comparison to those of the control earthworms for the highest soil PCB concentration (25 mg/kg). Bundy et al. (2008) observed a similar pattern (decreased below control at low concentrations and increased above control at high concentrations) for the amino acids phenylalanine, tyrosine, valine, and leucine in earthworms exposed to a range of copper concentrations for 70 days. Likewise, other earthworm metabolomic studies have reported both increases (Brown et al. 2009, 2010; Jones et al. 2008; McKelvie et al. 2009, 2010; Rochfort et al. 2009) and decreases (Brown et al. 2009; McKelvie et al. 2010) in various amino acids which differ between the type of contaminant, length of exposure, and/or concentration. As PCBs have been demonstrated to interfere with glycolysis and therefore the production of pyruvate as a substrate for the citric acid cycle (Leroy et al. 2010), it is possible that amino acids were converted to pyruvate, acetyl CoA, acetoacetate or other citric acid cycle intermediates to supplement energy production (Horton et al. 2006). This could explain the initial decrease of amino acids in the current study as well as that observed for the organic acid lactate (also one of the top ten VIP metabolites), which can also be converted to pyruvate (Horton et al. 2006). A similar mechanism was proposed by Van Scoy et al. (2010) to have decreased lactate levels in salmon smolts exposed to both crude and dispersed oil. In contrast, increased amino acid levels in earthworms have previously been attributed to breakdown of muscle or other tissue under toxic stress (Brown et al. 2009, 2010; Jones et al. 2008; McKelvie et al. 2009, 2010; Rochfort et al. 2009). Therefore, the apparent rebound in the mean intensity of amino acids in earthworms from the highest soil PCB concentration (Fig. 1) may indicate that this concentration was sufficient to induce a rate of earthworm tissue degradation that exceeded the rate at which the resulting amino acids could be oxidized for energy production.
Alterations in metabolite correlations following PCB exposure
Traditional approaches to environmental metabolomics measure changes in the concentrations or relative proportion of key metabolites in organisms after exposure to an environmental stressor (Bundy 2005; Robertson 2005; Simpson and McKelvie 2009). However, some research has suggested that metabolic responses may also be reflected through changes in the correlations between specific metabolites (Camacho et al. 2005; Steuer 2006). In the current study, the strength of the correlation (r) of several metabolites to ATP, betaine, and valine was observed to differ significantly in earthworms at the highest soil PCB concentration (25 mg/kg) in comparison to the control earthworms (comparison of correlation coefficients performed by Fisher Z-transformation [1], P < 0.023, P < 0.012, and P < 0.04, respectively).
Specifically, the relationship of a number of the amino acids (glutamate, lactate, leucine, lysine, phenylalanine, tyrosine, and valine) with ATP changed from a weak negative correlation in the control earthworms (mean r = −0.08 ± 0.05, mean P = 0.74 ± 0.1, Fig. 4a) to a strong positive correlation in the earthworms exposed to 25 mg/kg PCBs (mean r = 0.86 ± 0.01, mean P = 0.001 ± 2.6 × 10−4, Fig. 4a). This positive correlation first emerged in earthworms exposed to soil PCB concentrations of 1.0 mg/kg, and became increasingly pronounced as soil PCB concentration was increased (Fig. 4a). In addition, in control earthworms, several amino acids (alanine, glutamate, lactate, leucine, lysine, phenylalanine, and tyrosine) showed a reasonably strong negative correlation with betaine (mean r = −0.685 ± 0.009, mean P = 0.030 ± 3.09 × 10−3, Fig. 4b), but this relationship was reversed to a weak positive relationship for earthworms exposed to soil PCB concentrations greater than or equal to 1.0 mg/kg (e.g., mean r = 0.44 ± 0.012, mean P = 0.211 ± 0.014 for earthworms in the 25 mg/kg treatment group, Fig. 4b). These changes in metabolite correlation between numerous amino acids and both ATP and betaine suggested that PCB exposure significantly altered aspects of both energy metabolism and membrane (osmolytic) function in E. fetida. This is congruent with the findings of both the univariate and multivariate (PCA and PLS) statistical analyses of earthworm metabolic profiles in the current study, and also with earlier studies that showed PCB exposure could disrupt these metabolic functions at an enzymatic level in diverse species such as rats, fish, and a freshwater amphipod (Berdanier et al. 1975; Davis et al. 1972; La Rocca and Carlson 1979; Lerner et al. 2007; Leroy et al. 2010). Moreover, the increasing correlation between ATP and lactate as well as a number of amino acids supports the hypothesis that amino acids and lactate are used as an alternative energy source by E. fetida earthworms following PCB disruption of glycolysis as suggested by the results of the univariate and multivariate analyses in this study. In contrast, although a significant increase in the correlation coefficient was detected between lactate and valine and between lysine and valine from the control earthworms (mean r = 0.78 ± 0.05, mean P = 0.01 ± 0.0067, Fig. 4c) to those exposed to 25 mg/kg of PCBs (mean r = 0.99 ± 7.06 × 10−4, mean P = 7.64 ± 0.18 × 10−8, Fig. 4c), a visual inspection of these relationships at each PCB exposure level (Fig. 4c) did not suggest any change in the underlying metabolic network as a strong positive correlation between these metabolites is conserved in tissue extracts from all soil PCB concentrations.

Examples of metabolite correlations which were altered significantly by PCB exposure. a Lysine:ATP. This pattern of correlation change is representative of similar changes observed between ATP and the following metabolites: glutamate, lactate, leucine, phenylalanine, tyrosine, and valine (not shown) b Lysine:betaine. This pattern of correlation change is representative of similar changes observed between betaine and the following metabolites: alanine, glutamate, lactate, leucine, phenylalanine, and tyrosine. (data not shown) c Lysine:valine. This pattern of correlation change is representative of similar changes observed between valine and lactate (data not shown)
Conclusions
This study provides several lines of evidence suggesting that changes in the earthworm metabolic profile can be directly and quantifiably correlated to soil PCB concentration in a freshly spiked, artificial soil after 2 days of exposure. These changes appear to match well with known toxic modes of action for PCBs that have been previously identified using longer exposure periods and/or other organisms. Although further study is required to explicitly link metabolic profile changes to traditional ecotoxicological endpoints, the results of this study suggest that 1H NMR metabolomics of D2O-buffer extracted earthworm metabolites may be useful to directly monitor the bioavailability and toxicity of PCBs in soil in a much shorter period of time than traditional methods. In addition, this study demonstrated that earthworm metabolomics is highly sensitive to PCB exposure, as changes in the metabolic profile were noted even at the lowest soil PCB concentrations in the study (0.5 mg/kg). However, since the current study used freshly spiked artificial soil, this may have overestimated earthworm sensitivity to PCBs in a realistic field setting as contaminant availability to soil organisms (and therefore toxicity) is expected to decrease over time (Alexander 2000; Semple et al. 2003). For example, Meier et al. (1997) reported no lethality (measured after 14 days) or reproduction impairment (measured after 21 days) in E. fetida exposed to an aged and weathered soil collected from a former electrical transformer storage yard that was contaminated with approximately 70 mg/kg of PCBs (Aroclor 1260). Therefore, future work is required to investigate the application of earthworm metabolomics to the assessment of PCB toxicity in weathered, field-contaminated soils such that this technique can be adapted for use as a risk assessment tool.
References
Alam TM, Alam MK, Neerathilingam M, Volk DE, Sarkar S, Shakeel Ansari GA, Luxon BA (2010) 1H NMR metabonomic study of rat response to tri-phenyl phosphate and tri-butyl phosphate exposure. Metabolomics 6:386–394
Alexander M (2000) Aging, bioavailability, and overestimation of risk from environmental pollutants. Environ Sci Technol 34:4259–4265
Berdanier CD, Tobin RB, Nielsen RC (1975) Effect of polychlorinated biphenyls and thiamin deficiency on liver metabolism in growing rats. J Toxicol Environ Health 1:91–105
Bohus E, Rácz A, Noszál B, Coen M, Beckonert O, Keun HC, Ebbels TMD, Cantor GH, Wijsman JA, Holmes E, Lindon JC, Nicholson JK (2009) Metabonomic investigations into the global biochemical sequelae of exposure to the pancreatic toxin 1-cyano-2-hydroxy-3-butene in the rat. Magn Reson Chem 47:S26–S35
Brinch UC, Ekelund F, Jacobsen CS (2002) Method for spiking soil samples with organic compounds. Appl Environ Microbiol 68:1808–1816
Broadhurst DI, Kell DB (2006) Statistical strategies for avoiding false discoveries in metabolomics and related experiments. Metabolomics 2:171–196
Brown SAE, Simpson AJ, Simpson MJ (2008) Evaluation of sample preparation methods for nuclear magnetic resonance metabolic profiling studies with Eisenia fetida. Environ Toxicol Chem 27:828–836
Brown SAE, Simpson AJ, Simpson MJ (2009) 1H NMR metabolomics of earthworm responses to sub-lethal PAH exposure. Environ Chem 6:432–440
Brown SAE, McKelvie JR, Simpson AJ, Simpson MJ (2010) 1H NMR metabolomics of earthworm exposure to sub-lethal concentrations of phenanthrene in soil. Environ Pollut 158:2117–2123
Bundy JG (2005) Environmental Applications of Metabonomic Profiling. In: Robertson DG, Lindon J (eds) Metabonomics in toxicity assessment. Taylor & Francis, Boca Raton, pp 453–498
Bundy JG, Spurgeon DJ, Svendsen C, Hankard PK, Weeks JM, Osborn D, Lindon JC, Nicholson JK (2004) Environmental metabonomics: applying combination biomarker analysis in earthworms at a metal contaminated site. Ecotoxicology 13:797–806
Bundy JG, Keun HC, Sidhu JK, Spurgeon DJ, Svendsen C, Kille P, Morgan AJ (2007) Metabolic profile biomarkers of metal contamination in a sentinel terrestrial species are applicable across multiple sites. Environ Sci Technol 41:4458–4464
Bundy JG, Sidhu JK, Rana F, Spurgeon DJ, Svendsen C, Wren JF, Stürzenbaum SR, Morgan AJ, Kille P (2008) ‘Systems toxicology’ approach identifies coordinated metabolic responses to copper in a terrestrial non-model invertebrate, the earthworm Lumbricus rubellus. BMC Biol 6:25
Camacho D, de la Fuente A, Mendes P (2005) The origin of correlations in metabolomics data. Metabolomics 1:53–63
Canadian Council of Ministers of the Environment (CCME) (1999) Canadian soil quality guidelines for the protection of environmental and human health. Canadian Council of Ministers of the Environment, Winnipeg
Cooper EL, Roch P (1992) The capacities of earthworms to heal wounds and to destroy allografts are modified by polychlorinated biphenyls (PCB). J Invertebr Pathol 60:59–63
Craig A, Cloarec O, Holmes E, Nicholson JK, Lindon JC (2006) Scaling and normalization effects in NMR spectroscopic metabonomic data sets. Anal Chem 78:2262–2267
Cramer RD (1993) Partial least squares (PLS): its strengths and limitations. Perspect Drug Discov Des 1:269–278
Cui Q, Lewis IA, Hegeman AD, Anderson ME, Li J, Schulte CF, Westler WM, Eghbalnia HR, Sussman MR, Markley JL (2008) Metabolite identification via the Madison Metabolomics Consortium Database. Nat Biotechnol 26:162–164
Davis PW, Friedhoff JM, Wedemeyer GA (1972) Organochlorine insecticide, herbicide and polychlorinated biphenyl (PCB) inhibition of NaK-ATPase in rainbow trout. Bull Environ Contam Toxicol 8:69–72
Dhalla NS, Zhao D (1988) Cell membrane Ca2+/Mg2+ ATPase. Prog Biophys Mol Biol 52:1–37
Dieterle F, Ross A, Schlotterbeck G, Senn H (2006) Probabilistic quotient normalization as robust method to account for dilution of complex biological mixtures. Application in 1H NMR metabonomics. Anal Chem 78:4281–4290
Eijsackers H, Van Gestel CAM, De Jonge S, Muijs B, Slijkerman D (2001) Polycyclic aromatic hydrocarbon-polluted dredged peat sediments and earthworms: a mutual interference. Ecotoxicology 10:35–50
Environment Canada (2004) Biological test method. Tests for toxicity of contaminated soil to earthworms (Eisenia andrei, Eisenia fetida, or Lumbricus terrestris). Environment Canada, Ottawa, Canada
Eriksson L, Johansson E, Kettaneh-Wold N, Trygg J, Wikström C, Wold S (2006) Multi- and megavariate data analysis part I basic principles and applications. Umetrics, Umeå
Filzmoser P, Varmuza K (2010) Chemometrics: Multivariate Statistical Analysis in Chemometrics. R package version 0:8
Gastaldi L, Ranzato E, Caprì F, Hankard P, Pérès G, Canesi L, Viarengo A, Pons G (2007) Application of a biomarker battery for the evaluation of the sublethal effects of pollutants in the earthworm Eisenia andrei. Comp Biochem Physiol C 146:398–405
Gibb JOT, Svendsen C, Weeks JM, Nicholson JK (1997) 1H NMR spectroscopic investigations of tissue metabolite biomarker response to Cu(II) exposure in terrestrial invertebrates: Identification of free histidine as a novel biomarker of exposure to copper in earthworms. Biomarkers 2:295–302
Guo Q, Sidhu JK, Ebbels TMD, Rana F, Spurgeon DJ, Svendsen C, Stürzenbaum SR, Kille P, Morgan AJ, Bundy JG (2009) Validation of metabolomics for toxic mechanism of action screening with the earthworm Lumbricus rubellus. Metabolomics 5:72–83
Hawkins DM, Basak SC, Mills D (2003) Assessing model fit by cross-validation. J Chem Inf Comput Sci 43:579–586
Hines A, Staff FJ, Widdows J, Compton RM, Falciani F, Viant MR (2010) Discovery of metabolic signatures for predicting whole organism toxicology. Toxicol Sci 115:369–378
Horton HR, Moran LA, Scrimgeour KG, Perry MD, Rawn JD (2006) Principles of biochemistry. Pearson Prentice Hall, Upper Saddle River
Hughes SL, Bundy JG, Want EJ, Kille P, Stürzenbaum SR (2009) The metabolomic responses of Caenorhabditis elegans to cadmium are largely independent of metallothionein status, but dominated by changes in cystathionine and phytochelatins. J Proteome Res 8:3512–3519
Jackson JE (1991) A user’s guide to principal components. John Wiley and Sons, New York
Johnson HE, Lloyd AJ, Mur LAJ, Smith AR, Causton DR (2007) The application of MANOVA to analyse Arabidopsis thaliana metabolomic data from factorially designed experiments. Metabolomics 3:517–530
Johnson KE, Knopper LD, Schneider DC, Ollson CA, Reimer KJ (2009) Effects of local point source polychlorinated biphenyl (PCB) contamination on bone mineral density in deer mice (Peromyscus maniculatus). Sci Total Environ 407:5050–5055
Jones OAH, Spurgeon DJ, Svendsen C, Griffin JL (2008) A metabolomics based approach to assessing the toxicity of the polyaromatic hydrocarbon pyrene to the earthworm Lumbricus rubellus. Chemosphere 71:601–609
La Rocca PT, Carlson GP (1979) The effect of polychlorinated biphenyls on adenosine triphosphatase activity. Toxicol Appl Pharmacol 48:185–192
Landrum PF, Leppänen M, Robinson SD, Gossiaux DC, Burton GA, Greenberg M, Kukkonen JVK, Eadie BJ, Lansing MB (2004) Effect of 3, 4, 3′, 4′-tetrachlorobiphenyl on the reworking behavior of Lumbriculus variegatus exposed to contaminated sediment. Environ Toxicol Chem 23:178–186
Lenz EM, Weeks JM, Lindon JC, Osborn D, Nicholson JK (2005) Qualitative high field 1H-NMR spectroscopy for the characterization of endogenous metabolites in earthworms with biochemical biomarker potential. Metabolomics 1:123–136
Lerner DT, Björnsson BT, McCormick SD (2007) Effects of aqueous exposure to polychlorinated biphenyls (Aroclor 1254) on physiology and behavior of smolt development of Atlantic salmon. Aquat Toxicol 81:329–336
Leroy D, Haubruge E, De Pauw E, Thomé JP, Francis F (2010) Development of ecotoxicoproteomics on the freshwater amphipod Gammarus pulex: identification of PCB biomarkers in glycolysis and glutamate pathways. Ecotoxicol Environ Saf 73:343–352
Lingrel JB, Kuntzweiler T (1994) Na + , K + -ATPase. J Biol Chem 269:19659–19662
McKelvie JR, Yuk J, Xu Y, Simpson AJ, Simpson MJ (2009) 1H NMR and GC/MS metabolomics of earthworm responses to sub-lethal DDT and endosulfan exposure. Metabolomics 5:84–94
McKelvie JR, Wolfe DM, Celejewski M, Simpson AJ, Simpson MJ (2010) Correlations of Eisenia fetida metabolic responses to extractable phenanthrene concentrations through time. Environ Pollut 158:2150–2157
Meier JR, Chang LW, Jacobs S, Torsella J, Meckes MC, Smith MK (1997) Use of plant and earthworm bioassays to evaluate remediation of soil from a site contaminated with polychlorinated biphenyls. Environ Toxicol Chem 16:928–938
Morgenthal K, Weckwerth W, Steuer R (2006) Metabolomic networks in plants: transitions from pattern recognition to biological interpretation. BioSystems 83:108–117
O’Halloran K (2006) Toxicological considerations of contaminants in the terrestrial environment for ecological risk assessment. Hum Ecol Risk Assess 12:74–83
OECD (Organisation for Economic Co-operation, Development) (1984) Guideline for testing of chemicals No. 207. Earthworm acute toxicity test. OECD, Paris
OECD (Organisation for Economic Co-operation, Development) (2004) Guideline for testing of chemicals No. 222. Earthworm reproduction test. OECD, Paris, France
Parsons HM, Ekman DR, Collette TW, Viant MR (2009) Spectral relative standard deviation: a practical benchmark in metabolomics. Analyst 134:478–485
R Development Core Team (2009) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Robertson DG (2005) Metabonomics in toxicology: a review. Toxicol Sci 85:809–822
Rochfort SJ, Ezernieks V, Yen AL (2009) NMR-based metabolomics using earthworms as potential indicators for soil health. Metabolomics 5:95–107
Rousseau R, Govaerts B, Verleysen M, Boulanger B (2008) Comparison of some chemometric tools for metabonomics biomarker identification. Chemometr Intell Lab Syst 91:54–66
Schock TB, Stancyk DA, Thibodeaux L, Burnett KG, Burnett LE, Boroujerdi AFB, Bearden DW (2010) Metabolomic analysis of Atlantic blue crab, Callinectes sapidus, hemolymph following oxidative stress. Metabolomics 6:250–262
Scholz M, Selbig J (2007) Visualization and analysis of molecular data. Methods Mol Biol 358:87–104
Semple KT, Morriss AWJ, Paton GI (2003) Bioavailability of hydrophobic organic contaminants in soils: fundamental concepts and techniques for analysis. Eur J Soil Sci 54:809–818
Simpson AJ, Brown SA (2005) Purge NMR: effective and easy solvent suppression. J Magn Reson 175:340–346
Simpson MJ, McKelvie JR (2009) Environmental metabolomics: new insights into earthworm ecotoxicity and contaminant bioavailability in soil. Anal Bioanal Chem 394:137–149
Sousa A, Pereira R, Antunes SC, Cachada A, Pereira E, Duarte AC, Gonçalves F (2008) Validation of avoidance assays for the screening assessment of soils under different anthropogenic disturbances. Ecotoxicol Environ Saf 71:661–670
Spiegel MR (1991) Schaum’s outline of theory and problems of statistics. McGraw-Hill, New York
Steenland K, Deddens JA (2004) A practical guide to dose-response analyses and risk assessment in occupational epidemiology. Epidemiology 15:63–70
Steuer R (2006) Review: on the analysis and interpretation of correlations in metabolomic data. Brief Bioinform 7:151–158
Suzuki MM, Cooper EL, Eyambe GS, Goven AJ, Fitzpatrick LC, Venables BJ (1995) Polychlorinated biphenyls (PCBs) depress allogeneic natural cytotoxicity by earthworm coelomocytes. Environ Toxicol Chem 14:1697–1700
Taylor NS, Weber RJM, Southam AD, Payne TG, Hrydziuszko O, Arvanitis TN, Viant MR (2009) A new approach to toxicity testing in Daphnia magna: Application of high throughput FT-ICR mass spectrometry metabolomics. Metabolomics 5:44–58
United States Environmental Protection Agency (EPA) (1990) Guidance on remedial actions for superfund sites with PCB contamination. United States Environmental Protection Agency, Washington DC, USA
Van Scoy AR, Yu Lin C, Anderson BS, Philips BM, Martin MJ, McCall J, Todd CR, Crane D, Sowby ML, Viant MR, Tjeerdema RS (2010) Metabolic responses produced by crude versus dispersed oil in Chinook salmon pre-smolts via NMR-based metabolomics. Ecotoxicol Environ Saf 73:710–717
Varmuza K, Filzmoser P (2009) Introduction to multivariate statistical analysis in chemometrics. CRC Press, Boca Raton
Vendeginste BGM, Massart DL, Buydens LMC, de Jong S, Lewi PJ, Smeyers-Verbeke J (1998) Handbook of chemometrics and qualimetrics: Part B. Elsevier, Amsterdam
Verouden MPH, Westerhuis JA, van der Werf MJ, Smilde AK (2009) Exploring the analysis of structured metabolomics data. Chemometr Intell Lab Syst 98:88–96
Viant MR (2008) Recent developments in environmental metabolomics. Mol Biosyst 4:980–986
Ville P, Roch P, Cooper EL, Masson P, J- Narbonne (1995) PCBs increase molecular-related activities (lysozyme, antibacterial, hemolysis, proteases) but inhibit macrophage-related functions (phagocytosis, wound healing) in earthworms. J Invertebr Pathol 65:217–224
Vulimiri SV, Berger A, Sonawane B (2010) The potential of metabolomic approaches for investigating mode(s) of action of xenobiotics: Case study with carbon tetrachloride. Mutat Res. doi:10.1016/j.mrgentox.2010.02.013
Warne MA, Lenz EM, Osborn D, Weeks JM, Nicholson JK (2000) An NMR-based metabonomic investigation of the toxic effects of 3- trifluoromethyl-aniline on the earthworm Eisenia veneta. Biomarkers 5:56–72
Weber R, Gaus C, Tysklind M, Johnston P, Forter M, Hollert H, Heinisch E, Holoubek I, Lloyd-Smith M, Masunaga S, Moccarelli P, Santillo D, Seike N, Symons R, Torres JPM, Verta M, Varbelow G, Vijgen J, Watson A, Costner P, Woelz J, Wycisk P, Zennegg M (2008) Dioxin- and POP-contaminated sites—Contemporary and future relevance and challenges: overview on background, aims and scope of the series. Environ Sci Pollut Res 15:363–393
Westerhuis JA, Hoefsloot HCJ, Smit S, Vis DJ, Smilde AK, Velzen EJJ, Duijnhoven JPM, Dorsten FA (2008) Assessment of PLSDA cross validation. Metabolomics 4:81–89
Whitfield Åslund ML, Celejewski M, Lankadurai BP, Simpson AJ, Simpson MJ Natural variability and correlations in the metabolic profile of healthy Eisenia fetida earthworms observed using 1H NMR metabolomics. Chemosphere. doi:10.1016/j.chemosphere.2011.01.036
Acknowledgments
We gratefully acknowledge funding for this research from the Natural Science and Engineering Research Council (NSERC) of Canada in the form of a Strategic Grant and Discovery Grant to MJS. AJS would like to thank the Ontario Government for an Early Researcher Award. We also thank Dr. Jennifer McKelvie, Brian Lankadurai, Jimmy Yuk, and Magda Celejewski for valuable discussions.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Whitfield Åslund, M.L., Simpson, A.J. & Simpson, M.J. 1H NMR metabolomics of earthworm responses to polychlorinated biphenyl (PCB) exposure in soil. Ecotoxicology 20, 836–846 (2011). https://doi.org/10.1007/s10646-011-0638-9
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10646-011-0638-9