
Abstract
Nelusetta ayraudi (the ocean leatherjacket) is an endemic Australian monacanthid species distributed from North West Cape (Western Australia) south to southern Queensland. The commercial and recreational fisheries targeting Nelusetta ayraudi have expanded substantially along the coast of New South Wales (NSW) in recent years but there exists little biological information on which to base effective management of this growing fishery. World-wide, only a few studies have aged monacanthids. Of these, researchers have interpreted periodic increments in bony structures such as vertebrae and anterior dorsal spines in preference to those found in otoliths. In this study we estimated age of N. ayraudi by counting growth increments in sectioned otoliths. The periodicity of increment formation was validated using a vital stain, (oxy-tetracycline), injected into young-of-the-year fish. Growth was rapid especially as juveniles with N. ayraudi attaining approximately 220 mm after 1 year and 340 mm after 2 years. No differences in growth rates were detected between sexes or between fish captured at different latitudes (zones). The largest male (605 mm, Total Length—TL) and female (656 mm, TL) were both recorded from northern NSW, with both sexes attaining the maximum age of 6+ years from northern and southern NSW. The von Bertalanffy parameters describing growth for N. ayraudi were \( {L_\infty } \) = 591 mm (TL), k = 0.377 year−1 and to = −0.247 years.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Leatherjackets or filefish of the family Monacanthidae belong to the highly modified and advanced group of fish which form the order Tetraodontiformes. Monacanthids are well represented throughout the world’s oceans with 102 species being recorded (Nelson 2006). The highest monacanthid diversity is found in Australian waters with a total of 60 species being recorded, 22 of which are only found in the southern half of the continent (<30°S), in cooler temperate waters (Hutchins 2000; Allen et al. 2006). Of the many monacanthids inhabiting these waters, the ocean leatherjacket (Nelusetta ayraudi), also known as the chinaman or sand leatherjacket, is one of largest monacanthids in the world and the most commonly captured species from Australian waters (Miller 2007). Thought to be endemic to Australian waters, N. ayraudi are distributed from North West Cape in Western Australia around the south of the continent to Cape Morton in Queensland (Kailola et al. 1993). In New South Wales (NSW) waters, annual commercial landings of N. ayraudi have increased rapidly from 134 to 430 tonnes between 2000–2001 and 2006–2007 (Miller and Stewart 2009). Despite the rapidly expanding commercial fishery, there exists almost no information on the biology of N. ayraudi on which to base the management of this species.
Ageing monacanthids has proven to be difficult and the available literature indicates a lack of studies using otoliths to estimate age. The few studies that have used otoliths to estimate age have examined daily incremental development and growth of embryonic and pre-larval settling juveniles using whole sagittal otoliths (Kingsford and Milicich 1987; Kawase and Nakazono 1994; Rogers et al. 2001; Ben-David and Kritizer 2005). The lack of studies using otoliths to estimate age in post-larval monacanthids is curious given that otoliths are recognized as being the most widely used and reliable structure for estimating age in bony fish (Campana 2001). One reason may be that monacanthids have very small and fragile otoliths which consequently can be difficult to process and interpret successfully (Grove-Jones and Burnell 1991; Mancera-Rodriguez and Castro-Hernandez 2004). This difficulty, coupled with the fact that few monacanthid fisheries are of sufficient economic value to warrant substantial research, appears to have hampered age estimation in this group of fish. Studies that have reported estimates of age and growth in monacanthids have used vertebrae (Shiqin and Yachu 1980; Park 1985; Grove-Jones and Burnell 1991), anterior dorsal spines (Johnson and Saloman 1984; Manooch and Drennon 1987; Mancera-Rodriguez and Castro-Hernandez 2004) and length frequency analyses (Peristiwady and Geistdoerfer 1991). Validation of the accuracy and precision of these alternative methods of estimating age and growth is limited.
In this paper we describe the use of otoliths to estimate the age of N. ayraudi. Age was estimated by counting opaque zones in sectioned sagittal otoliths from wild caught fish and the method was validated by using a vital stain (oxy-tetracycline—OTC) injected into young-of-the-year-fish. Estimates of size-at-age were used to model rates of growth, which were compared between sexes and between latitudes (zones) in eastern Australia.
Materials and methods
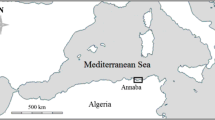
N. ayraudi were collected from commercial catches from two zones along the coast of NSW. The northern zone was described as the area of coastline stretching from Forster/Tuncurry (32°11′.00 S, 152°31′.00 E) to the border of NSW and Queensland (28°10′.02 S, 153°33′.03 E). The southern zone was described as the region stretching from Kiama (34°40′.20 S, 150 51′.54 E) to the Victorian border (37°31′.50 S, 149°58′.70 E) (Fig. 1). Twenty fish per month were collected from each zone between March 2003 and March 2004. Due to a limited number of large (older) fish in these monthly samples, a further 19 large fish were collected from commercial catches at Iluka and Ballina (northern zone) during September 2005. Sagittal otoliths were removed from these fish for age estimation. Juvenile N. ayraudi were also sampled each month from the Port Hacking River (34°47′.50 S, 151°08′.00 E) (Fig. 1), between December 2002 and April 2003 and their total lengths (TL, mm) recorded to estimate monthly growth of the cohort.

Study locations along the coast of New South Wales, Australia, showing sampling zones
Oxy-tetracycline was used to stain the otoliths of wild-caught young-of-the-year (0+) captive reared fish. During December 2002, juvenile N. ayraudi were collected from the Port Hacking River using small opera house traps covered with 12 mm mesh set on a seagrass (Zostera capricorni and Posidonia australis)/sand bottom. Fish were placed into aerated 5,000 L tanks with flow through ambient seawater and maintained prior to injection with OTC in March of 2003. Prior to injection the fish were sedated with 50 mL of benzocaine solution (ethyl-p-amino benzoate dissolved in 100% ethanol at 100 g·1,000 mL−1), weighed (g), measured (TL, mm) and given an intraperitoneal injection of tetracycline (Engemycin 100—oxytetracycline hydrochloride at 5 mg·mL−1) at a dose of 50 mg·kg−1 body weight (McFarlane and Beamish 1987). Any leatherjacket louse (Ourozeuktes owenii) found attached to the fish was removed at this time. Between 2002 and 2004, fish were sampled intermittently and their otoliths removed for examination. Sagittal otoliths were removed by cutting horizontally above the eyes of the fish, exposing the brain cavity. The otolith sacculus, extending downwards on either side of the forefront of the brain cavity, were extracted, otoliths removed and allowed to dry for a short period of time before being set in resin blocks. A single bladed low speed diamond saw was used to cut a thin (∼0.5 mm) cross-section from the dorsal to ventral margins through each otolith core (primordium). These otolith sections were mounted on a glass slide and viewed under reflected light against a black background with 4× magnification. Counts of opaque zones were made along a radius from the core to the outer edge of the ventral lobe of the otolith using a microscope mounted video camera interfaced with a computer, running ‘Image Pro Plus’ image analysis software (Version 4.5.1, media Cybernetics Inc, Bethesda, MD). The otoliths from fish that were injected with OTC were observed under reflected light and opaque zones counted and measured. Observations were then made using ultraviolet (UV) light to identify the marked area of OTC on the otolith. Each otolith was measured from the core to the OTC mark and to the outside edge of the otolith. All otoliths were re-read to examine the precision of estimates of counts of opaque zones. The coefficient of variation (CV) for the two readings for each otolith was calculated and averaged across all otoliths (Campana 2001).
A birth date of 1st August was assigned to all fish based on the peak in the spawning season (Miller 2007). Age was calculated as the number of opaque zones plus the proportion of the year following this date that the fish was sampled. N. ayraudi have a short winter spawning period (Miller 2007) and therefore errors in under-estimating or over-estimating the absolute age of fish born slightly earlier or later than 1st August will be minor. The von Bertalanffy growth function (VBGF) was fitted to the size-at-age data for wild caught ocean leatherjackets, using solver in Microsoft Excel 2003 to minimise the sum of squares. An analysis of residual sums of squares (ARSS) (Chen et al. 1992) was used to compare the VBGFs between: (1) sexes from each latitudinal zone, and; (2) latitudinal zones.
Results
Otoliths from N. ayraudi are typical of those described for other monacanthids (Furlani et al. 2007) in being very small, hour-glass shaped, the dorsal margin flat and slightly irregular and the ventral margin rounded and irregular (Fig. 2a). They had a diameter from the posterior to anterior margins of between 0.5 mm in younger fish to around 3 mm in older fish and were very fragile. When sectioned, the internal structure of all otoliths appeared complex. The core was densely opaque with a larger translucent zone surrounding it with scattered finer streaks of opaque markings throughout. This zone represented the rapid growth fish experience while juvenile. This structure was followed by a more definite opaque zone that was scored as the first annulus. The mean (± SE) distance (mm) from the core to the outside edge of the first annulus in the 585 wild fish examined was 0.345 mm ± 0.002 mm. Annuli beyond the first opaque zone were characterized by reasonably well defined alternating opaque and translucent zones (Fig. 2b).

a Whole otolith, scale: 0.5 mm; b thin-sectioned otolith viewed under reflected light against a black background. The estimated age and size of this fish was 4+ years and 551 mm, Scale: 0.5 mm; c a captive reared 2+ year old sectioned otolith under reflected light, Scale: 0.25 mm; and, d the same captive reared 2+ year old sectioned otolith under ultra violet light displaying the oxy-tetracycline (OTC) mark, Scale: 0.25 mm
The young-of-the-year N. ayraudi that were injected with OTC and maintained in the aquaria ranged between 97 and 193 mm TL. The otoliths of sampled fish all had clearly visible OTC marks when viewed under UV light (Fig. 2c, d). Thirty were sampled before they formed their first annulus, 21 were sampled when 1+ years and 7 were sampled when 2+ years (Fig. 3). Otolith growth from the OTC mark to the edge of the otolith indicated that the otoliths increased in diameter approximately linearly during the 26 months of the validation experiment (Fig. 3). The mean (± SE) distance from the core to the outside edge of the first opaque zone in these otoliths was 0.325 mm ± 0.006 mm which was similar to fish from the wild (see above). The OTC mark, administered during March, was always present within the translucent zone prior to the first opaque zone, indicating that the opaque zone was probably formed sometime during the winter period. The first opaque zone was scored as completed in 1 of 4 fish during July and in all fish by August.

Sagittal otolith growth after the oxy-tetracycline (OTC) mark in February 2003 (n = 58)
Estimates of age were assigned to the 585 otoliths examined from both the northern and southern zones. Of the otoliths re-examined, 80.5% were not different from the original readings, 18.8% differed by ± 1 year and 0.7% differed by ± 2 years. The coefficient of variation (CV), averaged across all ages, was 0.066. Six age classes (1 to 6 years) were recorded from the 585 wild caught fish sampled from commercial landings. Only two fish, one male and one female were estimated as being 6+ years old. The largest recorded fish was a female from the northern zone which measured 656 mm (TL) and was estimated at 5.8 years. The largest male (605 mm TL) was also from the northern zone and was estimated to be 5.2 years old. The largest fish from the southern zone was a female (596 mm TL) estimated at 3.8 years old and the largest male from the southern zone (583 mm TL) was estimated to be 4.2 years old.
Comparisons of rates of growth between sexes from each zone revealed no significant differences (ARSS northern zone, f (3,312) = 0.55, P = 0.64 and southern zone, f (3, 261) = 1.38, P = 0.25). Similarly, there were no significant differences observed in rates of growth when the two sexes were combined between each zone (ARSS f (3, 579) = 1.34, P = 0.26). As there were no significance differences in the rates of growth between sexes and zones, growth data for all fish were pooled to provide a general VBGF for N. ayraudi. The VBGF parameters (±SE) estimated were k = 0.377 year−1 (±0.0224) t o = −0.247 year (±0.032), and \( {{\hbox{L}}_\infty } \) = 591 mm (±15.90) (Fig. 4). The relationship between TL and body weight for all fish sampled was described by a power relationship: Body weight (g) = 0.00003 × TL2.808.

Estimated length at age plot for N. ayraudi. The von Bertalanffy growth function (VBGF) parameters for all fish (including juveniles) combined from each of the two zones are shown. Note: open squares = fish aged from otoliths; closed diamonds = juvenile fish aged from the nominated birth date
Sampling of young-of-the-year N. ayraudi between December and April indicated rapid growth during this period (Table 1). The mean length of juvenile N. ayraudi sampled during April was 176 mm TL and, based on a birthday of 1st August, these fish were approximately 9 months old.
Discussion
This study has demonstrated that the technique of using sectioned otoliths to estimate the age of fish (Campana 2001) can be successfully applied to a member of the family Monacanthidae. While the otoliths of this diverse and abundant family of fish are very small, fragile and irregular in shape (Furlani et al. 2007), they display alternating opaque and translucent zones that can be counted and used to estimate age. A review of the literature on ageing monacanthids, and the closely related family of Balistidae, shows that our results in demonstrating the utility of using sectioned otoliths to estimate age could lead to improvements in future studies of age and growth in these families of fish. In Korea (Park 1985), South Australia (Grove-Jones and Burnell 1991), and China (Shiqin and Yachu 1980), studies have estimated the age of black filefish (Navodon modestus), ocean leatherjackets (Nelusetta ayraudi) and green filefish (Navodon septentrionalis) using sectioned vertebrae. Anterior dorsal spines have been used in other ageing studies for the plane-head filefish (Stephanolepis hispidus) from the Canary Islands (Mancera-Rodriguez and Castro-Hernandez 2004), the closely related balistids, the gray triggerfish (Balistes capriscus) in the north eastern gulf of Mexico (Johnson and Saloman 1984), and the queen triggerfish (Balistes vetula) from the U.S Virgin Islands and Puerto Rico (Manooch and Drennon 1987). None of these studies made use of sectioned otoliths, with an accompanying validation, which is generally accepted as the best ageing technique for the majority of fish species (Campana 2001). Rather, researchers have interpreted the concentric bands in vertebrae and dorsal spines when estimating annual ages. These bony structures are not ideal for estimating age because skeletal growth can be variable and is influenced by many internal and external factors. Vertebrae continually resorb tissue which can destroy periodic growth increments and the spines of older fish can become hollow and lose record of younger growth patterns (Panfili et al. 2002). In addition, the precision of readings from both vertebrae and spines tend to be poor compared to those from otoliths and validation of the ageing interpretations difficult (Campana 2001). The negative ramifications of using incorrect age estimates in fishery management are well documented (Campana 2001) and our findings that sectioned otoliths can be used to age monacanthids and balistids will be of use to future studies of these families of fish. Nelusetta ayraudi sagittal otoliths were well stained by OTC at a dosage of 50 mg·kg−1 body weight. The 58 otoliths examined as part of the validation experiment had clearly visible OTC marks when viewed under ultra-violet light. Marking these fish with OTC showed that opaque zones were formed once per year during the winter. Many other temperate species in Australian waters have also been shown to form annual opaque zones during the winter months (Stewart and Hughes 2007; Morton et al. 2008). The finding that otolith growth from the core to the first opaque zone of fish in aquaria was similar to that of wild fish provided some confidence that the captivity did not affect the validation experiment.
The internal structure of otoliths from N. ayraudi was complex and alternating opaque and translucent zones were often diffuse. This pattern was interpreted as being the result of a fast growth rate, particularly during the early years. Variations in somatic growth have been shown to effect the internal structural development of otoliths (Fowler 1995), with fish having faster growth rates being associated with less distinct annuli (Esteves and Burnett 1993; Stewart and Hughes 2007). Nevertheless, the overall precision from ageing N. ayraudi (mean CV of 6.6%), was comparable to those reported in other ageing studies (Campana 2001).
Growth of juvenile monacanthids has been poorly documented for other species that are captured in large commercial fisheries around the world. Data exist for the growth of the early stages of development of several juvenile species such as Parika scaber in New Zealand (Kingsford and Milicich 1987), Stephanolepis hispidus in the USA (Rogers et al. 2001) Navodon modestus (Kakuda 1979) and Rudarius ercodes and Paramonacanthus japonicus in Japanese waters (Kawase and Nakazono 1994). Ishida and Tanaka (1983) showed recruitment and growth of juvenile small filefish (Rudarius ercodes) in Odawa Bay Japan, while Peristiwady and Geistdoerfer (1991) reported recruitment of juvenile Monacanthus tomentosus in sheltered bays and estuaries of West Seram, Moluccas, Indonesia. In the present study, juvenile N. ayraudi exhibited rapid growth during the summer months between December and April. A mean maximum monthly growth of 31 mm occurred during February. Pollard (1994) noted that seagrass beds (Zostera capricorni and Posidonia australis) played an important role in the early stages of growth for juvenile N. ayraudi and suggested that the species migrated to deeper waters as it grew larger. Grove-Jones and Burnell (1991) also had the highest capture rates of juvenile N. ayraudi from sheltered bays and estuaries, in the months of February, with a decline in numbers in the months that followed. Again, they assumed that these fish were recruiting to the fishery offshore. They also found that the mean growth rates were highest in February and April (27 mm and 28 mm respectively). N. ayraudi is amongst the largest monacanthids in the world, with a maximum recorded length of 700 mm (Hutchins and Swainston 1986). The largest fish sampled during the present study were substantially smaller than this, suggesting that ocean leatherjackets may attain ages considerably greater than 6+ years. A summary of ageing studies done world-wide indicates that other species of monacanthids are relatively short lived (<9 years) (Table 2). Nevertheless, our results indicate very fast growth rates. No differences in growth rates were found between males and females. The lack of differences in growth rates with latitude may be a result of migration between our sampling zones.
Commercial fishers in NSW have provided anecdotal evidence that commercially-sized fish undergo large latitudinal movements seasonally, especially during winter when fish are spawning. Although these spawning migrations have not been confirmed in NSW, Lindholm (1984) reported that N. ayraudi in South Australian waters were highly mobile and that larger fish were captured from deeper waters. This study has demonstrated that monacanthids can be successfully aged using sectioned otoliths and recommends that future studies on this and other closely related families adopt our methods. We consider that the improvements in accuracy and precision by using sectioned otoliths in preference to the commonly used vertebrae and dorsal spines outweigh the difficulties in handling small, fragile otoliths. It would be useful to use sectioned otoliths to age species that have previously been studied using vertebrae and dorsal spines. This new information on age and growth of N. ayraudi in eastern Australia has provided the first insight into how the stock may respond to the rapidly increasing fishery. The relatively few age classes and rapid growth rates suggest that N. ayraudi maybe resilient to fishing; however further information on the reproductive biology, life-history, movements, stock structure and fishery landings are still required if informed management is to be implemented.
References
Allen GR, Allen JC, Cross NJ (2006) Zoological Catalogue of Australia. Volume 35. Fishes. Part 3 (of 3). Actinopterygii: Acanthopterygii (in part): Perciformes. (Notothenioidei to Stromateoidei) to Tetraodontiformes. Australian Government. Department of the Environment and Heritage. CSIRO Publishing
Ben-David J, Kritizer JP (2005) Early life history and settlement of the slender filefish, Monacanthus tuckeri (Monacanthidae), at Calabash Caye, Turneffe Atoll, Belize. Environ Biol Fish 73:275–282
Campana SE (2001) Accuracy, precision and quality control in age determination, including a review of the use and abuse of age validation methods. J Fish Biol 59:197–242
Chen Y, Jackson DA, Harvey HH (1992) A comparison of von Bertalanffy and polynomial growth functions in modelling fish growth data. Can J Fish Aquat Sci 49(6):1228–1235
Chen WZ, Zheng YJ, Li CS (1998) Stock assessment and catch predictions of the filefish, Thamnaconus septentrionalis, with the help of microcomputer. Journal of Shanghai Fisheries University 7(1):13–18
Esteves C, Burnett JM (1993) A comparison of growth rates for American plaice, Hippoglossoides platessoides, in the Gulf of Maine-Georges Bank region derived from two different data sources. Ref. Doc. 93-09, Woods Hole Lab. Natl. Mar. Fish. Serv. NOAA, Woods Hole MA 02543, p 8
Fowler AJ (1995) Annulus formation in otoliths of coral reef fish—a review. In: Secor DH, Dean JM, Campana SE (eds) Recent developments in fish otolith research. University of South Carolina Press, Columbia, pp 319–329
Furlani D, Gales R, Pemberton D (2007) Otoliths of common temperate Australian fish: a photographic guide. CSIRO, Melbourne, 208 pp
Grove-Jones RP, Burnell AF (1991) Fisheries biology of the ocean leatherjacket (Monacanthidae: Nelusetta ayraudi) in the eastern waters of the Great Australian Bight, South Australia. Fishing Industry Research and Development Council, Grant No: DFS01Z. pp 1–107
Hutchins B (2000) Leatherjackets. In: Andrew N (ed) Under Southern Seas: the ecology of Australia’s rocky reefs. Krieger, Malabar, pp 194–201
Hutchins B, Swainston R (1986) Sea fishes of southern Australia. Complete field guide for anglers and divers. Swainston, Perth
Ishida Y, Tanaka S (1983) Growth and maturation of the Small Filefish Rudarius ercodes in Odawa Bay. Bull Jpn Soc Sci Fish 49:547–553
Johnson AG, Saloman CH (1984) Age, growth, and mortality of gray triggerfish, Balistes capriscus, from the north-eastern gulf of Mexico. Fish Bull 82(3):485–492
Kailola PJ, Williams MJ, Stewart PC, Russell RE, Mcnee A, Grieve C (1993) Australia Fisheries Resource. Bureau of Resource Sciences, Department of Primary Industries and Energy, and the Fisheries Research and Development Corporation. Commonwealth of Australia
Kakuda S (1978) On the spawning clusters of the filefish Navodon modestus in the Seto Inland Sea. J Fac Fish Anim Husb Hiroshima Univ 17:165–173
Kakuda S (1979) On the growth of the File-fish, Navodon modestus, in the Seto Inland Sea. J Fac Appl Biol Sci 18:197–205
Kawase H, Nakazono A (1994) Embryonic and pre-larval development and otolith increments in two filefishes, Rudarius ercodes and Paramonacanthus japonicus (Monacanthidae). Jpn J Ichthyol 41(1):57–63
Kingsford M, Milicich MJ (1987) Presettlement phase of Parika scaber (Pisces: Monacanthidae): a temperate reef fish. Mar Ecol Prog Ser 36:65–79
Lindholm R (1984) Observations on the chinaman leatherjacket Nelusetta ayraudi (Quoy and Gaimard) in the Great Australian Bight. Aust J Mar Freshwat Res 35:597–599
Mancera-Rodriguez NJ, Castro-Hernandez JJ (2004) Age and growth of Stephanolepis hispidus (Linnaeus, 1766) (Pisces: Monacanthidae), in the Canary Islands area. Fish Res 66:381–386
Manooch CS III, Drennon CL (1987) Age and growth of yellowtail snapper and queen triggerfish collected from the U.S. Virgin Islands and Puerto Rico. Fish Res 6:53–68
McFarlane GA, Beamish RJ (1987) Selection of dosages of oxytetracycline for age validation studies. Can J Fish Aquat Sci 44:905–909
Miller ME (2007) Key biological parameters and commercial fishery for ocean leatherjackets, Nelusetta ayraudi, (Monacanthidae) off the coast of New South Wales, Australia. Master of Environmental Science, School of Biological Sciences, University of Wollongong, NSW, Australia. http://ro.uow.edu.au/theses/26
Miller ME, Stewart J (2009) The commercial fishery for ocean leatherjackets (Nelusetta ayraudi, Monacanthidae) in New South Wales, Australia. Asian Fish Sci 22:257–264
Morton JK, Gladstone W, Hughes JM, Stewart J (2008) A comparison of the life histories of three co-occurring wrasses (Teleostei: Labridae) in coastal waters of south-eastern Australia. Mar Freshw Res 59:560–574
Nelson JS (2006) Fishes of the world, 4th edn. Wiley, Hoboken, 601 pp
Panfili J, Pontual H (de), Troadec H, Wright PJ (eds) (2002) Manual of fish sclerochronology. Brest, France: Ifremer-IRD coedition, 464 p
Park BH (1985) Studies on the fishery biology of the Filefish Navodon modestus (Gunther) in Korean Waters. Bull Fish Res Dev Agency (South Korea) 34:1–64
Peristiwady T, Geistdoerfer P (1991) Biological aspects of Monacanthus tomentosus (Monacanthidae) in the seagrass beds of Kotania Bay, West Seram, Moluccas, Indonesia. Mar Biol 109:135–139
Pollard DA (1994) A comparison of fish assemblages and fisheries in intermittently open and permanently open coastal lagoons on the south coast of NSW, South-Eastern Australia. Estuaries 17(3):631–646
Rogers SJ, Hare JA, Lindquist DG (2001) Otolith record of age, growth and ontogeny in larval and pelagic juvenile Stephanolepis hispidus (Pisces: Monacanthidae). Mar Biol 138:945–953
Shiqin C, Yachu H (1980) A preliminary study on the age and growth of filefish (Navodon septentrionalis). Journal of Fisheries of China 4(2):197–206
Stewart J, Hughes JM (2007) Age validation and growth of three commercially important Hemiramphids in south-eastern Australia. J Fish Biol 70:65–82
Acknowledgements
This study could not have been possible without the support and enthusiasm of the commercial fishermen from the NSW ocean trap (fin-fish) and deep water lobster fisheries. Thank you to Noel Gogerly, Phil Roach, Ray Blake, and Steve Drake for their expertise in catching ocean leatherjackets.
Author information
Authors and Affiliations
Corresponding author
Additional information
An erratum to this article can be found at http://dx.doi.org/10.1007/s10641-010-9675-0
Rights and permissions
About this article
Cite this article
Miller, M.E., Stewart, J. & West, R.J. Using otoliths to estimate age and growth of a large Australian endemic monocanthid, Nelusetta ayraudi (Quoy and Gaimard, 1824). Environ Biol Fish 88, 263–271 (2010). https://doi.org/10.1007/s10641-010-9639-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10641-010-9639-4